
Callulina hanseni, Loader, Simon P., Gower, David J., Müller, Hendrik & Menegon, Michele, 2010
|
publication ID |
https://doi.org/10.5281/zenodo.199640 |
|
DOI |
https://doi.org/10.5281/zenodo.5616311 |
|
persistent identifier |
https://treatment.plazi.org/id/03C99D08-FFCC-8777-269B-22F4FEB3FE59 |
|
treatment provided by |
Plazi |
|
scientific name |
Callulina hanseni |
| status |
sp. nov. |
Callulina hanseni View in CoL sp. nov.
Figures 1 View FIGURE 1. a , 2 View FIGURE 2 , 3, 4 View FIGURE 4 ; Tables 1 View TABLE 1 , 2 View TABLE 2 , 3 View TABLE 3 .
Callulina sp. 2 Menegon et al. (2008) p.114, appendix 1, and table 3, 4.
Holotype. BMNH 2008.130 (Field Tag MW 6960), an adult female, BMNH 2008.17. Collected on the Maskati side of the Nguru South Forest Reserve, Tanzania, 06º 03' 51.1"S, 37º 30' 33.3"E, 1790 m ( Figure 1 View FIGURE 1. a b) by David Gower, Roy Hinde, Simon Loader, Hendrik Müller, Maria Perkins, and Mark Wilkinson on January 19th 2008.
Paratypes. 13 specimens: BMNH 1983.45 collected near Maskati Mission 1900m West side of Nguru Mountains, Mwomero District, Morogoro Region, Tanzania by Jan Keilland in Sept. 1982. BMNH 2008.127- 128, 2008.131-134, 2008.136-138 collected on the Maskati side of the Nguru South Forest Reserve, Mwomero District, Morogoro Region, Tanzania, 06º 03' 51.1"S, 37º 30' 33.3"E, 1790 m by David Gower, Roy Hinde, Simon Loader, Hendrik Müller, Maria Perkins, and Mark Wilkinson between January 19th–21st 2008. MTSN 8138, 8140 and 8192 collected on the Maskati side of the Nguru South Forest Reserve, Mwomero District, Morogoro Region, Tanzania, 06º 03' 51.1"S, 37º 30' 33.3"E, 1790 m by Michele Menegon between October 26th–30th 2004 (see Menegon et al. 2008, Table 1 View TABLE 1 , Nguru Site 1 ‘Maskati’). BMNH 2008.136 and MTSN 8138 and 8140 have been sequenced for partial fragments of 12S, 16S, and cytb (see Genetic difference section and Appendix 1).
Diagnosis. The new species of Callulina is assigned to the brevicipitid genus based on the following characteristics: Truncated or expanded terminal phalanges (simple in Spelaeophyrne, Probreviceps , Breviceps , and Balebreviceps ); single posterior denticulated row in the palate of Callulina (two denticulated rows in Probreviceps , glandular mass in Breviceps ).
A large, robust Callulina . SUL 20.6– 42.5mm. TL: SUL ratio 36–42%. Tympanum present, 0.25– 0.60% of SUL. Tympanum to eye distance 1.2 – 3.0 mm, 0.04–0.07 % of SUL. Fingertips expanded (width of subarticular tubercle>0.78–0.94 % of the width of fingertips). Dark brown dorsally, and ventrally, with distinctive darker brown colouration on anterior margin of chin against cream colouration. Legs have large continuous glandular ridge on both tibiofibulae and tarsal joints.
Callulina hanseni differs from C. laphami and C. shengena in the presence of a tympanum, more granular skin, expanded fingertips, and absence of interocular patterning. Callulina hanseni differs from C. dawida in having expanded fingertips. Callulina hanseni differs from C. stanleyi and C. kisiwamsitu in the presence of large continuous glands on legs. Callulina hanseni differs from C. kreffti in the degree of expansion of fingertips and the presence of large continuous glands on legs (see also Table 2 View TABLE 2 and Figure 4 View FIGURE 4 for comparative measures). The distinctiveness of C. hanseni from other Callulina species is also supported by its disjunct distribution, and mtDNA sequence data (see Genetic difference).
The extent of the expansion of the fingertips is, in general, a useful character to distinguish Callulina species but some specimens of some species fall outside the main range of variation and might question the usefulness of this character. Some specimens of C. kreffti (BMNH 2000-189, 2000-196, 2005.1508, FMNH 250471) have> 0.75% width at first subarticular, relative to expansion at fingertip. Also C. stanleyi FMNH 251384 has larger expansion of finger tips (<0.75) than characterised for this species. Expanded fingertips are highly susceptible to dehydration, such that preservation can alter measurements substantially, and caution needs to be applied when using this character to distinguish between species.
Description of holotype. (see Figure 2 View FIGURE 2 ) Body robust. Tips of fingers expanded, distal width of terminal phalanx 0.83 of the width of distal subarticular tubercle. Tips of fingers expanded slightly laterally, with circummarginal grooves posterior to distal phalanx. The distal phalanx is expanded beyond the width of the base of the phalange, toe tips ‘Y’ shaped. First finger shortest, second and fourth equal, third longest. Inner metatarsal tubercle rounded and raised, separated marginally at the point of contact on the edge of the palm by a wedge shaped medial palmar tubercle from a larger outer metatarsal tubercle. Slightly smaller palmar tubercles present on palm. Large subarticular tubercles at the base of each finger. Third and fourth finger with smaller tubercles between basal articular tubercle and subarticular tubercles. Dorsoventrally expanded toe tips, edged with longitudinal folds on a smooth ventral surface. Tips of toes very slightly expanded laterally, with circummarginal grooves posterior to distal phalanx. Slightly smaller palmar tubercles present on palm. Large subarticular tubercles at the base of each finger. Third and fourth finger with smaller paired tubercles between basal articular tubercle and subarticular tubercles. Expanded toe tips with smooth ventral surface with longitudinal folds. First toe approximately same length as second, third and fifth equal and longer than first and second toe, fourth longest. Inner metatarsal tubercle large, rounded and raised, almost touching a smaller sized, rounded, raised, outer metatarsal tubercle. Smaller palmar tubercles present on base of foot. Subarticular tubercles at the base of each toe, large subarticular tubercles on third, fourth and fifth toes at the phalangeal joints. All tubercles on hands and feet grey/blue colour against light and dark brown background. Skin very granular throughout. Large continuous raised glandular mass on arms, broken at elbow joint, and tapering onto the dorsal surface of the wrist. Large continuous glandular masses on each part of the leg joint, tapering to the dorsal surface of the posterior margin of the foot.
Canthus rostralis rounded, loreal region sloping slightly concavely. Loreal and canthal regions dark brown. Snout just visible in ventral view. Nostrils rounded, nearer to tip of snout than eye. Interorbital space flat, greater than width of eyelid. Pupil horizontal, iris grey. Tympanum present but small and difficult to distinguish because of the granular skin surface. Tympanum ovoid, shorter transversely than longitudinally. Tympanic region slightly paler brown/cream colour. Dark edged glandular tympanic ridge extends from posterior edge of eyelid to arm insertion. Tongue rounded. Single posterior denticulated row on the palate. Vent posteroventrally positioned.
Measures of holotype. ED = 5.0; ETD = 3.1; HL = 13.6; IOD = 7.6; JW = 15.1; LF3 = 5.9; LT4 = 7.0; ND = 2.9; NED = 3.5; NLD = 1.9; SUL = 42.5; TD = 1.1; TL = 15.9; TSL = 11.9; WDF3 = 1.8; WDTF3 = 1.5.
C. hanseni (n = 14) C. kanga (n = 2)
Morphological and colour variation. In preservation holotype and paratype specimens generally similar in their colouration, with a dark brown dorsum, slightly paler brown laterally. Smaller specimens (mainly males) with paler brown/grey colouration and dark brown/black mottling. BMNH 2008.132 is paler. MTSN specimens have much paler colouration overall likely due to different preservation procedure (fixed in ethanol rather than formalin). The dark colouration of BMNH 1983.45 seems to have slightly faded over time. Venter in all paratypes is pale brown with darker patches on lateral sides. Distinctive darker brown patch on anterior margin of chin, present in all specimens. This distinctive chin patterning is darker in some specimens (BMNH 2008.137, 131, 138, MTSN 8192).
