
Bythaelurus vivaldii, Weigmann, Simon & Kaschner, Carina Julia, 2017
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4263.1.4 |
|
publication LSID |
lsid:zoobank.org:pub:68B3A67E-5DB1-4552-9465-108895ECBBC1 |
|
DOI |
https://doi.org/10.5281/zenodo.6033252 |
|
persistent identifier |
https://treatment.plazi.org/id/3B508794-FFC7-FFB8-22A7-6AC111C0080F |
|
treatment provided by |
Plazi |
|
scientific name |
Bythaelurus vivaldii |
| status |
sp. nov. |
Bythaelurus vivaldii View in CoL n. sp.
Vivaldi’s Catshark
Figures 2–10 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9 View FIGURE 10 ; Table 1
Scylliorhinus hispidus: Brauer, 1908: 7 View in CoL –8 (in part).
Halaelurus lutarius: Springer & D’Aubrey, 1972: 6 View in CoL –8 (in part); Bass et al., 1975: 11 –12 (in part); Springer, 1979: 82 –83 (in part).
Bythaelurus View in CoL sp.: Kaschner et al., 2015: 121, 132, 136 (collection numbers of both specimens interchanged); Weigmann et al., 2016: 427, 429.
Holotype ZMB 17410, presumably adult female, 325 mm TL, RV ‘ Valdivia’ , station 265, off Somalia, 6°24'1" N, 49°31'6" E, 628 m depth, 30 Mar 1899. GoogleMaps
Paratype ZMB 22423, presumably immature female, 272.7 mm TL, data the same as holotype.
Diagnosis. A supposedly very small scyliorhinid species with the following characteristics: body firm and stout in presumably adult holotype (slender in supposedly juvenile paratype); snout long (preorbital length 6.1– 6.6% TL) and broad, bell-shaped in dorsoventral view with distinct lateral indention; pre-outer nostril length subequal to internarial space; preorbital snout length subequal to interorbital space; preoral snout length 1.4–1.8 times in mouth width; eye length 13.9–15.0 times in predorsal distance, 6.8–6.9 times in head length and 1.6–2.3 times eye height; head length 2.6–2.8 times width at level of maximum outer extent of anterior nostrils; head width at level of maximum outer extent of anterior nostrils 1.2 times width at level of lateral indention of head, 1.3–1.4 times preorbital length, and 8.5–8.6% TL; roof of mouth loosely set with rather small, knob-like to elongated oral papillae, oral papillae on tongue hardly detectable; pelvic-fin anterior margin 2.0–2.1 times in pectoral-fin anterior margin; first dorsal-fin base 2.2–2.3 times in interdorsal space; length of second dorsal-fin inner margin 1.3–2.3 times in second dorsal-fin height; second dorsal-fin base length 5.3% TL; anal-fin base 0.7–0.8 times interdorsal space. Coloration: reported to have 8–9 dark, broad but inconspicuous transverse bars on the back, but preserved coloration plain beige with scattered remains of dark brown. Upper jaw with 68–75 and lower jaw with 64 rows of small tri- to pentacuspidate teeth with outer surface of crown furrowed by strong longitudinal ridges from base of cusps to tip and strongly structured by reticulations from basal areas to well into cusps; monospondylous trunk vertebrae centra 38, diplospondylous precaudal centra 43–44, total centra about 135–140. Dermal denticles leaflike to teardrop-shaped on snout and tricuspidate on trunk and tail. The new species is distinguished from all congeners in the western Indian Ocean by a larger pre-second dorsal fin length, a longer head, a larger interdorsal space, a larger intergill length, a longer pectoral-fin posterior margin, a shorter caudal fin, an intermediate caudal fin preventral margin, and a larger internarial width.
Description of the holotype ( Figures 2–10 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9 View FIGURE 10 ). Values of the paratype in parentheses if differing. Morphometric measurements and meristics are given in Table 1.
