
Bothriolepis leptocheira jeremejevi ( Rohon, 1900 ) Lukševičs & Beznosov & Stūris, 2017
|
publication ID |
https://doi.org/ 10.4202/app.00265.2016 |
|
DOI |
https://doi.org/10.5281/zenodo.11061574 |
|
persistent identifier |
https://treatment.plazi.org/id/615E8A3A-4C53-FFF0-FCAC-84C7A1C7F4F6 |
|
treatment provided by |
Felipe |
|
scientific name |
Bothriolepis leptocheira jeremejevi ( Rohon, 1900 ) |
| status |
comb. nov. |
Bothriolepis leptocheira jeremejevi ( Rohon, 1900) comb. nov.
Figs. 3–14 View Fig View Fig View Fig View Fig View Fig View Fig View → View Fig View Fig View Fig View Fig View Fig .
1900 Bothriolepis jeremejevi sp. nov.; Rohon 1900: 30, fig. 18, 19.
1932 Bothriolepis jeremejewi Rohon, 1900 ; Gross 1932: 34.
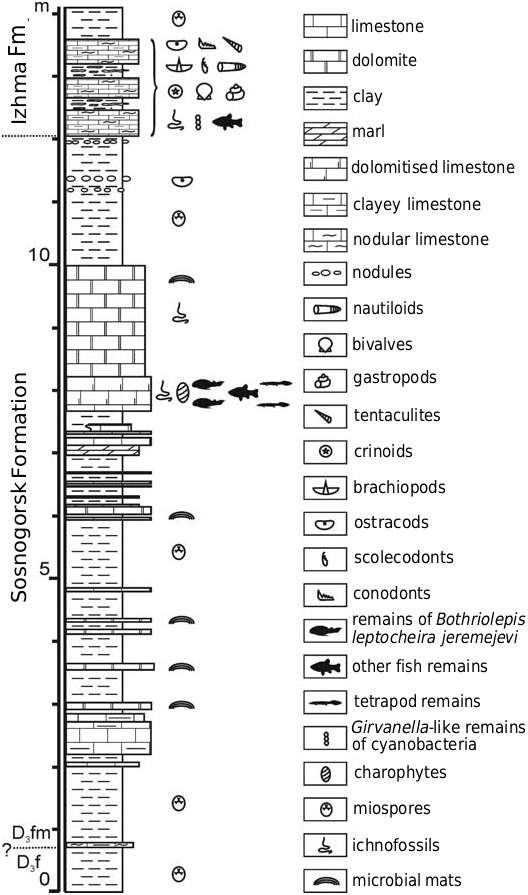
Lectotype: PIN 1350/12 fragmentary Cv 1, illustrated by Rohon (1900: fig. 19).
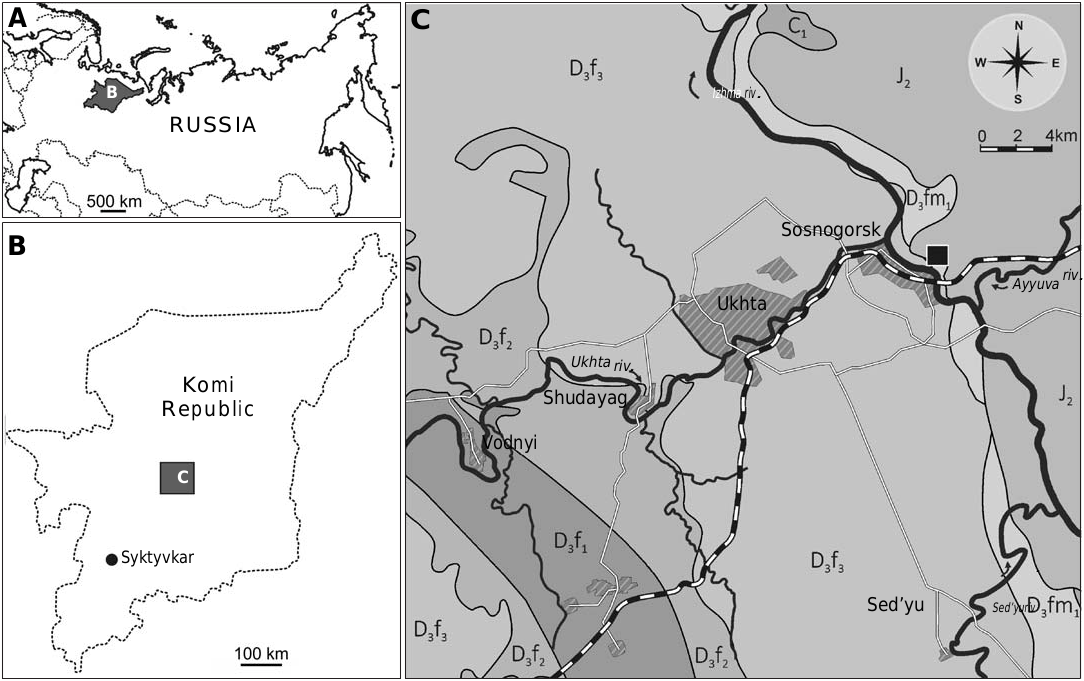
Type locality: Outcrop on the banks of the river Izhma, opposite Sosnogorsk, South Timan, Komi Republic, Russia.
Type horizon: Sosnogorsk Formation, Volgograd Regional Stage, lower Famennian, Upper Devonian .
