
Callulina kanga, Loader, Simon P., Gower, David J., Müller, Hendrik & Menegon, Michele, 2010
|
publication ID |
https://doi.org/ 10.5281/zenodo.199640 |
|
DOI |
https://doi.org/10.5281/zenodo.5616313 |
|
persistent identifier |
https://treatment.plazi.org/id/03C99D08-FFC9-877D-269B-224BFAC6FEA7 |
|
treatment provided by |
Plazi |
|
scientific name |
Callulina kanga |
| status |
sp. nov. |
Callulina kanga View in CoL sp. nov.
Figures 1 View FIGURE 1. a , 4 View FIGURE 4 , 5 View FIGURE 5 , 6 View FIGURE 6 , 7, 8; Tables 1 View TABLE 1 , 2 View TABLE 2 , 3 View TABLE 3 .
Callulina sp. 4 Menegon et al. (2008) p.114, appendix 1, and table 3, 4.
Holotype. MTSN 8205, an adult male, collected in Kanga Forest Reserve, Mwomero District, Morogoro Region, Tanzania, 5° 58' 37.67''S, 37°41'57.28''E, 760 m by Michele Menegon on November 4th 2004 (see Figure 1 View FIGURE 1. a b). This specimen has also been sequenced for partial fragments of 12S, 16S and cytb (see Genetic difference and Appendix 1).
Paratype. MTSN 5017, an adult male, collected at the Kanga Forest Reserve, Mwomero District, Morogoro Region, Tanzania, 05°55'00.4" S, 037°42'20.1" E, 1140 m by Frontier-Tanzania on 19/04/06. MTSN 5017 has been sequenced for partial fragments of 12S, 16S, and cytb (see Appendix 1).
Diagnosis. The new species of Callulina is assigned to the brevicipitid genus based on the following characteristics: Truncated or expanded terminal phalanges (simple in Spelaeophyrne, Probreviceps , Breviceps , and Balebreviceps ); single posterior denticulated row in the palate of Callulina (two denticulated rows in Probreviceps , glandular mass in Breviceps ).
A small sized Callulina , Snout-urostyle length 23.1–23.7 mm, TL: SUL ratio 0.36–0.38. Tympanum present but sometimes obscured by granular skin, TD: SUL ratio 0.04–0.05. TD: TED ratio 0.75–0.85. Fingertips expanded, with the ratio between the width at the first subarticular versus width at distal phalanx 0.70. The species lacks any glandular masses on arms or legs and also lacks interocular patterning.
Callulina kanga most clearly resembles C. kreffti . Generally, C. kreffti have a larger tympanum relative to the distance of tympanum to the eye, while in C. kanga the tympanum diameter is less than the distance between tympanum and eye. However, in some specimens of C. kreffti the distance of tympanum to the eye is greater than the size of the tympanum (eight of 58 specimens: BMNH 1974.24, 2000-205, ZMB 21777, 23341, MCZ A 13624-25, 25493, USNM 200072) (see Table 2 View TABLE 2 and Figure 4 View FIGURE 4 ). Tympanum size seems to be sexually dimorphic ( Loader et al. 2009a for C. dawida and C. hanseni ) such that differences might also reflect overlap between the different sexes, but not all specimens were sexed to evaluate this. Morphologically C. kanga and C. kreffti are very similar, but are clearly distinguished based only on call and molecular data and highly disjunct distribution.
With the exception of C. kreffti , C. kanga differs from all other species in expansion of fingertips (<0.70 distal width at third terminal phalanx - distal width at subarticular tubercle), presence of tympanum (absent in C. laphami and C. shengena ), absence of large continuous glandular mass on legs ( C. dawida , C. hanseni , and C. shengena ). The species can also be distinguished on the basis of DNA sequence, call, and distribution data.