Morphometric analyses indicate sexual dimorphism in C. hanseni . Based on analyses of eight females and six males, females are larger (SUL, student t-test => 0.00096), although relative proportions of limbs are not significantly different (tibia, tarsus, and humerus, student t-test = <0.05). As in C. dawida , the position and size of the tympanum differ between males and females. Males have a relatively larger tympanum, for example the TD: SUL ratio (student t-test => 0.048); and the position of the tympanum relative to the eye (ttest => 0.046) is significantly different.
Colour in life. A brown or dark brown animal with scattered black mottlings (especially in smaller animals) and white tipped warts on flanks, throat and belly. Wide paler dorsolateral bands can be present. Iris is bright orange. See Figure 3.
Advertisement call. No calls were recorded for this species.
Natural history. The specimen collected by Jan Keiland in 1982 was found inside a rotting log in moist evergreen forest. The new series collected in 2004 (MTSN) and 2008 (BMNH) were mainly collected from branches both below and above head height. This included specimens located low down on small shrubs but also climbing to a great heights (observed and collected ca. 10 m above ground) on branches, tree trunks and bare rock in montane forest (see Figure 9 View FIGURE 9 ). All specimens were collected in primary montane rainforest.
Conservation status. Callulina hanseni has been found only in the Nguru South Forest Reserve at an elevation of 1790m and above ( Figure 1 View FIGURE 1. a c). This distribution comprises a maximum distributional area of less than 100km 2, qualifying this species as critically endangered (CR B1b (iii)) under IUCN criteria. The conservation status will need to be re-evaluated if specimens are discovered below 1790m. Population density is unknown. The distribution pattern is similar to that seen in the recently described species Arthroleptis nguruensis ( Poynton et al. 2008) , and is indicative of a distinct Nguru upper montane fauna. Other undescribed taxa also appear to be confined to this upper montane belt (e.g. Callulina sp. Hoplophryne sp., and Probreviceps sp.).
Etymology. The species name is a patronym for Dr. James Hansen, who has made important scientific contributions towards climate science and as a supporter of the African Rainforest Conservancy trust has contributed towards the conservation of Eastern Arc forests. The specific epithet should be treated as a noun in genitive case.

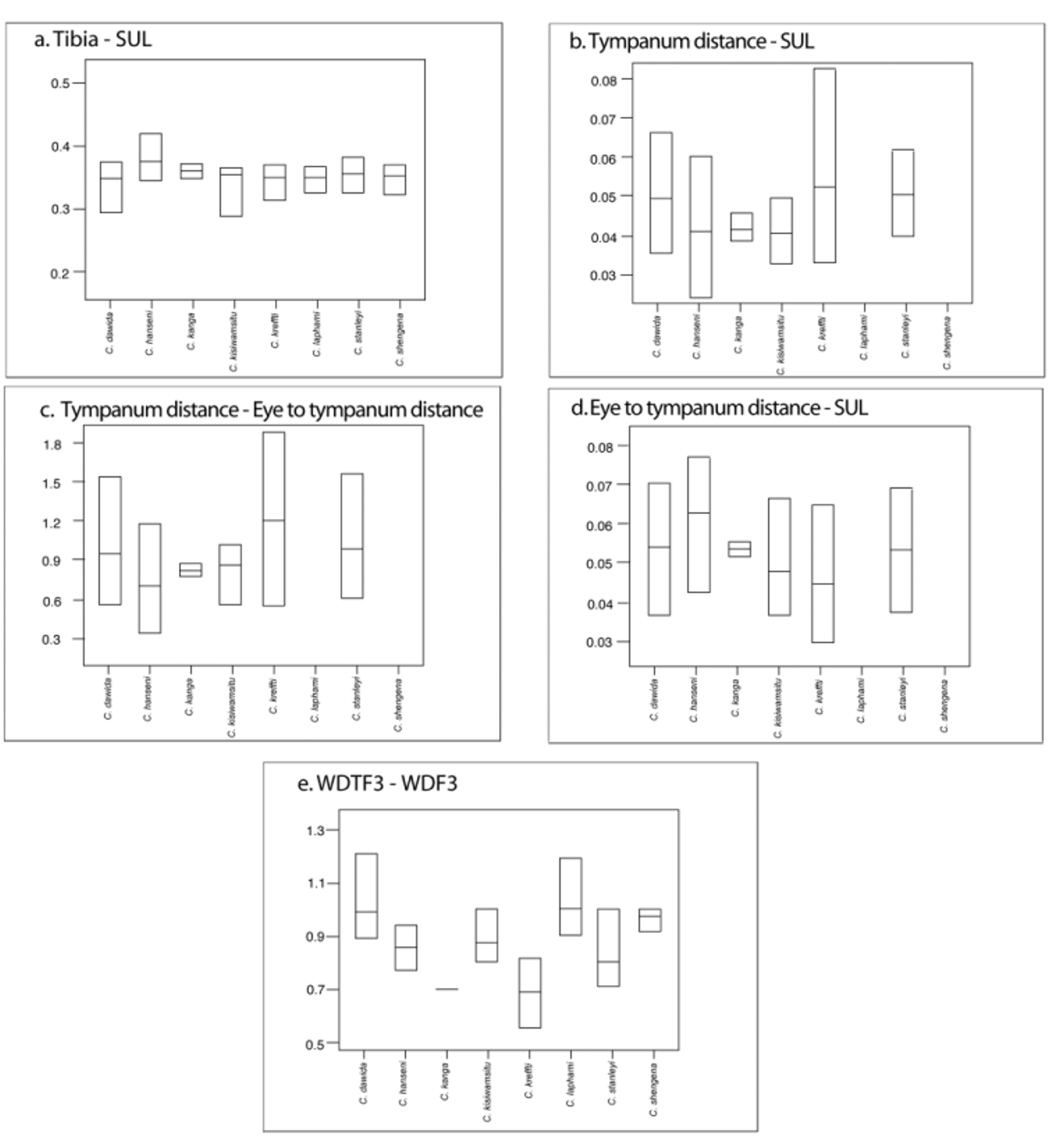

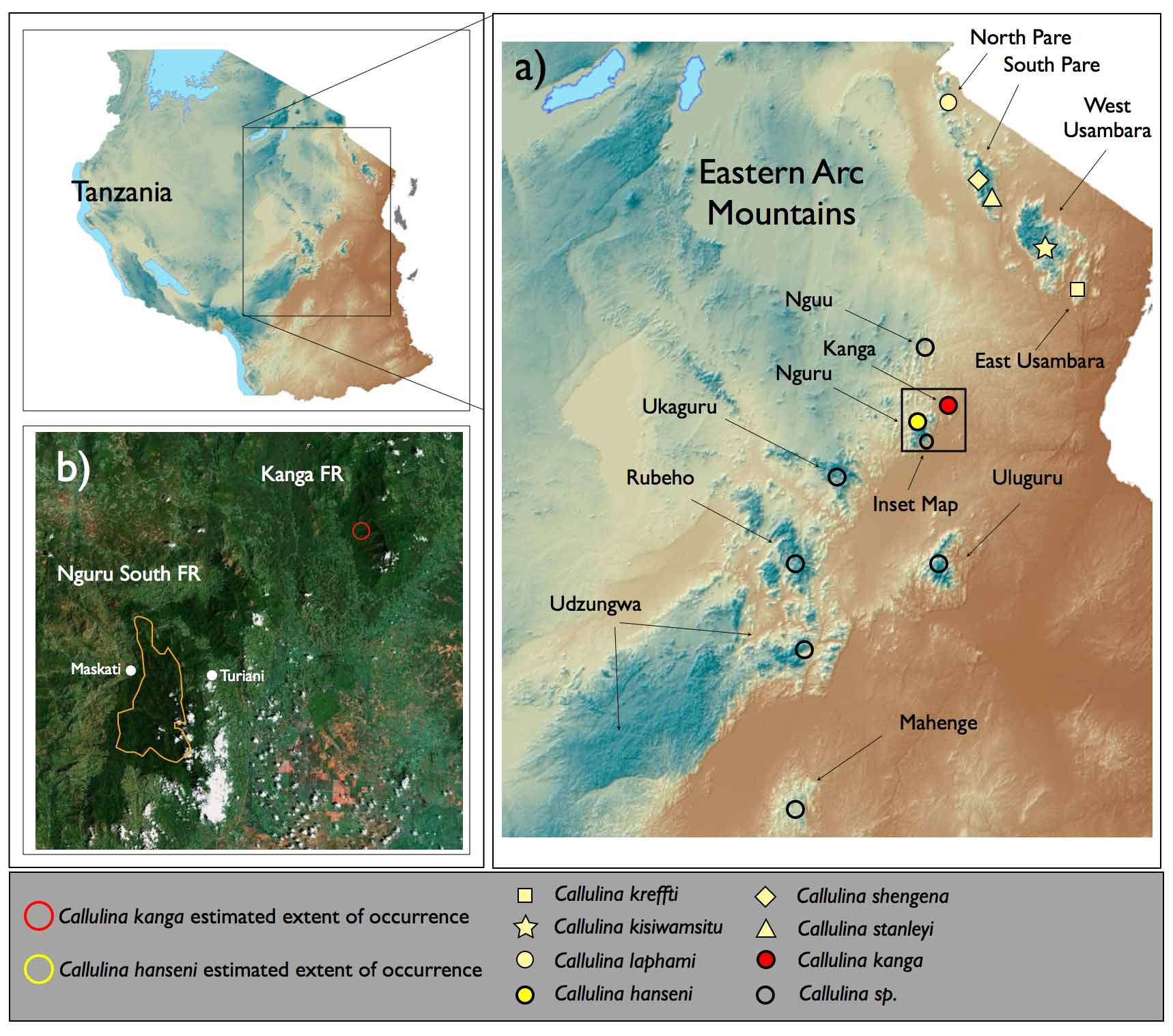
TABLE 1. Morphometric data for the two new species of Callulina, all measurements in mm. See Materials and Methods for explanation of abbreviations.
| Min. | Max. | Mean | Min. | Max. | Mean | |
|---|---|---|---|---|---|---|
| SUL | 20.6 | 42.5 | 30.7 | 23.1 | 23.7 | 23.4 |
| TL | 8.6 | 15.7 | 11.9 | 8.7 | 8.8 | 8.8 |
| TD | 0.9 | 1.9 | 1.4 | 0.9 | 1.1 | 1.0 |
| ETD | 1.2 | 3 | 1.9 | 1.2 | 1.3 | 1.3 |
| ED | 2.7 | 4.8 | 3.8 | 2.8 | 2.9 | 2.9 |
| ND | 1.6 | 2.8 | 2.1 | 1.4 | 1.6 | 1.5 |
| NED | 2 | 3.3 | 2.6 | 2 | 2.1 | 2.1 |
| HW | 7.6 | 15 | 10.6 | 7.4 | 7.6 | 7.5 |
| LF3 | 2.9 | 6.1 | 4.5 | 2.9 | 3 | 3.0 |
| LT4 | 3.7 | 7.5 | 5.8 | 3.7 | 3.9 | 3.8 |
| TSL | 5.8 | 12.1 | 8.8 | 5.9 | 6.1 | 6.0 |
| HL | 6.3 | 13.1 | 9.7 | 7 | 7.4 | 7.2 |
| NLD | 1.1 | 1.8 | 1.5 | 1 | 1 | 1.0 |
| WDF3 | 0.9 | 1.8 | 1.3 | 1 | 1 | 1.0 |
| WDTF3 | 0.8 | 1.5 | 1.1 | 0.7 | 0.7 | 0.7 |
| IOD | 4.2 | 7.4 | 5.9 | 4.7 | 4.7 | 4.7 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.