B. vivaldii n. sp. B. vivaldii n. sp. presumably presumably adult female immature female paratype holotype ZMB 17410 ZMB 22423
B. vivaldii n. sp. B. vivaldii n. sp. presumably presumably adult female immature female paratype holotype ZMB 17410 ZMB 22423
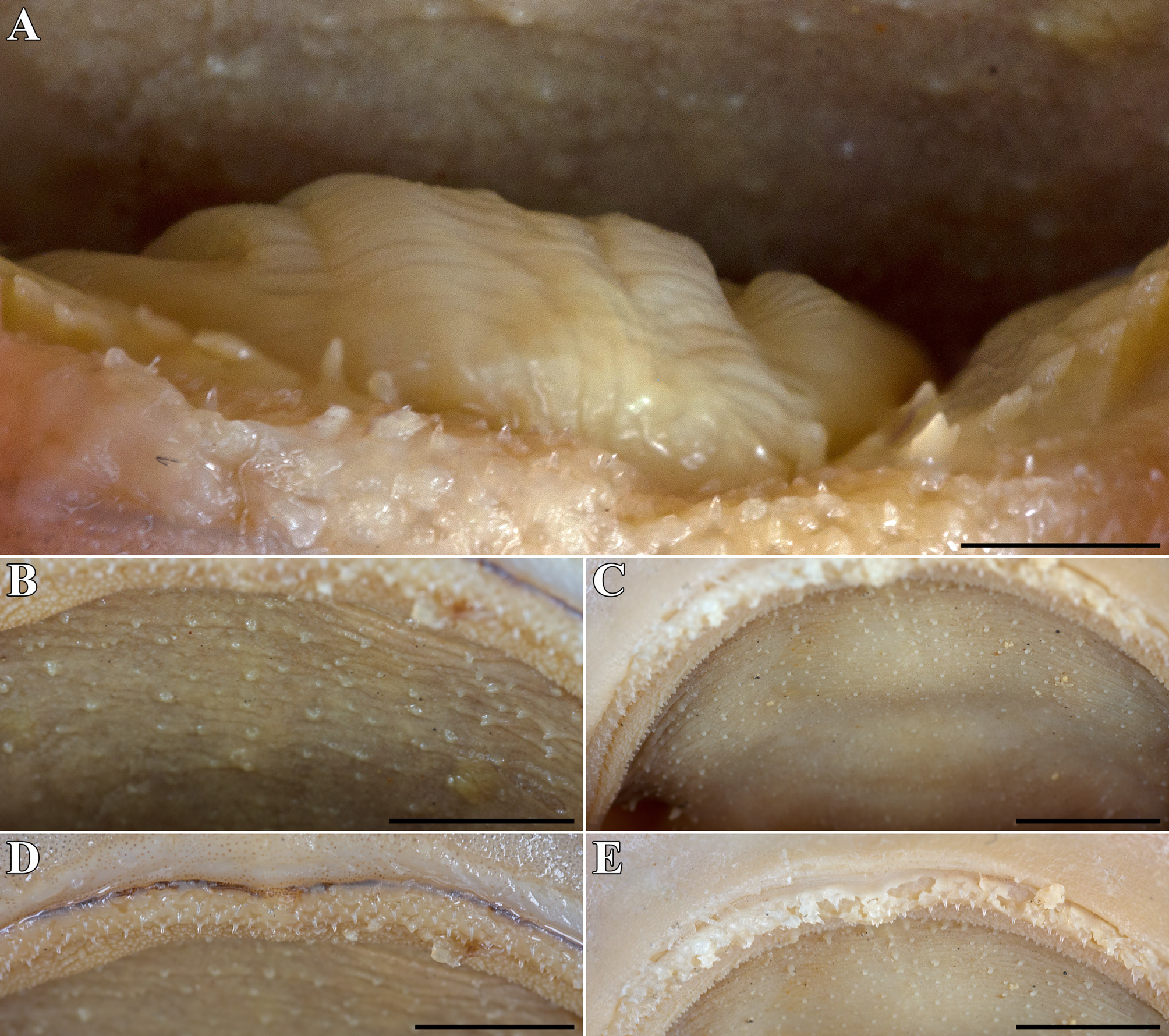
B. vivaldii n. sp. B. vivaldii n. sp. presumably presumably adult female immature female paratype holotype ZMB 17410 ZMB 22423 External morphology. Body firm and stout with trunk width 11.0% TL (rather slender with trunk width 9.4% TL in presumably juvenile paratype), subcircular in cross section at mid-trunk, laterally compressed and tapering posterior to cloaca; head region broad, long abdominal and caudal sections ( Figures 2–4 View FIGURE 2 View FIGURE 3 View FIGURE 4 ). No predorsal, interdorsal, or postdorsal ridges; no postanal ridge; no lateral ridges on caudal peduncle. Trunk slightly shorter than tail, distance from tip of snout to anterior cloaca 45.8% (44.8%) TL; pre-first dorsal fin length 48.2% (49.6%) TL, pre-second dorsal fin length 67.0% (67.9%) TL, ventral precaudal length 73.5% (73.9%) TL. Head broad and dorsoventrally flattened, with a broadly rounded snout; laterally uncompressed (slightly compressed) in gill region ( Figures 5–6 View FIGURE 5 View FIGURE 6 ); no supraorbital crest on chondrocranium; head length 2.8 (2.6) times width at level of maximum extent of anterior nostrils and 1.6 (1.3) times pectoral - pelvic space; head width at level of maximum outer extent of anterior nostrils 1.2 times width at level of lateral indention of head, 1.3 (1.4) times preorbital length, and 8.5% (8.6%) TL; head width at posterior edge of nostrils 1.4 and at mouth corners 1.9 (1.8) times width at level of lateral indention of head; head width at middle gill slits 1.8 (1.5) times width at level of lateral indention of head. Snout long and broad, its tip broadly rounded, strongly bell-shaped in dorsoventral view with distinct lateral indention; pre-outer nostril length 1.1 (1.0) times internarial width and 0.4 times interorbital width; preoral length 0.7 (0.6) times mouth width and 1.0 (0.9) times preorbital length; preorbital length 3.6 (3.7) times in head length and 0.9 times interorbital space. Eyes rather small and elongated, dorsolaterally on head, eye length 6.9 (6.8) times in head length, 13.9 (15.0) times in predorsal distance, and 2.3 (1.6) times eye height; nictitating lower eyelids, anterior and posterior eye notches, and suborbital groove present ( Figures 5–6 View FIGURE 5 View FIGURE 6 ). Spiracles large and slit-like, close behind but well separated from eyes, dorsolaterally on head and somewhat lower than level of eye notches, spiracle length 4.0 (4.4) times in eye length and 9.0 times in interorbital width. Gill slits moderately long, well separated, their upper ends clearly below lower edge of eye; gill area fully scaled, gill filaments not visible externally; gill openings increasing in size from first to fourth, the height of the fifth, which is above pectoral-fin origin, is subequal to that of the second one. Nostrils oblique, expanding diagonally inwards from snout edge, clearly not reaching level of mouth, with large, triangular anterior nasal flaps and much smaller but still distinct posterior flaps; pre-outer nostril length 1.1 (1.0) times nostril width and 0.5 (0.6) times preoral snout length, nostril width 1.0 (0.9) times internarial width and 0.9 times eye length ( Figures 5–6 View FIGURE 5 View FIGURE 6 ). Mouth very broad, width 1.4 (1.8) times preoral length, 0.7 times head width at mouth corners, 2.6 (2.3) times in head length, and 3.7 (2.7) times mouth length. Upper and lower labial furrows well developed, upper ones not reaching midpoint between mouth corner and posterior margin of nostril, lower furrows 2.0 (1.6) times as long as upper ones. Tongue moderately long, flat and rounded, lightcolored, with oral papillae hardly detectable ( Figure 7 View FIGURE 7 , A). Entire roof of mouth loosely set with rather small, knoblike to elongated papillae ( Figure 7 View FIGURE 7 , B–C); fleshy buccal curtain along inner margin of upper and lower jaws very densely set with small, knob-like to elongated papillae ( Figure 7 View FIGURE 7 , D–E).