Description of holotype. (See Figure 5 View FIGURE 5 ) Body robust. Tips of fingers expanded, distal width of terminal phalanx 0.70 of the width of distal subarticular tubercle. Tips of fingers expanded laterally, with circummarginal grooves posterior to distal phalanx. Distal phalanx expanded beyond the width of the base of the phalanx. First finger shortest, second and fourth equal, third longest. Inner metatarsal tubercle rounded and raised, separated marginally at the point of contact on the edge of the palm by a wedge-shaped medial palmar tubercle from a larger outer metatarsal tubercle. Slightly smaller palmar tubercles present on palm. Large subarticular tubercles at the base of each finger. Third and fourth finger with smaller paired tubercles between basal articular tubercle and subarticular tubercles. Dorsoventrally expanded toe tips with slightly folded but smooth ventral surface. Tips of toes slightly expanded laterally, with circummarginal grooves posterior to distal phalanx. Toe tips with generally smooth ventral surface with some folds. First toe same length as second, third and fifth equal and larger than first and second toe, fourth longest. Inner metatarsal tubercle large, rounded and raised, almost touching a smaller sized, rounded, raised, outer metatarsal tubercle. Smaller palmar tubercles present on base of foot. Subarticular tubercles at the base of each toe, large subarticular tubercles on third, fourth and fifth toe at the phalangeal joints. All tubercles on hands and feet grey/blue/pale colour against pale brown background. Nostrils rounded, nearer to tip of snout than eye. Interorbital space flat, larger than width of eyelid. Pupil horizontal. Tympanum present and ovoid, higher than wide. Canthus rostralis rounded, loreal region sloping slightly concavely. Snout just visible in ventral view. Vent is posteroventrally positioned. Tongue rounded. Single posterior denticulated row on the palate. Incision around tympanic region on left side, and large, square-shaped piece of body wall missing on left ventrolateral side at midbody.
Dorsum brown with scattered, darker brown/black regular patches. Two large middorsal pale irregular patches are evident, one level with the arm insertion and the other on the sacral region. Skin granular, with slightly larger glandular masses on lateral sides and posterior end around arm insertion, thighs, and urostyle region. Ventral surface paler brown, cream-tan. Symmetrical, irregular white blotches on either side of the chin, extending to the corner of the mouth, and part of the canthus. Tympanic region pale brown/cream. Dark edged tympanic ridge extends from posterior edge of eyelid to arm insertion. Loreal and canthal regions dark brown.
Measures of holotype. ED = 2.8; ETD = 1.3; HL = 7.0; IOD = 4.7; JW = 7.4; LF3 = 2.9; LT4 = 3.9; ND = 1.6; NED = 2.0; NLD = 1.0; SUL = 23.7; TD = 1.1; TL = 8.7; TSL = 5.9; WDF3 = 1.0; WDTF3 = 0.7.
Measures of paratype (MTSN 5017 – see Figure 6 View FIGURE 6 ).— ED = 2.9; ETD = 1.2; HL = 7.4; IOD = 4.7; JW = 7.6; LF3 = 3.0; LT4 = 3.7; ND = 1.4; NED = 2.1; NLD = 1.0; SUL = 23.1; TD = 0.9; TL = 8.8; TSL = 6.1; WDF3 = 1.0; WDTF3 = 0.7.
Morphological and colour variation. The paratype is very similar to the holotype in the overall body proportions and key morphometric measures. The paratype has a much smoother skin, and has only a few dark symmetrical patches on the dorsum. Incision on left hand side of the tympanic region.
Colour in life. An overall brown colouration with scattered whitish areas giving parts a marbled appearance. The whitish areas are more conspicuous around the eyes and eyelids, tympanum area and on the back. Small white-tipped warts are irregularly present, especially on flanks throat and belly. Iris is grey. See Figure 7.
Advertisement call. Typical call represents the only example of a clearly biphasic call within the genus Callulina ; it is formed by an initial whistle consisting of a single unpulsed long note with two peaks of amplitude followed, after a brief interval, by a single pulse train and, after another interval, by a group of similar 3 to 10 pulse trains. The typical call is usually reached after a series of ‘pre-calls’ consisting of the whistle followed by a single or double pulse train. Pre-calls can be repeated 3 to 5 times. These observations are based on recordings of 9 calls at the type locality, estimated to be from 4 individuals. Parameters are given as mean, (range), and standard deviation. Typical calls consist of a whistle that starts at 1.69 kHz (1.66–1.72) SD 0.02, rises and ends at 2.1 kHz (2.09–2.23) SD 0.51. Whistle duration is 0.2 s (0.18–0.25) SD 0.02. The whistle is followed by a series of 3 to 10 pulse trains, each train consisting of four or five pulses. The single pulse train duration is 0.08 s (0.074–0.096) SD 0.007, with a pulse rate of about 7/8 pulses per second. See Figure 8.