Nomenclatorial remarks: Gross (1932: 26) selected the AMD plate 473 from the Cil’ma River (North Timan) as a of Bothriolepis jeremejewi Rohon, 1900 (erroneous spelling by Gross) illustrated by Rohon (1900: fig. 18). This specimen was most probably found in the lower Frasnian deposits and belongs to Asterolepis radiata ( Rohon, 1900) ; it has never been found neither in CNIGR nor in PIN collections since the beginning of 1920-ies. However, the second specimen illustrated by Rohon (1900: fig. 19), 318b from the river Izhma (South Timan) has been identified by Alexander Ivanov in 1994 in the collection of PIN. Rohon (1900) erroneously identified it as the distal segment of the pectoral fin; in fact this is fragmentary Cv 1 PIN 1350/12 from the outcrop on the banks of the river Izhma, judging from the original label. This specimen is about 40 mm long light brown plate embedded in the light yellow limestone; colour and preservation of the bone, as well as character of the rock are typical for the “fish dolomite” from the locality on the right bank of the river Izhma opposite Sosnogorsk, South Timan, Russia. Therefore the PIN 1350/12 illustrated by Rohon (1900: fig. 19) is designated here as the lectotype of Bothriolepis leptocheira jeremejevi ( Rohon, 1900) .
Material.— PIN 208, part of the head shield, two fragmentary Cv 1, Nu, AMD, proximal segment of the pectoral appendage (on a single slab of limestone); IG KSC 155 (more than 160 disarticulated but mainly complete bones prepared out of the rock, some semiarticulated head shields, and three specimens demonstrating almost complete ventral wall of the trunk armour on several slabs), and IG KSC 71/И (many blocks with disarticulated fish remains, usually each block containing several specimens; only small part of all specimens is prepared); CNIGR 1226/1073, PVL; 1226/1074, PMD; 1226/1078, AMD (for complete list of specimens from IG KSC see the Appendix 1). All material from the type locality and horizon .
Description.— Bothriolepis leptocheira is well-represented in limestone from the Sosnogorsk locality by some articulated head shields and ventral walls of the trunk armour, some articulated proximal segments of the pectoral fin, as well as by numerous disarticulated plates of the head and trunk shields, and pectoral appendage. Most are from individuals of moderate size; however, there are remains from a very large, as well as from very small individuals. The plates are usually slightly flattened, sometimes deformed and bear cracks, most probably due to compaction of the fossil-bearing limestone.
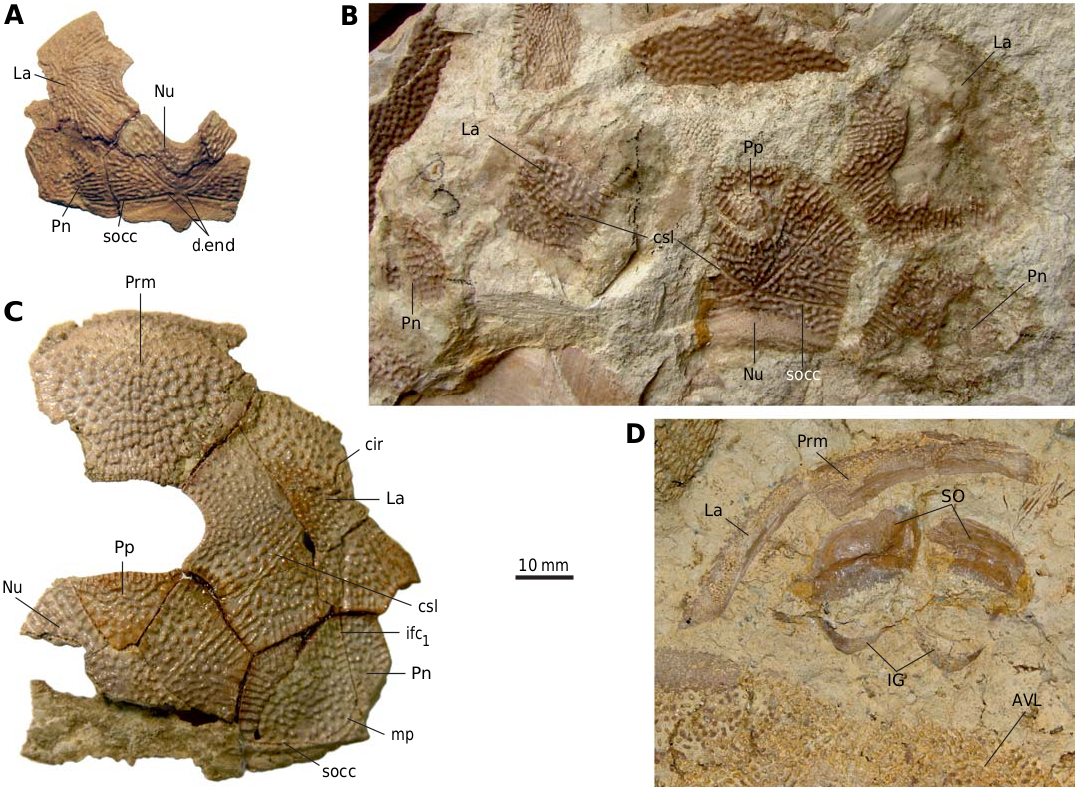
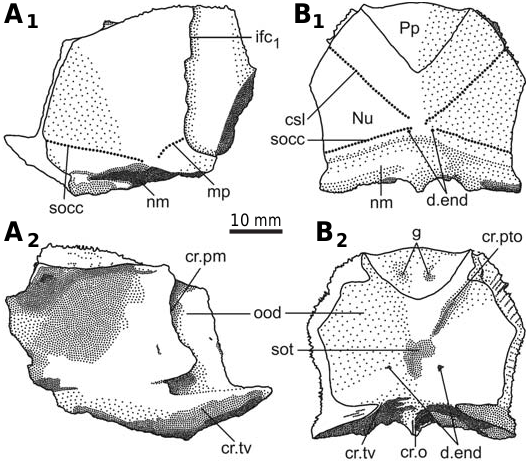
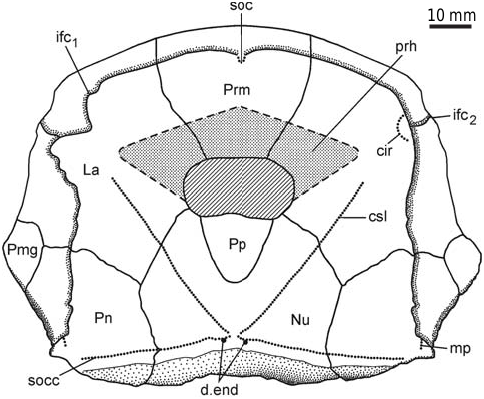
The head shield ( Fig. 3 View Fig ) is moderately broad, with an average B/L index 1.24 (n = 2), slightly narrower than the head shield of Bothriolepis leptocheira curonica Gross from Latvia, but almost similar to that of B. ornata ( Lukševičs 2001) . The rostral margin is moderately convex and only slightly longer than the posterior margin. The head shield is weakly vaulted both rostrocaudally and transversely, as in Scottish material ( Miles 1968), but the anterior part of the Prm and La plates is strongly curved. The antero-lateral corner, the prelateral notch (nprl; Figs. 4C, D View Fig 2 View Fig , 5 View Fig ) and the lateral process are usually weakly defined, as in B. leptocheira from Scotland and Latvia. The orbital fenestra is well seen in IG KSC 155/162 ( Fig. 3C View Fig ) and IG KSC 155/45 representing individual of moderate size; it is relatively small as in other subspecies of B. leptocheira , and with L/B index of 0.53. The preorbital recess (prh) is of trifid type, with the lateral horns less extended laterally than these in B. maxima or B. gigantea , but more broad at their base ( Figs. 4 View Fig , 5 View Fig ). The borders of the otico-occipital depression (ood; Figs. 6 View Fig , 7 View Fig ) on the internal surface of the head shield are well defined by the paramarginal crista (cr.pm) on the paranuchal plates Figs. 6 View Fig , 7 View Fig ), which are rather low on the lateral plates ( Figs. 4 View Fig , 5 View Fig ). The antero-lateral corners of the otico-occipital depression (the imprints of the anterior postorbital processes of the endocranium, pr.po; Fig. 5F View Fig 2 View Fig ) are moderately narrow at the base in comparison with these in B. ornata . In most cases the anterior postorbital processes do not extend in front of the anterior margin of the orbital fenestra, contrary to these in Scottish material ( Miles 1968: 76).
The premedian plate (Prm; Figs. 3C View Fig , 4H View Fig , 5H View Fig ) is slightly wider than long, with a B/L index of 1.07–1.37. It is slightly narrower than the Prm of B. leptocheira from Latvia (1.23– 1.48; Lukševičs 2001), but approximately coincides with proportions of Prm in Scottish material of B. leptocheira Miles [1968: 76] has mentioned B/L index of 1.14, 1.20, and 1.28). Prm is weakly arched in a lateral direction, but rather strongly vaulted in the parasagittal section with curvature in the anterior most part. The rostral margin is convex, it is 2.5–3.2 times longer than the slightly concave or straight orbital margin, similar to B. leptocheira from Latvia and Scotland (for comparison see Lukševičs 2001: figs. 32–34 and Miles 1968: pl. 20: 1, 2), and bears slightly defined median rostral projection ( IG KSC 155/114; Fig. 4H View Fig 2 View Fig ) seen only in anterior view. Also the position of the infraorbital sensory groove (ifc 1) that crosses the plate close to its rostral margin and the well-defined anterior section of the supraorbital sensory line (soc) are similar characteristics in all three subspecies of B. leptocheira .
The lateral plate (La; Figs. 3 View Fig , 4A–G View Fig , 5A–G, I View Fig ) is short and broad with an overall shape and proportions resembling these of B. leptocheira curonica . The infraorbital sensory groove crosses the plate in its anterior part, not far from the anterior margin and along its lateral margins. The path of the sensory groove significantly varies in various specimens, e.g., on La plate IG KSC 155/15 ( Figs. 4A View Fig , 5A View Fig ) it displays a strong deviation from its usual path ( Fig. 4B–G View Fig ) because it partly follows the course of the semicircular pit-line groove, which is missing in this specimen ( IG KSC 155/15). The plate is rather strongly vaulted in its anterior most part in large individuals ( IG KSC 155/66; Fig. 4E View Fig 2 View Fig ), but almost flat in small individuals ( IG KSC 155/69; Fig. 4F View Fig ). A moderately small, but deep pit (p) is positioned just posterior to the spiracular groove (spg; Fig. 5F View Fig 2 View Fig ). The spiracular groove is moderately long and shallow. It is widest at the lateral margin of the head shield and narrows only slightly close to the pit. The anterior and posterior crests delimiting the spiracular groove are well developed along their entire extent. The anterior attachment for the submarginal plate (a 1 Sm; Fig. 5F View Fig 2 View Fig ) is strongly developed transverse structure, which borders the anterior crest of the spiracular groove. This attachment is very close to the more anteriorly located triangular attachment area for the prelateral plate (a.PrL; Fig. 5F View Fig 2 View Fig ). The posterior attachment for the submarginal plate (a 2 Sm) constitutes an elongate boss ( Fig. 5F View Fig 2 View Fig ), widening posteriorly. The attachment reaches anteriorly to the posterior crest delimiting the spiracular groove.
The postpineal plate (Pp; Figs. 6–8 View Fig View Fig View Fig ) is, as usually observed in Bothriolepis , slightly wider than it is long and has an L/B index varying from 0.96 to 0.97, it is slightly less elongated than in the material of B. leptocheira from Scotland ( Miles 1968: pl. 20: 4) and Latvia ( Lukševičs 2001: fig. 34). The rounded anterior margin of Pp protrudes more into the orbital fenestra of small specimens, e.g., IG KSC 155/155 ( Fig. 6C View Fig ) than in larger specimens ( Figs. 6A, B View Fig ). IG KSC 155/146 is an asymmetrical plate with wider right part of the bone and pointed, rather than rounded posterior angle ( Fig. 6B View Fig ). Two large, rounded pits (g; Fig. 7B View Fig 2 View Fig ) on the internal surface of the postpineal are separated by a low median ridge.
The nuchal plate (Nu; Figs. 3 View Fig , 6–8 View Fig View Fig View Fig ) has an L/B index of about 0.83 ( Miles 1968 mentioned an L/B index of 0.83 for Scottish material and Lukševičs 2001 mentioned an L/B index of 0.84 for material from Latvia). The plate is always broadest across the lateral corners. The anterior division of the lateral margin is usually convex and a little shorter than the concave posterior division ( Figs. 6D–H, K View Fig , 7B View Fig ). The orbital facets are very short, tapering to a point in IG KSC 155/53 and IG KSC 155/154 ( Fig. 6F, G View Fig ). The posterior margin is usually strongly concave; however, IG KSC 155/54-1 and IG KSC 155/56 demonstrate convex posterior margin ( Fig. 6H, I View Fig ). The median process is more pronounced in specimens with a concave posterior margin. The external openings for the endolymphatic ducts (d.end) are closely set to one another, in a distance of 2.9 mm in IG KSC 155/53. The supraoccipital cross-commissural pit-line groove (socc) is usually well defined, however, in IG KSC 155/53 ( Fig. 6F View Fig ) it is seen only as a line of pores along the posterior margin. Internally, the openings for the endolymphatic ducts are larger than their external openings, divided by the large supraotic thickening (sot; Fig. 7B View Fig 2 View Fig ). The transverse nuchal crista (cr.tv) is high and well developed. Posteriorly, the median occipital crest of the head shield (cro; Figs. 6 View Fig , 7B View Fig 2 View Fig ) is usually well developed.
The paranuchal plate (Pn; Figs. 3 View Fig , 6 View Fig , 7 View Fig ) is of moderate breadth, with an L/B index of about 0.92, whereas in B. leptocheira curonica it is broader, the index reaching 0.86 ( Lukševičs 2001); Miles (1968) mentioned that Pn of B. l. leptocheira is poorly known and in the restoration ( Miles 1968: text-fig. 36) it has an L/B index of about 0.80. The lateral division of Pn composes 39–44% (41.9 on the average) of the general breadth of the plate, thus being slightly broader than in B. leptocheira curonica with 38.7% on the average ( Lukševičs 2001). A long supraoccipital cross-commissural pit-line groove (socc) is present in all examined specimens; it terminates approximately at the level of the middle of the plate posterior margin. B. hydrophila (Agassiz, 1844) from the Upper Old Red Sandstone of Dura Den, Scotland, also possess a long supraoccipital groove; however, it never extends to the Pn plate. The middle pit-line groove (mp) is always present, as it is in B. leptocheira leptocheira and B. leptocheira curonica ( Lukševičs 2001) ; sometimes it is rather long, but it is not fused with the supraoccipital pit-line groove, contrary to B. leptocheira curonica where it is fused in some specimens ( Lukševičs 2001: fig. 33A, B). Internally, the paramarginal crest (cr.pm) is very distinct; it is of semilunar shape in IG KSC 155/117 ( Fig. 7A View Fig ). The postero-lateral angle of the otico-occipital depression is complicated by two postero-lateral projections. Paranuchal trochlea does not reach the lateral margin of the plate.
The postmarginal plate IG KSC 155/161 ( Fig. 4J View Fig ) is shorter and broader than it is in B. leptocheira curonica ( Lukševičs 2001: figs. 32A, B, F, 33A, B, F), more closely resembling B. leptocheira leptocheira ( Miles 1968: text-fig. 36) by its irregular quadrangular shape with an almost straight postero-mesial margin and strongly convex antero-mesial margin.
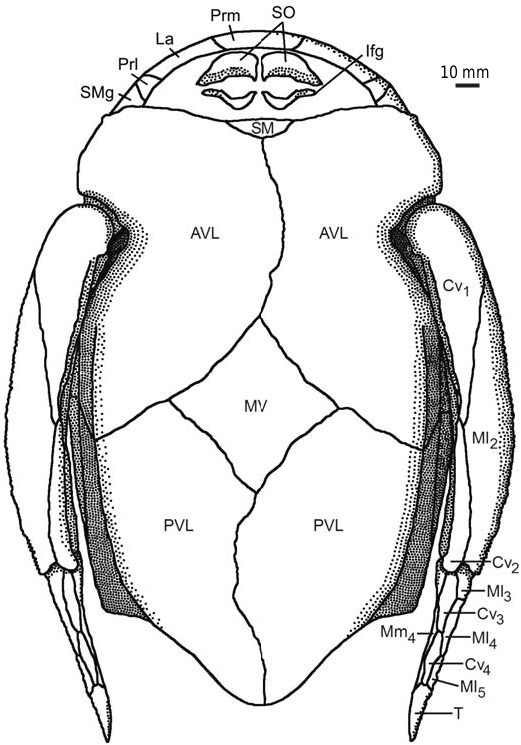
Thesuborbitalplates( SO)havenotbeenpreviouslyfound in this species. The right and left SO are preserved only from one individual: in IG KSC 155/10-3 these are seen in visceral (internal) view ( Fig. 3D View Fig ), and the IG KSC 155/10-2 ( Fig. 11A View Fig ) shows their cast. The shape of the plate is rhombic with a slightly convex anterior and an almost straight posterior margin. The lateral margin is slightly concave, forming a prominent postero-lateral corner. The mesial margin is oblique therefore the split between pair of suborbital plates widens posteriorly. The ventral margin seems to bear rather small tubercles showing weakly defined denticulation. The visceral surface is dominated by a pronounced transverse ridge, which begins near the dorso-mesial corner, and extends obliquely, towards the middle of the lateral edge of the plate. It forms a posteriorly oriented convex curve. The height of this ridge varies: large close to the mesial margin and becoming lower laterally. The morphology of the visceral surface of SO is almost similar to those described for other species of Bothriolepis : B. canadensis ( Stensiö 1948) , Bothriolepis sp. from Gogo ( Young 1984), and B. yeungae Johanson (1998) .
The S-shaped infragnathals of B. leptocheira are also described here for the first time; they are preserved in the IG KSC 155/10-2 and IG KSC 155/10-3 ( Figs. 3D View Fig , 11A View Fig , 14 View Fig ). The lateral projection of the right infragnathal lies very close to the tip of posterolateral corner of SO ( Fig. 3D View Fig ). Infragnathal is strongly curved in the mesial part and tapers mesially.
The trunk armour is narrow, relatively low, with a rather flattened dorsal wall, and the lateral wall more than three times as long as it is high. The length of the dorsal wall, estimated from the largest specimens of AMD and PMD plates, probably, slightly exceeds 200 mm. Thus it is larger than in B. leptocheira from Scotland ( Miles [1968] estimated the dorsal wall reached a length of some 140 mm), but smaller than in B. leptocheira curonica from Latvia estimated by Lukševičs (2001) as reaching 240 mm. The median dorsal ridge is weakly defined only in the posterior part of the PMD. The dorso-lateral and ventro-lateral ridges are well marked. The ventral wall of the trunk armour is not completely flat as it was demonstrated in the 3D model of B. canadensis ( Béchard et al. 2014) , but it is slightly convex both in transverse and longitudinal directions. The ventral wall is moderately wide with an estimated B/L index of 0.6 in IG KSC 155/10-3 ( Figs. 3D View Fig , 14 View Fig ), exceeding the estimated B/L index of about 0.53 in Latvian material of B. leptocheira ( Lukševičs 2001) .However, this difference could be explained by the use of separate AVL and PVL plates of moderate size to estimate the B/L index of the ventral wall of B. leptocheira curonica . The ventral wall is slightly narrower than that restored for B. prima and B. obrutschewi ( Karatajūte-Talimaa 1966: figs. 3, 10), but almost perfectly similar in the proportions to B. canadensis ( Béchard et al. 2014: fig. 3).
The anterior median dorsal plate ( AMD; Fig. 9 View → ) is weakly arched, relatively narrow, with a B/L index of about 0.78 in the Sosnogorsk material. The index is 0.83 in Latvian material ( Lukševičs 2001: 549), and about 0.77 in Scottish material ( Miles 1968: 78). The anterior margin is weakly convex, sometimes slightly concave in its middle ( IG KSC 155/44; Fig. 9G View → ), relatively narrow, and about 1.3 times longer than the posterior margin. The antero-lateral and lateral corners, and the postlevator processes (pr.pl) are well defined, especially in larger specimens. The posterior division of the lateral margin is 1.4–1.6 times shorter than the anterior division. There is no median dorsal ridge. Overlap areas for ADL and MxL are sometimes normally developed as typical for Bothriolepis ( IG KSC 155/44; Fig. 9G View → ), but usually demonstrate the so called Remigolepis - type overlapping: IG KSC 155/140 and IG KSC 155/42 ( Fig. 9E, F View → ) show the anterior ¼–⅓ of the posterior lateral margin overlapping MxL (cf. MxL) and remaining ¾ or ⅔ of that margin are overlapped by MxL. The same feature is also seen in IG KSC 155/108, where the AMD overlaps MxL to 1/6 of the common suture Fig. 9B View → ), and IG KSC 155/5, where the cf. MxL is even smaller ( Fig. 9A View → ). Unusual are sutural connections of AMD and ADL in IG KSC 155/44, where the posterior ⅓ of the anterior lateral margin of AMD is overlapped by ADL (oa. ADL; Fig. 9G View → ). Internally, the levator fossa (f.retr) is longer than it is wide; it is delineated by the low and narrow postlevator thickenings (alr). The low and short medial crest mc; Fig. 9A, B View → ) is more than twice shorter than the levator fossa. The supranuchal area (sna) is well defined and slightly broader medially from the antero-lateral corner. The centre of the anterior ventral pit (pt 1; Fig. 9A, B, D View → ) is located slightly anteriorly to the tergal angle (dma). The median ventral ridge (mvr) is relatively low; in IG KSC 155/113 ( Fig. 9D View → ) it is extremely short and divides into two crests which continue as a moderately deep median ventral groove (grm).
The posterior median dorsal plate ( PMD; Fig. 9H–N View → ) is broad in comparison with the AMD. Its B/L index varies from 0.84 to 1.00, 0.92 on the average in the Sosnogorsk material, whereas it reaches about 0.95 in the material from Scotland ( Miles 1968) and varies from 0.88 to 1.01, 0.95 on the average, in the material from Latvia ( Lukševičs 2001). The anterior margin is rather narrow, strongly convex and pointed in small individuals, and convex but rounded in large individuals. The posterior margin is well convex, 1.7–2.7 times longer than the anterior margin in the material from Sosnogorsk, but only 2–2.5 in the Scottish and Latvian material. The posterior corner of the PMD is not pronounced as in the Scottish ( Miles 1968: text-fig. 38) and Latvian material ( Lukševičs 2001: fig. 35C). The median dorsal ridge (dmr) is present only in the posterior third of the plate in medium-sized individuals, and only in the posterior quarter in the largest PMD IG KSC 155/142, or even absent in some large PMD; however, in the smallest individual IG KSC 155/148 reaching only 19 mm in length, it is seen along the whole plate. IG KSC 155/7 differs from all other specimens in possessing two distinct abnormalities which are most probably related to one another: (i) the posterior margin bears 5–6 mm wide unornamented area (pua) that superficially resembles the obtected nuchal area of the head shield; (ii) a branch of the sensory line canal (dlg 2) is clearly seen along the same posterior margin of the left side of the plate ( Fig. 9H View → ) in the position where this canal has been reported in B. canadensis ( Graham-Smith 1978: fig. 2e). The canal crosses the lateral margin of the PMD close to its posterior margin, runs parallel to the posterior margin and a few milimeters from the dorsal median ridge turns posteriorly and becomes deeper than in the most part of its course, “diving” into the bone. The 5–6 mm narrow belt between the canal and pua is ornamented by separate round and high tubercles, in some cases two or three of them fused into very short ridges, while the rest of the surface bears mainly vermiculating ridges and only several separate tubercles close to the canal. The ventral surface of the PMD shows a relatively low posterior transverse thoracic crest (cr.tp), in some specimens (e.g., IG KSC 155/1; Fig. 9J View → ) forming a distinct shelf on the short area posterior to the crest. There is a wide median ventral groove (grm) and a peculiar posterior ventral pit (pt 2; Fig. 9J, N View → ), which is funnel-shaped, quite elongated and oblique in sagittal view so that the anterior wall is very low and the posterior wall is rather high.
The dorsal lamina of ADL is of moderate breadth, about 2.6–3.3 times (2.9 on the average) as long as it is broad ( Fig. 10A–G View Fig ), thus the proportions of the ADL of B. leptocheira jeremejevi are more similar to those of B. leptocheira curonica (e.g., LDM 98/58; Lukševičs 2001: figs. 36B, 37A) and ADL is shorter than it is 3.5 times longer and broader in B. leptocheira leptocheira RSM 1859.33.632D ( Miles 1968: pl. 20: 3). The postnuchal ornamented corner (pnoa) is prominent in all studied specimens. The width of the dorsal lamina is usually slightly larger than the height of the lateral lamina. The dorso-lateral ridge (dlr) is well pronounced. Usually, the AMD overlaps the mesial margin of the ADL, but in IG KSC 155/116 ( Fig. 10A View Fig 2 View Fig ) the ADL overlaps the AMD (cf. AMD) along the posterior third part of the common suture. The outline of the mesial margin of this specimen greatly complements the lateral margin of AMD plate IG KSC 155/44 ( Fig. 9G View → ). Thus, this feature is not remarkably rare in B. leptocheira jeremejevi . The lateral lamina of ADL is 4.7 times as long as it is high in IG KSC IG 155/112 ( Fig. 10G View Fig ) and more than 6 times as long as it is high in IG KSC IG 71/i-68; the lateral lamina of ADL is slightly higher in the material from Latvia ( Lukševičs 2001: fig. 37B), and Scottish material does not show the lateral lamina well ( Miles 1968: 78). The processus obstans (pro) is rather deep, but the course of the main lateral line groove (lcg) conforms the usual pattern.
The dorsal lamina of MxL is slightly less than twice as long as it is broad ( Fig. 10H–J View Fig ). The dorsal corner is clearly seen. The lateral lamina is moderately low, about 2–2.5 times as long as it is high, but relatively higher than in material from Latvia where it is about three times as long as it is high ( Lukševičs 2001: fig. 37E). The posterior oblique sensory line groove (dlg 2) usually terminates in some distance from the lateral margin (e.g., IG KSC 155/82); as in B. leptocheira curonica LDM 98/53 ( Lukševičs 2001: fig. 36C), IG KSC 155/115 shows a dlg 2 crossing the dorso-lateral ridge ( Fig. 10I View Fig ). The separate dorso-ventral pit-line groove (dxp) is present in a medium sized IG KSC 155/144 ( Fig. 10H View Fig ), similarly to B. prima Gross, 1942 ( Karatajūte-Talimaa 1966: fig. 3), B. obrutschewi Gross, 1942 ( Karatajūte-Talimaa 1966: fig. 10), and B. evaldi Lyarskaya, 1986 ( Lyarskaya 1986). The dorso-lateral ridge (dlr) is well defined both in the ADL and MxL plates of small and medium sized specimens, as it is also seen in material from Latvia; Miles (1968: 78) claimed that “there is no clear development of the dlr on either the MxL or ADL ”; in fact dlr is not clearly seen, particularly in MxL, because of preservation of the Scottish material as internal impressions.
The semilunar plate ( SM) is not known, but from the configuration of the anterior margin of the AVL plates, the SM is of average proportions and normal shape ( Fig. 14 View Fig ).
The ventral lamina of the AVL is 1.6–2 times as long as it is broad ( Fig. 11 View Fig ). The anterior margin of the ventral lamina is rounded without clearly defined corners, similar to B. leptocheira leptocheira ( Miles 1968: 79, text-fig. 39) and B. leptocheira curonica ( Lukševičs 2001: figs. 37F, G, 38A). The subcephalic division comprises 21–30% (24% on average) of the total length of the ventral lamina (about 20% in the Scottish material [ Miles 1968]; about 21–25% with 22.5% on average in the Latvian material [ Lukševičs 2001: 551]). The lateral lamina is not preserved within the material from Scotland. The lateral lamina is low, and the ventral lamina is 3.8 times as broad as the lateral lamina is high in B. leptocheira curonica LDM 98/30. The lateral lamina is relatively higher in the Sosnogorsk material, e.g., IG KSC 155/122 ( Fig. 11D View Fig ) shows the ventral lamina is 2.9 times as broad as the lateral lamina is high. The right AVL overlaps the left AVL by a very narrow overlap area. The pectoral fin attachment area is usually not well preserved in the studied Sosnogorsk material, except on AVL IG KSC 155/122 ( Fig. 11D View Fig ), which shows the typical morphology of Bothriolepis . Internally, the crista transversalis interna anterior (cit 1; Fig. 11A View Fig ) is developed as a high, thin ridge running at an oblique angle across half the width of the anterior portion of the plate. The transverse thickening (cit 2) is more prominent in comparison with cit 1 as it is seen in the IG KSC 155/10-2 ( Fig. 11A View Fig ).
The posterior ventro-lateral plate ( PVL) has variable proportions ( Fig. 12 View Fig ). It is relatively narrow with a ventral lamina that is 2.1–2.7 times as longer than it is broad. The subanal division is rather narrow and relatively longer than in the Latvian material where it occupies about one third (28–38%) of the total PVL plate length. This is comparable with the 33% in B. leptocheira leptocheira ( Miles 1968: 80) , but slightly different from B. leptocheira curonica ( Luksevics 2001: figs. 38B, C, 39C) where it varies between 22% and 32%. The lateral lamina is moderately high in IG KSC 155/4 ( Fig. 12A View Fig ). The ventro-lateral ridge (vlr) is clearly defined both in the AVL and PVL. The left PVL overlaps the right PVL plate.
The median ventral plate ( MV; Fig. 11F–K View Fig ) is not known in the material from Scotland ( Miles 1968: 80) and Latvia Lukševičs 2001: 552), whereas it is well represented in the Sosnogorsk material by several disarticulated specimens and also found in partially articulated ventral walls of the trunk armour. This plate is slightly elongated with an L/W index of about 1.2 on average. The shape of AVL and PVL plates in B. leptocheira curonica ( Lukševičs 2001: 552) suggests a moderately small size of the MV; whereas in B. leptocheira jeremejevi , it is rather large ( Fig. 11A, F–K View Fig ). Almost all MV plates are from relatively large individuals, the plate reaching 30 to 61 mm in length. Some plates are asymmetrical in shape, e.g., IG KSC 155/77 ( Fig. 11H View Fig ) and 155/106 ( Fig. 11K View Fig ). Usually all margins of the plate are slightly convex or almost straight, but in IG KSC 155/133 both posterior lateral margins are concave ( Fig. 11J View Fig ). Overlap areas usually are rather narrow ( Fig. 11H, J View Fig ).
The pectoral fin is represented in the Sosnogorsk material by several disarticulated bones, at least three specimens showing articulated plates of the proximal segment associated with the AVL (e.g., Fig. 11B View Fig ), and one articulated distal segment ( Fig. 13E View Fig ). The proximal segment bears small rarely set lateral and mesial spines. The lateral spines are larger than the mesial ones (which are even not seen in figured specimens) as in B. leptocheira curonica ( Lukševičs 2001: figs. 38A, 39B); Miles (1968) suggested the mesial spines were perhaps absent in B. leptocheira leptocheira . The proximal segment is very long and narrow, it is 4.6 times longer than it is broad in IG KSC 155/10. It is slightly less elongated than in the Scottish material (5.5 to 6 times as long as it is broad; Miles 1968: 80) and in the Latvian material (4.8–6 [5.5 on the average] times as long as it is broad; Lukševičs 2001: 553). The Cd 1 has an L/B index varying from 2.6 to 2.9 (2.7–3.7, 3.3 on average in the Latvian material; Lukševičs 2001: 553); measurements were made during the field work in 2009. The Cv 1 is gently longer than the Cd 1 and slightly more elongated with an L/B index of 3.5–4.1 (3.9 on the average) ( Fig. 13A, B View Fig ). The Cd 2 is slightly, about 1.2 times, longer than it is broad in IG KSC 155/10. Similarly to that of B. leptocheira curonica , the distal segment ( Fig. 13E View Fig ) is adorned with small and rounded marginal spines; the lateral spines are large, sharp and proximally directed. The distal segment is very long and narrow, with an L/B index of 5.4 in both IG KSC 155/21 and IG KSC 155/22 (L/B index of 5.7 in LDM 89/4). The Cd5 plate is present in the material from all three countries.
The ornamentation is fine and basically of the reticular type ( Miles 1968: 80) in small individuals, changing into vermiculating type in the medium-sized fish, and even in the tubercular type on the largest specimens (e.g., Prm; IG 155/114), instead of becoming smooth in quite large individuals from Latvia. The network of anastomosing ridges is usually broken into radially arranged shorter ridges on the head shield plates. On the posterior margin of the PMD, on the subcephalic and subanal divisions of the AVL and PVL, the anastomoses between the ridges reduce, and nodose short ridges are present. The ornamentation on MV depends on the size of the plate: it is of reticular type in the smallest plates and becomes of the vermiculating type, i.e., short radiating nodose ridges, along the margins of the largest plates, whereas it remains of the reticular type in the central part. The ornamentation of the pectoral appendage is reticulate in general. On the anterior part of the Cd 1 and Cv 1 the ornament is radially arranged, whereas on the distal part of the proximal segment the ornamentation becomes smooth. The longitudinal striation in the ornament of the dorsal side of the distal segment is well shown in IG KSC 155/21 ( Fig. 13E View Fig 1 View Fig ).
Remarks.—This species was established by Traquair (1893) based on the material from Scotland and well-described by Stensiö (1948) and Miles (1968). Gross (1942) erected Bothriolepis curonica as a separate species from the lowermost Famennian of Latvia based on a small set of bones collected in 1934 in the Bienes locality. This description was repeated by Stensiö (1948) in Addenda to his monograph without further comments. Since then, the collection of the remains of this fish was significantly added by Vitaliy Sorokin and Lyubov Lyarskaya in 1975–1982 and by EL in 2013, who collected specimens from the Kalnamuiža locality in western Latvia. Specimens described by Gross (1942) and material collected from the Kalnamuiža site in 1970–80ies were briefly described and figured ( Lukševičs 1987), accenting the close morphological resemblance of B. curonica with B. leptocheira . A direct comparison of the specimens belonging to these species has indicated that they are conspecific ( Lukševičs 2001). Nevertheless, several features supported designation of the Latvian material to the subspecies Bothriolepis leptocheira curonica Gross, 1942 , which is morphologically very close to Bothriolepis leptocheira leptocheira from Scotland ( Miles 1968).
B. leptocheira jeremejevi ( Rohon, 1900) differs from the nominal subspecies B. leptocheira leptocheira from Scotland in its: (i) larger size; (ii) slightly narrower Pn plate; (iii) more acute posterior angle of PMD plate; (iv) slightly shorter ADL. It differs from B. l. curonica in its: (i) slightly narrower Prm and Pn plates, shorter and broader Pmg plate; (ii) slightly narrower AMD plate; (iii) relatively higher lateral wall of MxL and AVL plates; (iv) relatively shorter pectoral fin armour and separate plates of the fin; (v) coarser ornamentation. Overall proportions of the plates of the trunk armour and of the bones of the pectoral fin, distribution and path of sensory line canals as well as ornamentation of B. leptocheira jeremejevi are rather similar to those of B. leptocheira from Scotland and from the earliest Famennian of Latvia. The dissimilarities between the three subspecies mainly concern ornamentation, slightly differing proportions of some plates, relatively shorter but higher trunk armour, and shorter pectoral fins of B. leptocheira jeremejevi in comparison with B. leptocheira curonica .
Stratigraphic and geographic range.—Known only from the type locality and horizon.








| PVL |
Paleontologia de Vertebrados Lillo |
| IG |
Institute of Geology |
| MV |
University of Montana Museum |
| SM |
Sarawak Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Order |
|
|
Family |
|
|
Genus |
Bothriolepis leptocheira jeremejevi ( Rohon, 1900 )
| Lukševičs, Ervīns, Beznosov, Pavel & Stūris, Valdemārs 2017 |
Bothriolepis jeremejewi
| Gross, W. 1932: 34 |
Bothriolepis jeremejevi
| Rohon, J. V. 1900: 30 |