
Branchinecta lutulenta, Rogers, D. Christopher & Hill, Matthew A., 2013
|
publication ID |
https://doi.org/10.11646/zootaxa.3694.3.5 |
|
publication LSID |
lsid:zoobank.org:pub:20516F66-54A7-436F-8092-D9E5C0ED4EC9 |
|
DOI |
https://doi.org/10.5281/zenodo.6156778 |
|
persistent identifier |
https://treatment.plazi.org/id/FB7FA973-5908-FFA1-56C1-F92C3DE4B50E |
|
treatment provided by |
Plazi |
|
scientific name |
Branchinecta lutulenta |
| status |
sp. nov. |
Branchinecta lutulenta View in CoL sp. nov.
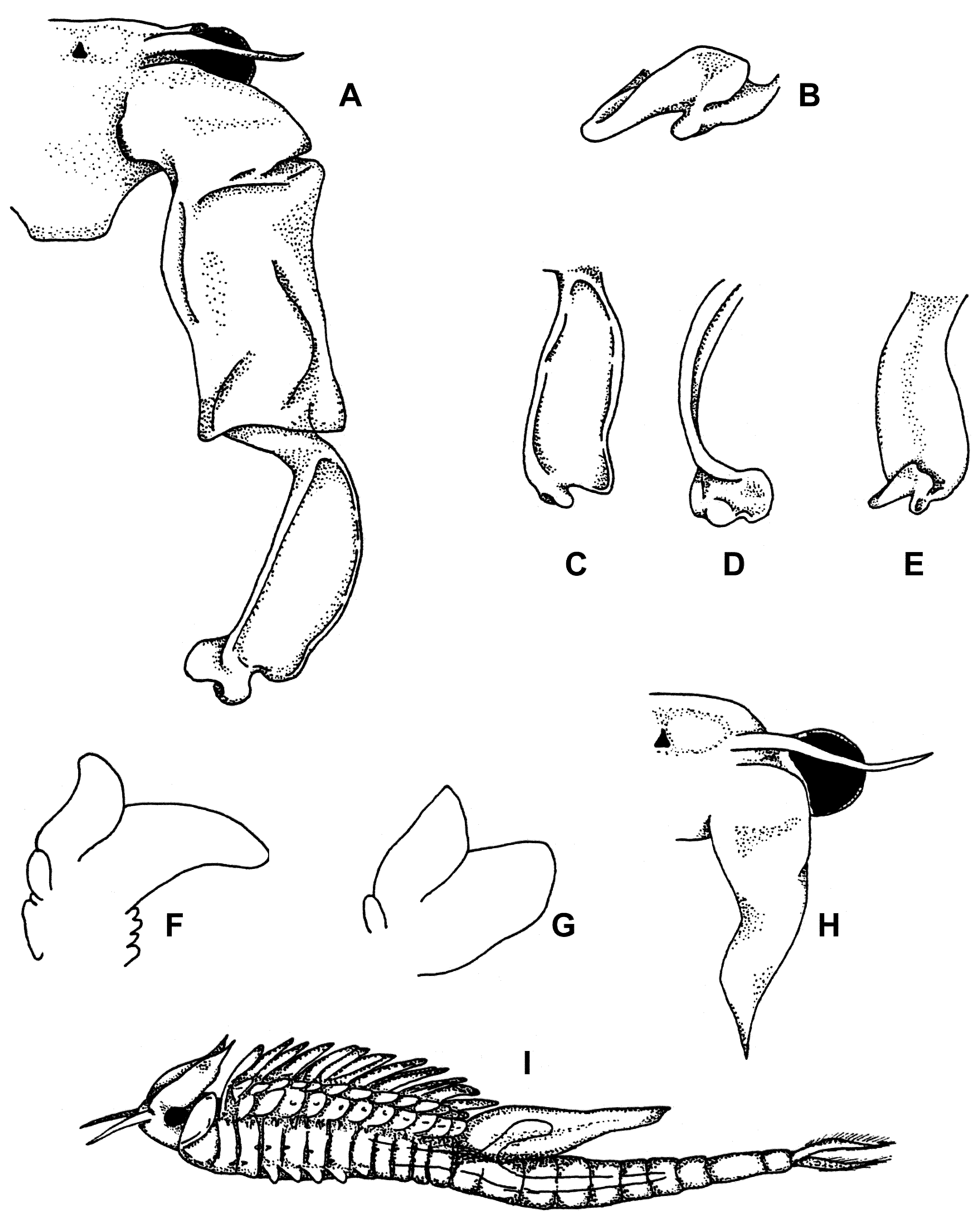
( Figure 1 View FIGURE 1 )
Etymology. The specific epithet comes from the Latin adjective; “ lutulentus, -a, -um ” meaning “muddy”. Lynch probably chose this name to reflect the thick, sticky, saline mud where the habitat of this fairy shrimp occurs. The common name that he gave this animal was the “muddy fairy shrimp”. Lynch spelled the name “lutalenta” on his handwritten labels in the specimen vials, which we assume is a misspelling, since his copious slide mounted material was labeled “ lutulenta ”.