Natural history. The holotype was collected from a branch above head height, at a place where several other individuals were calling but not located. The paratype was discovered cryptically hiding in the axils of leaves in primary submontane forest (see Figure 9 View FIGURE 9 ). The species was not heard calling from the ground, and the expanded fingertips might suggest a strictly arboreal habit.
Tibia - SUL TD – SUL TD – ETD ETD – SUL WDTF3 –WDF3
C. dawida = 21
Conservation status. Callulina kanga was collected in Kanga Forest Reserve at two localities at an elevation of 760m and 1140m ( Figure 1 View FIGURE 1. a b; see also Menegon et al. 2008 Table 1 View TABLE 1 , Kanga Site 3 and FT Kanga Site 7). Kanga was surveyed by two separate teams (totalling around one month of survey time), during which specimens and calls were restricted to two small localities, despite searching many other locations and forest types, and at different altitudes. If the species is localised to this particular band of lower submontane forest then the species has an extremely narrow distributional range (740–1140m), and would be of high conservation concern. Even if the species occurred throughout Kanga FR (500–2019m – an unlikely range for Afromontane frogs) with surveys simply failing to record or collect specimens, the maximum estimated of occurrence would be equal to or less than 66.64 km 2. This estimated area of occurrence would still be small enough for C. kanga to qualify as critically endangered (CR B1b (iii)) under IUCN criteria. Although only two specimens were recovered, many animals were heard calling, and population density remains to be quantified.