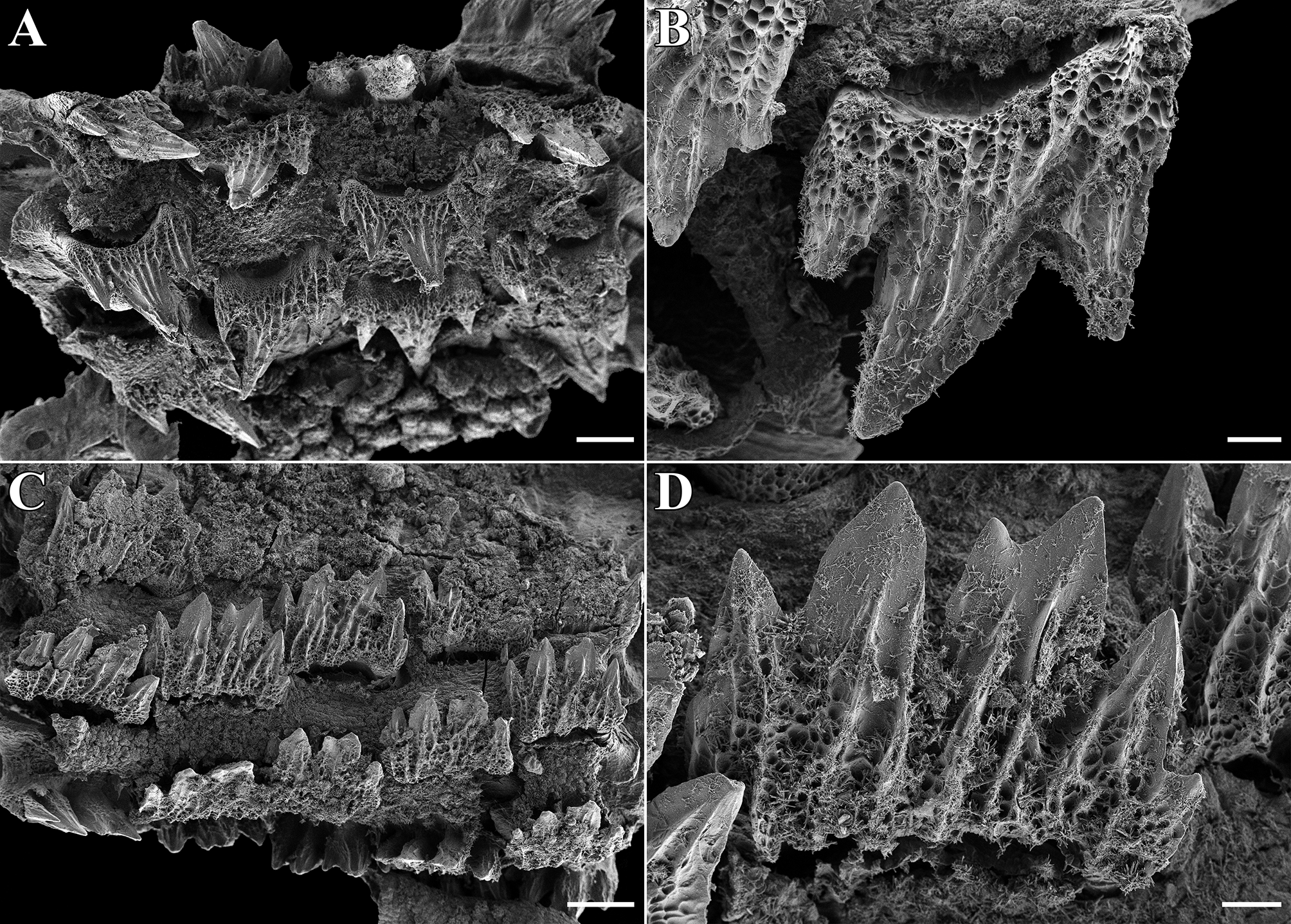
Upper jaw with 68 (about 75) and lower jaw with 64 (not counted due to jaw damage) diagonal rows of small teeth. Anterolateral teeth in upper jaw tricuspidate, partially with small additional cusplets, with median cusp much longer than lateral cusps ( Figure 8 View FIGURE 8 , A–B); posterolateral teeth in upper and lower jaws tri- to pentacuspidate and with median cusp only slightly longer than lateral cusps. Anterolateral teeth in lower jaw ( Figure 8 View FIGURE 8 , C–D) similar to posterolateral teeth. Outer surface of crown furrowed by strong longitudinal ridges from base of cusps to tip and strongly structured by reticulations from basal areas to well into cusps. Cutting edges of cusps without serrations.