Type locality. Both pools occur in poorly drained flood plain alluvium. The surrounding soil substrate has 15% carbonate content and the salinity (as electrical conductivity) is 8.0 mmhos/cm. The pools are alkaline. Water quality data was collected 28 March 2011. The upper and lower pools had conductivities of 3.2 mmhos/cm and 2.2 mmhos/cm and pH values of 8.89 and 8.58, respectively. The upper pool was approximately 16m in greatest diameter and 10cm deep. The lower pool was approximately 30m in greatest diameter and 30cm deep. The upper pool flowed into the lower pool via a 1 meter wide by 2cm deep vegetated channel. B. lutulenta sp. nov. was the only anostracan species present in the upper pool and co-occurring with Branchinecta mackini in the lower pool, the latter being much more common. Water samples collected from the site were analyzed in the lab and found to have a salinity of 0.10%.
Type material. Holotype. Male, data: USA: WASHINGTON: Grant Co.: 46˚ 49’ 55.65” N, 119˚ 47’ 31.14”W; pool approximately 150m north of Lower Crab Creek Road, approximately 15m west of fishing access road, and approximately 18m south of Crab Creek, south of Jericho; pool composed of an upper small pool, which drained into a larger lower pool, USNM 1207958.
Paratypes. One male, one female, same data: USNM 1207959.
Material examined. USA: WASHINGTON: Grant Co.: (From the original label) “pond 1.2 mile west of Smyrna”, USNM-00256561, 50 males, 50 females; (From the original label) “ 12.1 mile E of Beverley and 1.9 mile W of Smyrina (sic), pond” USNM-00170212, 50 males, 50 females; Pools at 46˚ 49’ 55.65” N, 119˚ 47’ 31.14”W, 28 March 2011, 21 males, 11 females, M. A. Hill;
Pools at 46˚ 49’ 55.65” N, 119˚ 47’ 31.14” W, 29 August 2012, 14 males, 9 females, reared from soil collected by M. A. Hill. In addition we examined 26 slide mounted specimens prepared by Lynch. These slides presented the second antennae, the thoracopods of both males and females, the brood pouch and the cercopods. Unfortunately, due to the age of the slides and how they were stored after Lynch’s death, the value of the slides was negligible.
Description. Male. ( Figure 1 View FIGURE 1 A–F) Average length (from anterior margin o head to posterior margin of telson, not including cercopods) of preserved material 22 mm.
Head rounded, anteriolateral corners not projecting over compound eye peduncles.
Labrum smooth, rounded, and directed posteriorly.
Mandibles and maxillae typical for the non-predatory members of the genus.
First antenna 1.25 times as long as stalked compound eye, with apex truncated, bearing four to six setae.
Second antennae extending posteriorly to fifth thoracic segment. Second antennal proximal antennomere subcylindrical, approximately four times as long as broad. Medial surface generally with small, scattered spinules. Apophysis absent. Anteriomedial pulvillus present. Second antennal distal antennomere subequal in length to proximal antennomere, laterally flattened, arcing medially, curving evenly along its length, broadening distally. Distal antennomere bent anteriorly in proximal third. Posterior margin expanded slightly in distal third, truncating towards apex. Apex broadly rounded, and twisted so that the medial surface is directed anteriorly, and the lateral surface directed posteriorly. Medial margin of apex folded back, medially, giving apex a trilobed appearance. Rasp absent.
Thoracopods serially homologous, increasing in size from the anterior pair to the fifth pair, then decreasing in size posteriorly to the eleventh pair. Praeepipodites and epipodites typical for genus. Thoracopod I with exopodite broadly oval and the endopodite triangular. Thoracopod V with exopodite oval, but arcing laterally, with a rounded apex; endopodite broad, elongate, arcing medially. Thoracopod XI with exopodite oval. Endopodite subtriangular.
Genital segments smooth and typical for the genus. Gonopods each with basal portion bearing a large ventral subconical, ventroposteriorly directed lobe, and a ventromedial digitiform spur. Everted gonopods extending to the second abdominal segment. Apices of everted gonopods bearing two lateral, spine covered mounds, each bearing ten to fifteen recurved spinules about one fourth as long as mound. Testes extending from genital segments (thoracic segments XII and XIII) posteriorly to abdominal segment IV.