Etymology. The species is named after the forest reserve ( Kanga ) where the type series was collected and the species appears to be restricted to. The specific epithet is considered to be a noun in apposition.
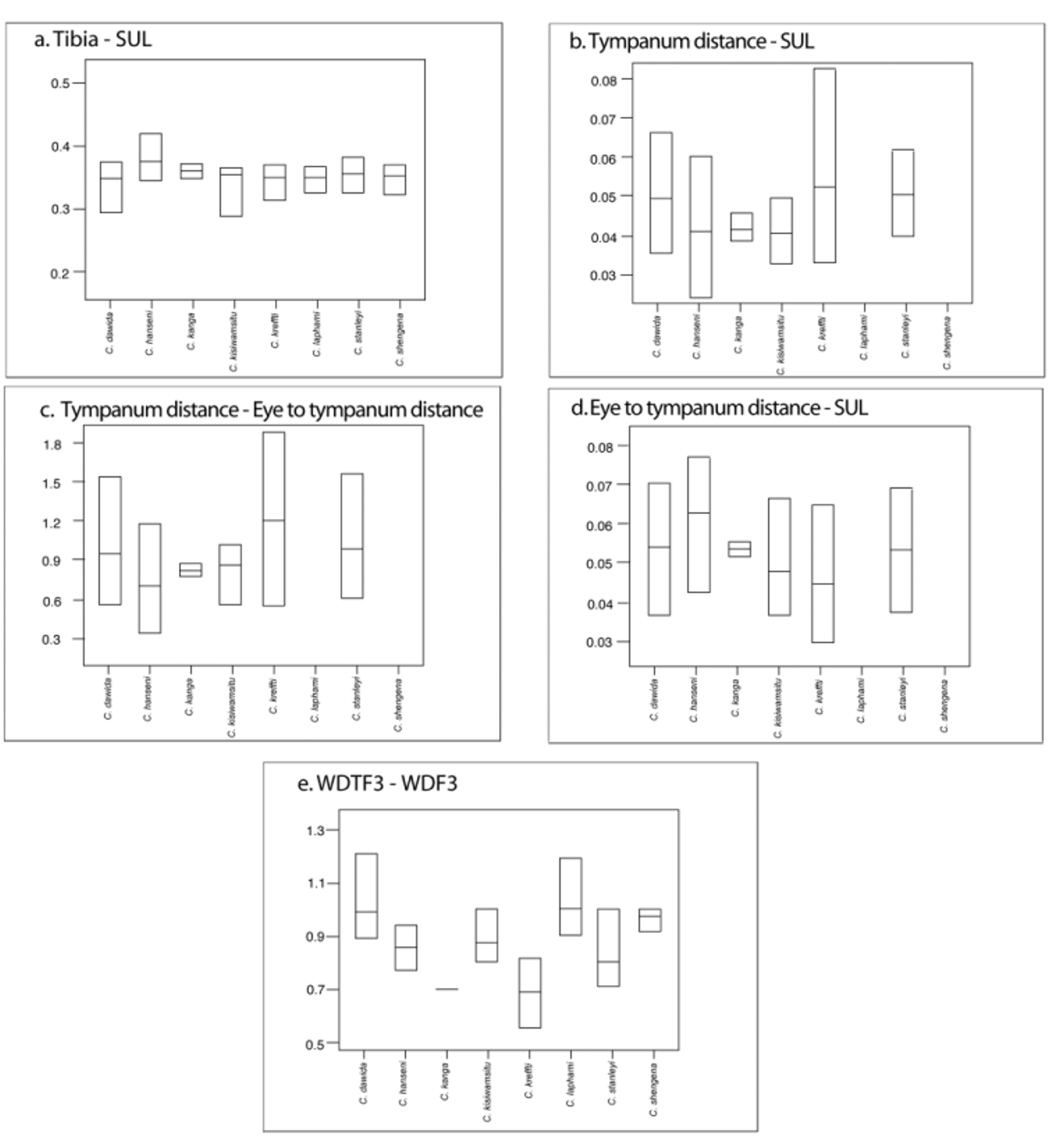
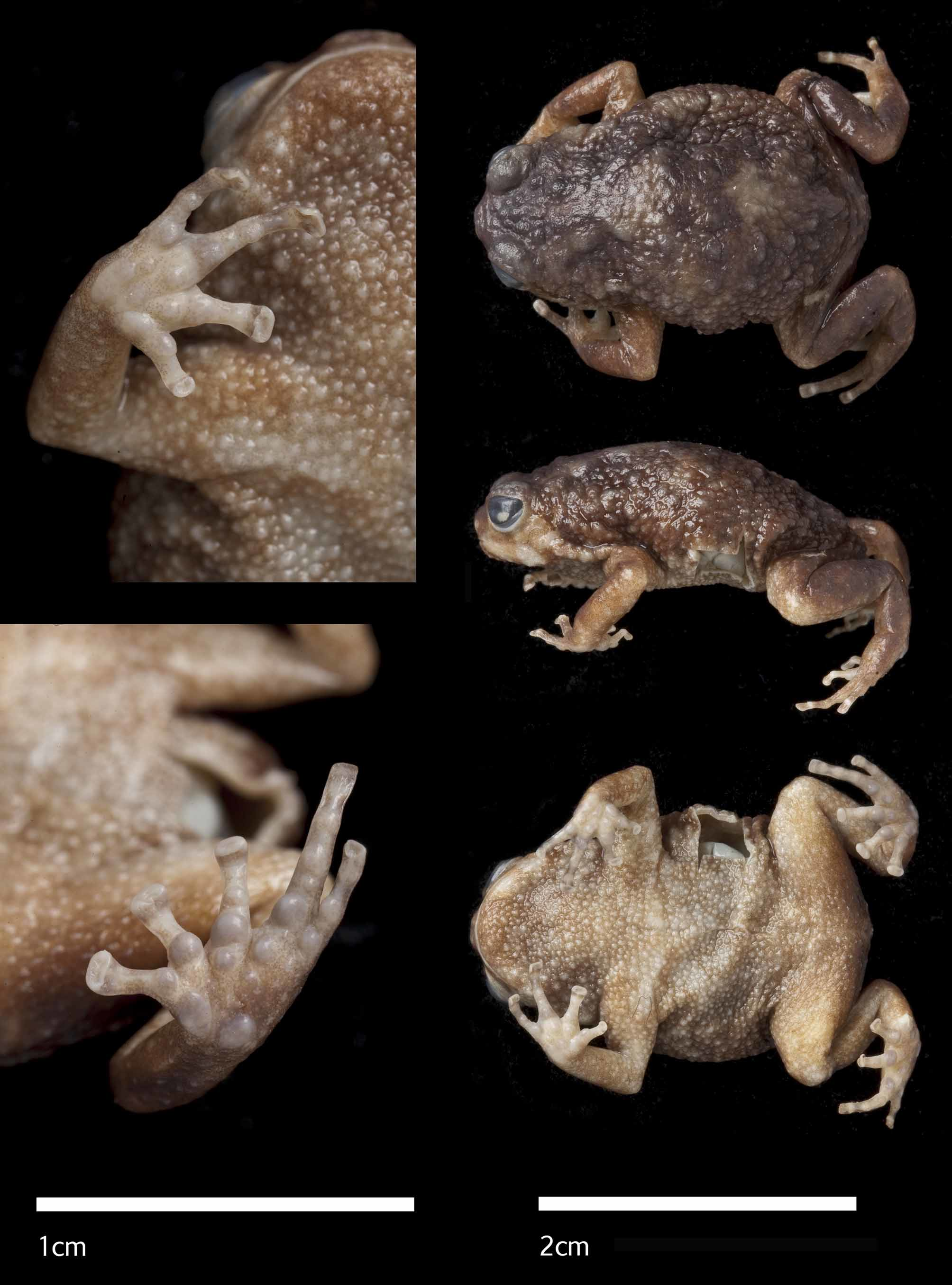


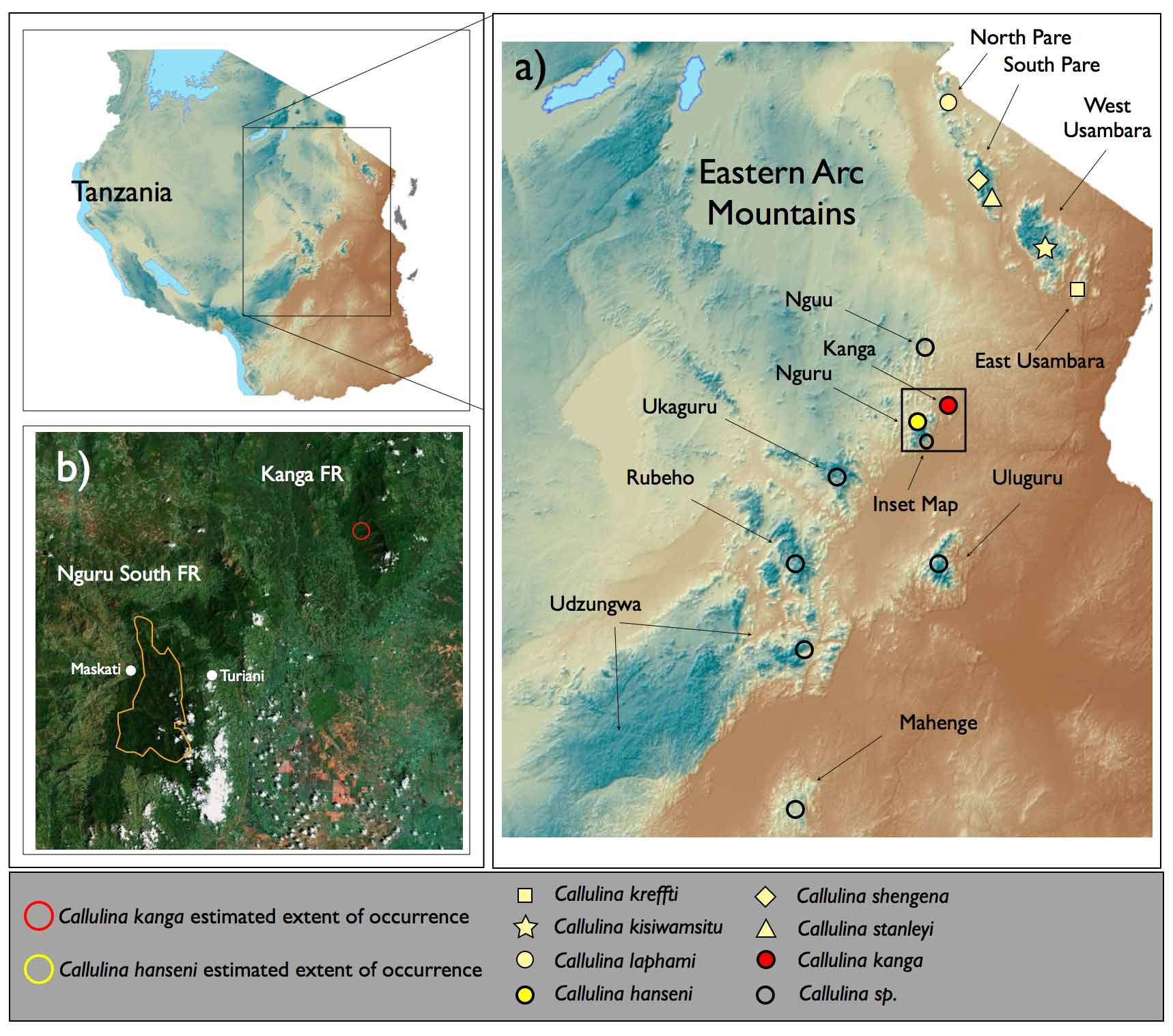
TABLE 2. Comparison of key morphometric characters across nominate species of Callulina (see Figure 4).
| Min | 0.297 | 0.0368 | 0.583 | 0.0366 | 0.9 |
|---|---|---|---|---|---|
| Max | 0.369 | 0.0671 | 1.556 | 0.0709 | 1.2 |
| Average | 0.346 | 0.0493 | 0.997 | 0.0538 | 0.996 |
| C. hanseni = 14 | |||||
| Min | 0.361 | 0.0259 | 0.355 | 0.0433 | 0.778 |
| Max | 0.424 | 0.0603 | 1.1875 | 0.0787 | 0.938 |
| Average | 0.385 | 0.0413 | 0.690 | 0.0625 | 0.855 |
| C. kanga = 2 | |||||
| Min | 0.367 | 0.0390 | 0.750 | 0.0519 | 0.7 |
| Max | 0.381 | 0.0464 | 0.846 | 0.0549 | 0.7 |
| Average | 0.374 | 0.0427 | 0.798 | 0.0534 | 0.7 |
| C. kisiwamsitu = 24 | |||||
| Min | 0.288 | 0.0330 | 0.583 | 0.0360 | 0.8 |
| Max | 0.379 | 0.0497 | 1.0 | 0.0663 | 1.0 |
| Average | 0.351 | 0.0407 | 0.841 | 0.0486 | 0.889 |
| C. kreffti = 58 | |||||
| Min | 0.318 | 0.033 | 0.545 | 0.0305 | 0.579 |
| Max | 0.376 | 0.082 | 1.875 | 0.0639 | 0.818 |
| Average | 0.346 | 0.053 | 1.223 | 0.0448 | 0.695 |
| C. laphami = 21 | |||||
| Min | 0.328 | - | - | - | 0.909 |
| Max | 0.370 | - | - | - | 1.2 |
| Average | 0.346 | - | - | - | 1.0108 |
| C. stanleyi = 11 | |||||
| Min | 0.328 | 0.04 | 0.607 | 0.0378 | 0.7143 |
| Max | 0.390 | 0.0638 | 1.571 | 0.0708 | 1.0 |
| Average | 0.358 | 0.0506 | 1.0114 | 0.0532 | 0.817 |
| C. shengena = 12 | |||||
| Min | 0.329 | - | - | - | 0.917 |
| Max | 0.377 | - | - | - | 1.0 |
| Average | 0.356 | - | - | - | 0.980 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.