Dermal denticles densely set and overlapping. Dermal denticles on dorsal and ventral ( Figure 9 View FIGURE 9 , A–B) snout leaf-like to teardrop-shaped, surface structured by reticulations in basal half, with four to six narrow ridges that do neither fuse nor reach the tip of the denticle. Denticles in branchial area, on lateral trunk ( Figure 9 View FIGURE 9 , C–D) and on lateral caudal fin ( Figure 9 View FIGURE 9 , E–F) tricuspidate, with long and pointed median main cusp and shorter, pointed lateral cusps at lower level, surface structured by reticulations in basal third, with or without short, narrow median ridge that, if present, does not reach the tip of the cusp and with two longer, narrow lateral ridges that do not fuse and reach or nearly reach the lateral edge of the median cusp and the tips of the lateral cusps, respectively. Dermal denticles on anterior dorsal caudal-fin margin ( Figure 9 View FIGURE 9 , G–H) slightly enlarged but otherwise similar to those in branchial area, on lateral trunk and on lateral caudal fin.
Pectoral fins subtriangular, non-falcate, anterior margin convex, its length 2.1 (1.8) times pectoral base length and 1.0 (0.9) times the length of the convex posterior margin, apex rounded, inner margin convex and 1.2 (1.0) times pectoral base length, inner pectoral corner broadly rounded ( Figures 5–6 View FIGURE 5 View FIGURE 6 ). Pectoral - pelvic space 1.3 (1.7) times length of pectoral-fin anterior margin and 1.1 (1.3) times interdorsal space.
Pelvic fins narrowly triangular with long, straight anterior and posterior margins and shorter straight inner margin, anterior margin half as long as pectoral-fin anterior margin; pelvic-fin origin clearly anterior to first dorsalfin origin, pelvic posterior tips below insertion of first dorsal fin ( Figure 10 View FIGURE 10 , A). Pelvic - anal space short, 10.7% (13.1%) TL and 0.7 (0.8) times pectoral - pelvic space.
First dorsal fin 1.8 (1.7) times higher than but about as long as second dorsal fin, anterior margin convex, apex rounded, posterior and inner margins straight, free rear tip angular; base length 1.2 (1.1) times fin height and 0.5 (0.4) times interdorsal space; first dorsal-fin origin behind level of pelvic-fin midbase ( Figure 10 View FIGURE 10 , A).
Second dorsal fin distinctly lower than first dorsal fin, anterior margin convex, apex rounded, posterior margin slightly convex, inner margin straight, free rear tip angular, base length 1.9 (1.7) times fin height and 0.4 times interdorsal space; second dorsal-fin origin over anal-fin midbase ( Figure 10 View FIGURE 10 , A).
Anal fin a long and low, uneven triangle, with long, straight anterior and posterior margins of about equal length and short inner margin, apex rounded, free rear tip pointed angular; base length 2.8 (3.1) times fin height, 0.8 (0.7) times interdorsal space, and 1.0 (0.7) times pelvic - anal space; base 1.9 (1.7) times longer than second dorsal-fin base and 6.6 (4.6) times anal - caudal space ( Figure 10 View FIGURE 10 , A). Anal-fin origin distinctly anterior to second dorsal-fin origin.
Caudal fin slender, short and strongly asymmetrical, its length 3.7 (4.2) times fin height and only 1.8 (1.9) times the interdorsal space; dorsal caudal margin weakly convex, no lateral undulations; upper caudal lobe very low, lower caudal lobe much deeper, with straight pre- and slightly concave postventral margins. Ventral caudal-fin origin far anterior of dorsal caudal-fin origin due to very long preventral margin, which is about as long as the postventral margin and forms a strongly elongated fleshy ridge in about anterior half of its length. Ventral corner bluntly angled; subterminal notch distinct; terminal lobe about 3.3 (4.4) times in caudal fin length; terminal caudal margin distinctly convex ( Figure 10 View FIGURE 10 , B).
Skeletal meristics (from radiographs, Table 1): monospondylous trunk vertebral centra: 38; diplospondylous precaudal centra: 44 (43); total precaudal centra: 82 (81); caudal centra (approximately): 58 (54); total centra (approximately): 140 (135). Spiral valve turns in paratype ZMB 22423: 7.