Cercopods as typical for the non-predatory members of the genus.
Female. ( Figure 1 View FIGURE 1 G, H, and I) Average length of preserved material (from anterior margin o head to posterior margin of telson, not including cercopods) 17 mm.
Head without dorsal protuberances. Anterior surface of head and labrum with fine spinules. Labrum, mandibles and maxillae as in male.
First antennae subequal in length to second antennae.
Second antennae smooth, without any protrusions. Apex acute, tapering to a short point.. Thorax with rounded, conical dorsolateral projections on segments III–VII. Projections on thoracic segments
III–VI similar in size and shape. Projection on thoracic segment VII one third the size of preceding segments. Thoracopods generally similar to male, save that the endopodites and exopodites are more ovate and not
elongated.
Genital segments and abdominal segments smooth. Brood pouch fusiform and smooth. Brood pouch extending to abdominal segment five or six. Ovaries biramal, extending anteriorly into thoracic segment VI, and posteriorly into abdominal segment IV or V.
Cercopods as typical for the genus.
Egg. Approximately 300 µm in diameter, with dense, small, shallow, rounded depressions 0.1–0.25 times the diameter of the egg.
Differential diagnosis. Branchinecta lutulenta sp. nov. is separated from all other Branchinecta species by the form of the male second antenna apex, which is rotated so that the medial surface is facing anteriorly, and by the medial fold in the apex, giving a trilobed appearance. Some South American species have a truly trilobed apex on the second antenna. However, in those species the apex is neither rotated nor folded. Males are most similar to B. lindahli Packard, 1883 , B. sandiegonensis Fugate, 1993 and B. oterosanvicentei Obregón-Barboza et al., 2002 in that they share a very similar second antennal proximal antennomere, and the broad, laterally flattened distal antennomere. However, the apex in these species is not rotated and folded.
Females are most similar to B. lindahli and B. oterosanvicentei , but can be separated by the dorsolateral projections (most clearly obvious in mature specimens) being limited to thoracic segments III–VII, as opposed to IV–XI in B. lindahli and B. oterosanvinceti .
Comments. The anostracan family Branchinectidae and the genus Branchinecta were recently redefined (Rogers and Coronel 2011). We describe here a new Branchinecta species, originally discovered in 1959 from two wetlands, but not seen again. We rediscovered this species after extensive field surveys in the vicinity of the original collecting sites.
James E. Lynch ( 1892–1975), Professor Emeritus of the College of Fisheries, University of Washington was an important figure in Branchinecta and Lepidurus taxonomy. He was an expert in Branchiopod crustaceans, as well as echinophilous ciliates (see Lynn and Berger 1973) and acanthocephalans (Lynch 1936, Van Cleave and Lynch 1950). Lynch discovered and described four species of Branchinecta from the Pacific Northwest of the USA (1937, 1958, 1960, 1972a), and after untangling the confusion created by Packard (1874, 1883) and Pearse (1912, 1918), redescribed three species according to modern standards (Lynch, 1958, 1964). Lynch’s work provided the basis for taxonomic evaluation of the Branchinecta species (Lynch, 1972a).
Lynch used to travel from his home in western Washington State to visit his sister in Spokane. On these trips, he would take various routes across the desert portions of the Columbia Plateau and collect branchiopod crustaceans (Lynch’s notes, USNM). In 1959, he collected an undescribed species of Branchinecta from two pools between Beverley and Smyrna. The material was deposited at the USNM after his death. Lynch had designated the material with a scientific and a common name, and we retain both. In both of Lynch’s collections as well as our own, B. lutulenta sp. nov. was found cöoccurring with B. mackini Dexter, 1956 . Both species are of similar age and density in all collections.
One of us (DCR) made efforts to find Lynch’s original B. lutulenta sp. nov. localities, or additional populations in the Beverly/ Smyrna/ Royal City vicinity in 1997, 1998, 1999, 2000, 2002, 2007 and 2008. One of us (MH) conducted separate surveys in 2007, 2008 and 2009 and successfully found our population in 2011. It was noted that Lynch’s site 1.2 miles West of Smyrna was likely destroyed by the digging of a quarry and associated drainage structures. The site 1.9 miles west of Smyrna possesses numerous clear, grassy pools devoid of anostracans along the railroad bed. A small pool here formed by a wheel rut in a dirt road contained Branchinecta mackini when sampled on 28 March 2011. The fate of Lynch’s original sites are unknown.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |