
Boophis piperatus, Glaw & Köhler & Riva & Vieites & Vences, 2010
|
publication ID |
https://doi.org/ 10.11646/zootaxa.2383.1.1 |
|
persistent identifier |
https://treatment.plazi.org/id/F566C51E-FF94-FFDC-E883-F8FF93EC1488 |
|
treatment provided by |
Felipe |
|
scientific name |
Boophis piperatus |
| status |
sp. nov. |
Boophis piperatus View in CoL sp. nov.
( Fig. 2 View FIGURE 2 , Appendix 9)
Remark. This species was referred to as Boophis sp. aff. rhodoscelis "Ranomafana" in Glaw & Vences (2007:152–153) and as Boophis sp. 30 by Vieites et al. (2009).
Holotype. ZSM 377 View Materials /2004 ( ZCMV 320 ), adult male (call voucher), from Ranomafana , Andranoroa river , near Ranomafanakely, 21°14.872' S, 47°22.580' E; 1138 m above sea level (a.s.l.), southeastern Madagascar, collected on 28 January 2004 by I. De la Riva and M. Vences. GoogleMaps
Paratypes. ZSM 378 View Materials /2004 ( ZCMV 328 ), female, same data as holotype GoogleMaps ; ZSM 379 View Materials /2004 ( ZCMV 338 ), male , ZSM 380 View Materials /2004 ( ZCMV 339 ), female , ZSM 381 View Materials /2004 ( ZCMV 341 ), adult male (call voucher) , UADBA 24317 View Materials ( ZCMV 342 ), sex unknown , ZSM 382 View Materials /2004 ( ZCMV 345 ), male , ZFMK 90151 About ZFMK (formerly ZSM 383 View Materials / 2004 [ ZCMV 346 ]), male, all with same data as holotype, but collected on 29 January 2004 GoogleMaps ; ZSM 299 View Materials /2006 ( ZCMV 3363 ), adult male (seen calling), Andranoroa river , no further data, but probably collected February / March 2006 .
Etymology. The specific name is a Latin adjective meaning "peppered", referring to the finely spotted dorsum, particularly evident in the holotype.
Diagnosis. Assigned to the genus Boophis based on the presence of an intercalary element between ultimate and penultimate phalanges of fingers and toes (verified by external examination), presence of nuptial pads and absence of femoral glands in males, absence of gular glands in males, enlarged terminal discs of fingers and toes, lateral metatarsalia separated by webbing, absence of outer metatarsal tubercle, and molecular phylogenetic relationships (see Vieites et al. 2009 for a complete molecular analysis of Boophis ). Assigned to the Boophis majori group based on the following combination of characters: small size (male SVL 28-30 mm); single subgular vocal sac; presence of vomerine teeth; smooth dorsal skin without folds, spines or tubercles; absence of distinct heel flaps or spines; presence of webbing between fingers; non-transparent ventral skin; brownish dorsal ground colour without green elements; occurrence of red colour on the hidden parts of the thighs and on the webbing of the feet. Boophis piperatus differs from all described species in the B. majori group by strong genetic differentiation (see below) and advertisement calls (see Glaw et al. 2001 and Vences et al. 2006, for comparative data on the calls of the other species). In brief, B. piperatus , B. feonnyala , B. miniatus , and B. picturatus differ from all other species in the group by having advertisement calls (call type 2 as described below) that are series of pulsed notes that directly follow each other, with a very short or almost missing inter-note interval. Among these species, the notes of B. piperatus are distinctly longer (135-283 ms) than those of B. picturatus (50-131 ms) and B. miniatus (35-85 ms), while no obvious difference exists between calls of B. piperatus and B. feonnyala . Morphologically, the new species differs from the three northern species of the Boophis majori group ( B. blommersae , B. marojezensis and B. vittatus ) by the presence of red colour on thighs and feet in preservative, and distinctly from B. pyrrhus and B. haematopus by the colouration in life and in preservative (see photographs in Glaw & Vences 2007). It differs from the syntopic B. majori by having larger size in males (SVL 27.9–29.6 mm vs. 21.5–24.5 mm) and females (SVL 36.2–36.9 mm vs. 29.0– 29.2 mm, see Glaw et al. 2001 for comparative data) and different colour in preservative (e. g. ground colour of back brown vs. reddish). It differs from B. miniatus and B. picturatus by different iris colouration in life (see photographs in Glaw & Vences 2007) and from B. feonnyala by slightly different size in males (SVL 27.9–29.6 mm vs. 25.0– 25.4 mm), a more strongly pointed snout, and lack of dark stripes or reticulations in the iris. It differs from the superficially similar species B. rhodoscelis and B. andrangoloaka by strong genetic differences and longer notes in advertisement calls (see call descriptions of those species below).
Description of the holotype. Adult male, SVL 28.7 mm. Body moderately slender; head slightly longer than wide, slightly wider than body; snout slightly subacuminate in dorsal view, moderately obtuse in lateral view; nostrils directed laterally, slightly nearer to tip of snout than to eye; canthus rostralis sharp in cross section, slightly concave in dorsal view, loreal region slightly concave; tympanum distinct, rounded, TD 56% of ED; supratympanic fold thin, distinct; vomerine odontophores distinct, well-separated in two elongated patches, positioned posteromedial to choanae; choanae medium-sized, elongated. Tongue posteriorly bifid, free. Arms slender, subarticular tubercles single, round; metacarpal tubercles not recognizable; fingers moderately webbed and with lateral dermal fringes; webbing formula 1(1), 2i(1.5), 2e(1), 3i(2), 3e(1.75), 4(1.25); relative length of fingers 1<2<4<3 (finger 2 distinctly shorter than finger 4); finger discs enlarged. Hindlimbs slender; tibiotarsal articulation reaching a little beyond tip of snout when hindlimb is adpressed along body; lateral metatarsalia separated by webbing; inner metatarsal tubercle small, distinct, elongated; no outer metatarsal tubercle; toes broadly webbed; webbing formula 1(0.25), 2i(0.5), 2e(0), 3i(0.75), 3e(0.25), 4i(1.25), 4e(1.25), 5(0.25); relative length of toes 1<2<5=3<4; toe discs enlarged. Skin finely granular on dorsal surfaces, smooth on throat, chest, and ventral surface of thighs, coarsely granular on belly. A tissue sample was removed from the right thigh.
Measurements (in mm): SVL 28.7, HW 10.2, HL 10.8, ED 4.1, END 2.5, NSD 2.1, NND 2.9, TD 2.3, TL 16.4, HAL 9.1, FOL 13.1, FOTL 22.7.
After almost four years in preservative, ground colour of upper surface of head, dorsum and legs pale brown with tiny cream and dark brown flecks; upper lip cream with brown flecks; a canthal dark brown stripe from eye to tip of snout, outlined by a cream line above; a pale brown triangle from interorbital region to tip of snout. Tiny red flecks on tip of snout, supratympanic fold and dorsal surface of digits; posterior surface of thighs with irregular, diffuse red markings; lower flanks and ventral surfaces creamy yellow.
In life, dorsal surfaces pale brown with scattered small, darker flecks. Lower flanks cream with irregular greenish grey blotches. A paler triangle on head and a dark canthal stripe. Upper surfaces of thighs and shanks the same colour as dorsum, with darker diffuse bars. Iris golden with brown irregular markings around the pupil; iris periphery black, followed by green posteriorly ( Fig. 2 View FIGURE 2 ). Ventral life colouration of holotype unknown.
Variation. Morphometric variation is given in Appendix 1. Overall, paratypes are similar to the holotype. There can be large cream blotches on upper lip; females are larger and red colouration in preservative is more intense and extended than in males. At least one specimen was observed calling in the wild (emitting notes of type 1) and had a single subgular vocal sac that was moderately inflated during calls.
Natural history. As reported first by Vences et al. (2006) under the name " Boophis sp. aff. rhodoscelis (Ranomafana) ", calling males of this species were found at night, 1–2 m high in the vegetation along Andranoroa river, in a rather disturbed area next to the edge of rainforest. Some single specimens were emitting call type 1 as described below while others were emitting call type 2 described below. Call type 2 was emitted in "waves" by a chorus of numerous specimens. Call type 3 that Vences et al. (2006) preliminarily assigned to this species in fact belongs to the syntopic Boophis sp. aff. majori "long calls" ( Glaw & Vences 2007). Specimens were regularly encountered at the same site during visits in 2004 and 2006. They often were sitting on leaves on tree branches overhanging the water, sometimes also in bushes at some meters distance from the river edge.
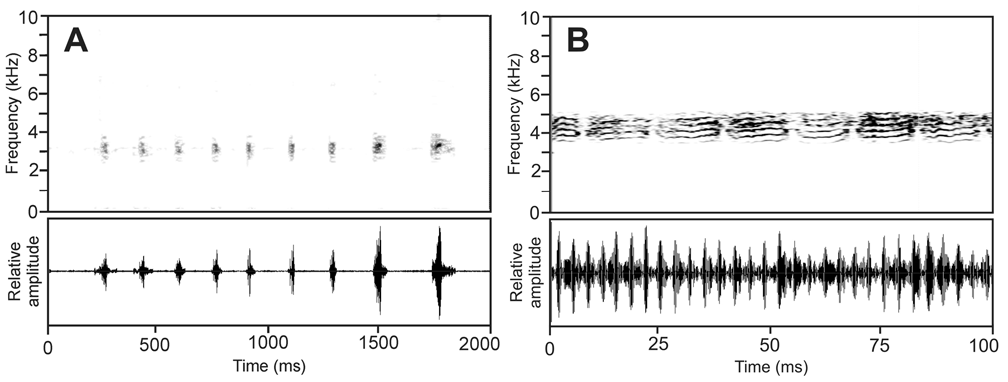
Vocalization. Calls of B. piperatus may contain two different call types ( Fig. 3 View FIGURE 3 ). Type 1 consists of a series of pulsed notes emitted in fairly regular intervals. Each note contains 2–8 indistinct pulses. Within the call, notes show an increase in amplitude and duration, with the two terminal notes usually being the longest and loudest. Call type 2 is a vocalization with strongly pulsed notes, separated by only very short intervals. Compared to call type 1, it is significantly higher in frequency and several parallel narrow frequency bands are present in the spectrogram. Numerical parameters for the different call types of the Ranomafana population are as follows: Call type 1: call duration, 1100–2000 ms (1474 ± 336; n = 5); note duration, 18–50 ms (34 ± 10; n = 17); inter-note intervals, 121–310 ms (163 ± 47; n = 18); note repetition rate, 5.4–6.0 notes/second; dominant frequency range 2800–3500 Hz, maximum call energy at 2870–3180 Hz. Call type 2: call duration, 3260–3300 ms (n = 2); note duration, 135–283 ms (253 ± 38; n = 16); inter-note interval, 29–63 ms (43 ± 9; n = 15); note repetition rate, 3.1–3.4 notes/second; pulse rate approximately 300 pulses/second; dominant frequency range, 3600–5200 Hz, maximum call energy at 4170–4710 Hz ( Vences et al. 2006, CD 1, track 57, call sections 1 & 2).
Molecular relationships. According to the phylogenetic tree ( Fig. 1 View FIGURE 1 ) this species belongs into a clade that also contains B. arcanus (described below), B. feonnyala , B. haematopus , B. majori , B. miniatus , B. picturatus , and B. pyrrhus . This clade receives only very low support [56% bootstrap support (BS); Bayesian posterior probability (BPP) not significant (<0.95)], and relationships among the included species are largely unresolved. This clade largely corresponds to the Boophis majori group (see Glaw & Vences 2006), except for three species ( B. blommersae , B. marojezensis , and B. vittatus ) that are grouped with high support in a separately placed clade (BS 70%, BPP significant). Apparently, the B. majori group sensu lato consists of two lineages, one (the B. majori group sensu stricto) that is here placed sister to the B. microtympanum group (with B. andrangoloaka , B. laurenti , B. microtympanum , B. rhodoscelis , B. williamsi ; BS 96%, BPP significant), and a second clade with B. blommersae , B. marojezensis , and B. vittatus that may merit a status as separate species group. The molecular differentiation among all species in the B. majori group sensu stricto is high. Boophis piperatus is genetically highly divergent: pairwise distances are 5.3% to B. miniatus , 7.7% to B. majori , 8.5% to B. feonnyala , and>7% to all other species in the clade. All sequenced individuals of B. piperatus have identical sequences in the 16S rRNA fragment studied. The analysis also indicates divergences (2.8%) among sympatric specimens assigned to B. majori sensu lato, indicating that the specimens named B. sp. aff. majori "long calls" by Glaw & Vences (2007) (with strongly divergent call structure) may represent a further distinct, undescribed species (see also Vieites et al. 2009).
Distribution. At present Boophis piperatus is only reliably known from its type locality, Andranoroa river near Ranomafanakely, in the Southern Central East of Madagascar, along the road connecting Vohiparara with National Road RN7 Antananarivo-Fianarantsoa (Appendix 10).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.