Coloration. Color in preservative: plain beige with scattered remains of dark brown, possibly representing relicts of the original, darker coloration (no remains of dark brown in paratype). Otherwise no spots, blotches or stripes detectable ( Figures 3–4 View FIGURE 3 View FIGURE 4 ). Brauer (1908) described the coloration of the ‘Valdivia’ specimens as follows: “graubraun auf dem Rücken, heller am Bauch. Auf dem Rücken einige, 8–9 dunkle, breite, aber wenig hervortretende Querbinden, welche nicht auf die ventrale Körperhälfte hinüberreichen. Durch diese Querbinden, welche von Alcock nicht erwähnt werden, ähnelt die Art Sc. quagga ( Alcock) ( Alcock 1899, p. 17, Taf. 27, fig. 1 ), aber die geringe Zahl der Querbinden und weiter die Lage der zweiten Rückenflosse unterscheidet diese Art von Sc. quagga .” Translated to English, the coloration was described by Brauer (1908) as “grayish-brown on the back, lighter on the belly. On the back, several, 8–9, dark, broad but inconspicuous transverse bars, which do not reach the ventral half of the body. Due to these transverse bars, which are not mentioned by Alcock, the species resembles Sc. quagga ( Alcock) ( Alcock 1899, p. 17, pl. 27, fig. 1 ), but the small number of transverse bars, as well as the position of the second dorsal fin distinguish this species from Sc. quagga .” Although Brauer correctly mentioned three specimens, it can be assumed that his detailed description (including that of the coloration) refers only to one or both specimens of Bythaelurus vivaldii , but not to the adult male holotype of B. tenuicephalus . This assumption is supported by the clearly differing preserved coloration that the holotype of B. tenuicephalus still shows, closely matching the fresh coloration of the paratype of B. tenuicephalus (see Kaschner et al. 2015). Furthermore, detailed measurements were provided by Brauer only for the largest of the three specimens, the female holotype of B. vivaldii , and both specimens of B. vivaldii were deposited in the ZMB, where Brauer was employed, whereas the holotype of B. tenuicephalus was deposited in the ZMH. A drawing showing the pigmentation pattern B. vivaldii reconstructed after the description by Brauer (1908) can be found in Figure 2 View FIGURE 2 . It remains unclear to what extent the fresh coloration of B. vivaldii indeed resembles that of Halaelurus quagga as no image was provided by Brauer (1908). Halaelurus quagga typically shows more than 20 clear and pronounced, very dark brown transverse bars of varying width dorsally from the snout tip to the tip of the caudal fin.
Size. A supposedly very small catshark, presumably maturing at around 30 cm TL and probably attaining about 35 cm TL. See under Remarks for additional comments on maturity staging.
Distribution. Known only from the upper slope off Somalia, 6°24'1" N, 49°31'6" E, in 628 m depth.
Etymology. The new species is named in honor of Antonio Vivaldi (1678–1741), a genius composer of Baroque era, to express its relationship to Bythaelurus bachi , named after sublime genius Johann Sebastian Bach. In the western Indian Ocean, B. vivaldii n. sp. is morphologically closest to B. bachi , in particular due to the stout body shape of large specimens and presence of oral papillae.
Remarks. The holotype has a stout body, whereas the smaller paratype has a rather slender body. Considering that stout body shape is pronounced only in adult specimens but not in juveniles in the morphologically closest congener in the western Indian Ocean, Bythaelurus bachi , it is assumed that the holotype of the new species is supposedly adult, whereas the paratype is presumably immature. Accordingly, a maturity size of females around 30 cm TL is supposed. Its immature reproductive organs evidence the maturity staging of the paratype. The maturity staging of the holotype, however, cannot be verified as its organs were removed. Based on a comparison of the relation between maturity size and maximum size in B. bachi , a maximum size of about 35 cm TL is assumed for the new species. This putative maximum size is based on an exceedance of maximum size vs. maturity size of 18% in females of its closest congener B. bachi , calculated based on the latter species’ maturity (about 37.7 cm TL) and maximum (about 44.5 cm TL) sizes of females as published by Weigmann et al. (2016). Based on its morphological similarity to B. bachi , it is supposed that B. vivaldii is also an oviparous species.
Discussion. Bythaelurus vivaldii n. sp. is morphologically closest to the recently described B. bachi , which is the only other Bythaelurus species in the western Indian Ocean that shares a stout body in large specimens and the presence of oral papillae. It further resembles B. vivaldii in the broad mouth and broad posterior head. Nevertheless, the new species differs from all congeners in the western Indian Ocean in a larger pre-second dorsal fin length (67–67.9% TL vs. 56.4–66.7% TL in B. bachi vs. 63.8–67.2% TL in B. naylori vs. 63.3% TL in B. clevai vs. 62.8% TL in B. hispidus vs. 65.9% TL in B. lutarius vs. 63.2–63.3% TL in B. tenuicephalus ), a longer head (23–24.1% TL vs. 17.6–23.3% TL vs. 13.6–18.7% TL vs. 21% TL vs. 21.7% TL vs. 21.3% TL vs. 19.9–22% TL), a larger interdorsal space (12.8–13.3% TL vs. 7.9–12% TL vs. 9–11.5% TL vs. 11.9% TL vs. 10.5% TL vs. 12.7% TL vs. 10.4–11.1% TL), a larger intergill length (5.6–6% TL vs. 2.5–5.3% TL vs. 2.6–5.5% TL vs. 3.3% TL vs. 4.7% TL vs. 4.3% TL vs. 3.7–4.3% TL), a longer pectoral-fin posterior margin (10.9–12.1% TL vs. 4.9–10.9% TL vs. 7.1–10.1% TL vs. 8.9% TL vs. 8.6% TL vs. 7.4% TL vs. 8.7–9.2% TL), a shorter caudal fin (23.8–24.4% TL vs. 25.5–37.4% TL vs. 28.8–33.7% TL vs. 30.1% TL vs. 29.5% TL vs. 26.6% TL vs. 27.9–30.3% TL), an intermediate caudal fin preventral margin (9.8–10.3% TL vs. 11.8–18.3% TL vs. 6.1–7.3% TL vs. 11.9% TL vs. 11.4% TL vs. 10.6% TL vs. 10.6–10.8% TL), and a larger internarial width (3.2% TL vs. 1.8–3.1% TL vs. 2.1– 2.8% TL vs. 3% TL vs. 3.1% TL vs. 2.6% TL vs. 2.7% TL). Furthermore, B. vivaldii has a smaller second dorsal fin than all congeners in the western Indian Ocean except for B. lutarius (total length 6.9–7.3% TL in B. vivaldii vs. 10–13.9% TL in B. bachi vs. 9.8–13% TL in B. naylori vs. 8.9% TL in B. clevai vs. 9% TL in B. hispidus vs. 8.1– 8.8% TL in B. tenuicephalus ; anterior margin length 5.1–7% TL vs. 9–13.5% TL vs. 9.1–12.5% TL vs. 8.3% TL vs. 8.7% TL vs. 7.7–8.7% TL; base length 5.3% TL vs. 6.7–9.7% TL vs. 6.8–8.6% TL vs. 6.9% TL vs. 6.8% TL vs. 5.9–6.9% TL). In contrast to its congeners in the western Indian Ocean with oral papillae, the tongue hardly bears any papillae in the new species (vs. tongue set with papillae of distinct size) and the papillae on the roof of mouth are smaller and rather loosely set. Additionally, the reported coloration of 8–9 dark, broad but inconspicuous transverse bars on the back would be a strong distinctive feature from all described congeners worldwide, but as described under Coloration the fresh coloration cannot be confirmed anymore as the preserved coloration is plain beige with scattered remains of dark brown.
In addition to the aforementioned differences to all congeners in the western Indian Ocean, Bythaelurus vivaldii n. sp. is distinguished from B. bachi by the absence (vs. presence) of composite oral papillae, fewer and smaller oral papillae in general, and a low (vs. high) diversity in dermal denticle morphology. In B. vivaldii , the dermal denticles have several ridges and are leaf-like to teardrop-shaped on the snout and tricuspidate on the trunk and tail, whereas in B. bachi the denticles have few ridges and their morphology is highly diverse in different body areas (see Weigmann et al. 2016 for details). B. vivaldii further differs from B. bachi in having smaller eyes (length 6.8–6.9 vs. 3.6–5.4 times in head length and 13.9–15 vs. 9–13 times in predorsal distance), a head length 1.3–1.6 vs. 0.6–1.3 times pectoral - pelvic space, a first dorsal fin 1.7–1.8 vs. 0.8–1.2 times higher than the second one, a first dorsal-fin base 1.1–1.2 vs. 1.2–2.8 times fin height and 2.2–2.3 vs. 1–1.8 times in interdorsal space, a pectoral - pelvic space 1.1–1.3 vs. 1.5–3.2 times interdorsal space, a ventral precaudal length 73.5–73.9% TL vs. 61.7– 72.8% TL, a dorsal - caudal space 4.6–4.7% TL vs. 0–4.1% TL, a pelvic - anal space 10.7–13.1% TL vs. 3–8.6% TL, an anal - caudal space 1.6–2% TL vs. 0–1.6% TL, a pelvic - caudal space 23.1–24.5% TL vs. 13.9–22% TL, a spiracle length 0.8–0.9% TL vs. 0.3–0.6% TL, a lower labial furrow length 2.3–2.4% TL vs. 1–2.3% TL, a preouter nostril length 1–1.1 vs. 1.1–1.9 times internarial width, more strongly structured teeth, and higher diplospondylous precaudal (43–44 vs. 33–43) and total (135–140 vs. 124–132) vertebral counts.
The new species—in addition to the differences listed in the first paragraph of the Discussion—further differs from Bythaelurus naylori in the presence (vs. absence) of oral papillae, a dorsal precaudal length 75.1–78% TL vs. 72.5–75.9% TL, a prepelvic length 41.3–43.5% TL vs. 47–51.6% TL, a snout-anterior vent length 44.8–45.8% TL vs. 49.9–54.7% TL, a dorsal - caudal space 4.6–4.7% TL vs. 1–2% TL, a pectoral - pelvic space 15–17.1% TL vs. 23.5–31.3% TL, a pelvic - anal space 10.7–13.1% TL vs. 4.8–9.4% TL, an anal - caudal space 1.6–2% TL vs. 6– 8.6% TL, an eye length 3.3–3.5% TL vs. 3.9–4.7% TL, an eye height 1.5–2% TL vs. 0.8–1.3% TL, a mouth width 9.2–9.6% TL vs. 7.2–9.2% TL, a caudal peduncle height 2.9–3.2% TL vs. 3.4–3.7% TL, a caudal peduncle width 1.6–2% TL vs. 2–2.3% TL, a smaller first dorsal fin (total length 7.5–8.4% TL vs. 9.9–11.1% TL, anterior margin length 7.4–8.6% TL vs. 9.8–11.4% TL, base length 5.7–6.1% TL vs. 6.4–8.9% TL), a shorter pelvic-fin anterior margin (5.1–5.7% TL vs. 5.8–8.2% TL), a smaller anal fin (anterior margin length 6.2–6.6% TL vs. 7.3–9.3% TL, height 2.9–3.7% TL vs. 4–6.7% TL), a lower caudal fin (height 5.7–6.6% TL vs. 6.5–8.0% TL), a shorter caudal fin postventral margin (10.6–11.4% TL vs. 11.3–14.9% TL), a larger pre-outer nostril length (3–3.5% TL vs. 2.2– 2.9% TL), and higher diplospondylous precaudal (43–44 vs. 25–38), total precaudal (81–82 vs. 64–78), caudal (54–58 vs. 44–54) and total (135–140 vs. 114–128) vertebral counts.
Compared to its slender-bodied congeners in the western Indian Ocean, i.e. Bythaelurus clevai , B. hispidus , B. lutarius and B. tenuicephalus , the new species is—in addition to the stout body shape of large specimens and the differences listed in the first paragraph of the Discussion—further distinguished by an anterior head of intermediate width (width at level of lateral indention 7.1–7.2% TL vs. 7.6% in B. clevai vs. 6.2% in B. hispidus vs. 6.7% in B. lutarius vs. 6.0–6.1% in B. tenuicephalus , width at max. outer extent of nostrils 8.5–8.6% TL vs. 9.2% in B. clevai vs. 8.8% in B. hispidus vs. 8.1% in B. lutarius vs. 7.2–7.3% in B. tenuicephalus ), a broader posterior head (width at mouth corners 12.9–13.5% TL vs. 11% TL vs. 11.7% TL vs. 9.4% TL vs. 10.1–11% TL, width at middle gill slits 10.4–12.7% TL vs. 9.5% TL vs. 8.9% TL vs. 9% TL vs. 9.5–10% TL), longer spiracles (0.8–0.9% TL vs. 0.5% TL vs. 0.3% TL vs. 0.7% TL vs. 0.4–0.6% TL), a broader mouth (width 9.2–9.6% TL vs. 6.6% TL vs. 7.5% TL vs. 6.5% TL vs. 7.6%), longer lower labial furrows (length 2.3–2.4% TL vs. 1.9% TL vs. 1.7% TL vs. 1.9% TL vs. 1.8–2.3% TL), a larger precaudal length (dorsally 75.1–78% TL vs. 74.5% TL vs. 72.5% TL vs. 72.8% TL vs. 72.8–73.7% TL, ventrally 73.5–73.9% TL vs. 70.6% TL vs. 69% TL vs. 70% TL vs. 68.6–71.3% TL), higher pectoral fins (height 9.6–12.1% TL vs. 9.6% TL vs. 9.1% TL vs. 5.8% TL vs. 8.5–9.8% TL), larger pelvic fins (total length 10.7–11.5% TL vs. 8.6% TL vs. 10.4% TL vs. 9.9% TL vs. 9.2–9.3% TL, base length 7–8% TL vs. 4.5% TL vs. 6.9% TL vs. 6.2% TL vs. 4.4–5.8% TL, height 4.9–5.3% TL vs. 3% TL vs. 2.6% TL vs. 1.9% TL vs.
2.8–3.4% TL, posterior margin 7.5–7.6% TL vs. 4.6% TL vs. 5% TL vs. 4.6% TL vs. 5.4–5.6% TL), a higher caudal-fin terminal lobe (height 5% TL vs. 2.7% TL vs. 2.7% TL vs. 4% TL vs. 2.7–2.8% TL), a higher trunk at pectoral-fin base ends (height 8.5–10.2% TL vs. 7.9% TL vs. 5.8% TL vs. 6.4% TL vs. 7.3–8.3% TL), and a higher tail at pelvic-fin base ends (5.5–5.9% TL vs. 4.8% TL vs. 4.3% TL vs. 4.5% TL vs. 4.5–4.9% TL). Additionally, the new species has a lower spiral valve turn count than the slender-bodied congeners in the western Indian Ocean with counts known (7 vs. 8–10 in B. hispidus vs. 10 in B. lutarius ).
The stout body shape of the holotype as compared to the more slender paratype and the aforementioned slender-bodied congeners in the western Indian Ocean is expressed in a larger trunk width at pectoral-fin base ends (11% TL) than in the paratype (9.4% TL) and the slender-bodied congeners (9.4% TL in B. clevai vs. 10% TL in B. hispidus vs. 8.5% TL in B. lutarius vs. 8–10.3% TL in B. tenuicephalus ). Additionally, most head width measurements are larger in both type specimens of the new species as compared to their slender-bodied congeners in the western Indian Ocean.
Bythaelurus vivaldii is further distinguished from each of those congeners in several morphological, morphometric and meristic differences, which are not compared in detail. Obvious differences, e.g., include the virtual absence of oral papillae in B. lutarius (vs. present in B. vivaldii ) and a snout without distinct lateral indentions anterior to the outer nostrils in dorsoventral view in B. tenuicephalus (vs. snout with strong lateral indentions).
A detailed comparison of Bythaelurus vivaldii with B. alcockii is not possible due to the lack of specimens or detailed descriptions. Following Alcock’s (1899) description of the lost holotype of B. alcockii , B. vivaldii has a strongly different coloration (blackish with hoary gray surface and some fins white-tipped posteriorly in B. alcockii ) and tooth morphology (anterolateral teeth in upper jaw with median cusp much longer than lateral cusps vs. teeth with cusps and lateral cusps of subequal length). Furthermore, the new species was caught in much shallower water (628 m vs. 1134–1262 m depth). The dark coloration and long snout combined with great catch depth, as reported for B. alcockii by Alcock (1899), indicate that this species possibly belongs to Apristurus rather than to Bythaelurus , like assumed by Compagno (1984a). However, the tooth morphology described for B. alcockii by Alcock (1899) does not fit the characters of neither Bythaelurus nor Apristurus species and therefore seems questionable. For the time being, B. alcockii remains a species of uncertain validity and generic assignment.
As Bythaelurus bachi View in CoL and B. naylori View in CoL fell as sister to B. dawsoni ( Springer, 1971) View in CoL from off New Zealand in molecular analyses published by Weigmann et al. (2016), the new species is also compared to the latter, allopatric congener, which supposedly differs in coloration, as well as numerous morphometric and meristic characters. Compared to B. dawsoni View in CoL , B. vivaldii View in CoL has a larger precaudal length (dorsally 75.1–78% TL vs. 63.7–67.9% TL, ventrally 73.5–73.9% TL vs. 59.4–63.9% TL), a larger pre-second dorsal fin length (67–67.9% TL vs. 54.5–57.1% TL), a larger pre-first dorsal fin length (48.2–49.6% TL vs. 40.8–43.3% TL), a longer head (23–24.1% TL vs. 18.9–19.7% TL), a larger prepectoral length (20.1–22.9% TL vs. 16.5–17.7% TL), a larger pre-anal fin length (60.2–62.5% TL vs. 47.7–50.5% TL), a larger interdorsal space (12.8–13.3% TL vs. 8.6–8.8% TL), a larger pelvic - caudal space (23.1–24.5% TL vs. 16.5–19.1% TL), a larger intergill length (5.6–6% TL vs. 3.9–4% TL), a smaller second dorsal fin (total length 6.9–7.3% TL vs. 10–11.1% TL, anterior margin length 5.1–7% TL vs. 10–11% TL, base length 5.3% TL vs. 7.3–7.7% TL, vertical height 2.8–3.1% TL vs. 3.8–3.9% TL), an anal-fin base length almost twice (vs. less than 1.5 times) second dorsal-fin base length, a much shorter caudal fin (caudal fin length 23.8–24.4% TL vs. 34.4–39.1% TL), a broader mouth (width 9.2–9.6% TL vs. 7.3–7.8% TL), a larger internarial width (3.2% TL vs. 2–2.1% TL), a broader posterior head (width at mouth corners 12.9–13.5% TL vs. 11.2–11.6% TL, width at middle gill slits 10.4–12.7% TL vs. 9.3–9.6% TL), and higher diplospondylous precaudal (43–44 vs. 31–36), total precaudal (81–82 vs. 67–73), and total (135–140 vs. 124–129) vertebral counts.
In comparison with its further congeners outside the western Indian Ocean, B. vivaldii View in CoL n. sp. supposedly differs in coloration and numerous morphometric differences, which are not compared in detail due to the large geographic distance and apparent morphological dissimilarity. The most obvious differences of the new species as compared to B. canescens ( Günther, 1878) View in CoL include an anal-fin base length almost twice (vs. less than 1.5 times) second dorsal-fin base length and much smaller maturity and maximum sizes (maturity size 30 cm TL, maximum size 35 cm TL vs. maturity size 52–59 cm TL, maximum size 73 cm TL), compared to B. giddingsi McCosker, Long & Baldwin, 2012 View in CoL the presence (vs. absence) of posterior nasal flaps, much higher tooth row counts per jaw (64–75 vs. 20–26), and a smaller maturity size (30 cm TL vs.> 45 cm TL), compared to B. immaculatus ( Chu & Meng, 1982) View in CoL the presence of distinct labial furrows, with lowers noticeably longer than uppers (vs. labial furrows reduced), a preorbital snout length almost twice eye length (vs. subequal to eye length), and a much smaller maximum size (35 cm TL vs. 76 cm TL), and compared to B. incanus Last & Stevens, 2008 View in CoL the presence of distinct labial furrows, with lowers noticeably longer than uppers (vs. labial furrows reduced), a pre-vent length smaller than tail length (vs. pre-vent length exceeding tail length), and a smaller maturity size (30 cm TL vs.> 45 cm TL).







| ZMB |
Museum f�r Naturkunde Berlin (Zoological Collections) |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Bythaelurus vivaldii
| Weigmann, Simon & Kaschner, Carina Julia 2017 |
Bythaelurus
| Kaschner 2015: 121 |
Halaelurus lutarius: Springer & D’Aubrey, 1972: 6
| Springer 1979: 82 |
| Bass 1975: 11 |
Scylliorhinus hispidus:
| Brauer 1908: 7 |