
Baetis (Baetis) zdenkae Soldán and Godunko, 2009
|
publication ID |
https://doi.org/ 10.5281/zenodo.185067 |
|
DOI |
https://doi.org/10.5281/zenodo.6223129 |
|
persistent identifier |
https://treatment.plazi.org/id/1E3887F1-C07B-4F30-FF03-FD7FFCCFFE49 |
|
treatment provided by |
Plazi |
|
scientific name |
Baetis (Baetis) zdenkae Soldán and Godunko |
| status |
sp. nov. |
Baetis (Baetis) zdenkae Soldán and Godunko View in CoL , sp. nov.
( Figs 1–29 View FIGURES 1 – 4 View FIGURES 5 – 7 View FIGURES 8 – 12 View FIGURES 13 – 16 View FIGURE 17 View FIGURES 18 – 21 View FIGURES 22 – 25 View FIGURES 26 – 29 )
Description. Male imago. Size: body length: 5.8–6.3 mm; fore wings length: 5.6–6.2 mm; cerci length: 10.6– 13.8 mm; tarsal segments: T1 = 0.68–0.72 mm; T2 = 0.54–0.57 mm; T3 = 0.32–0.34 mm; T4 = 0.23–0.25 mm; general relative tarsal segments length ratio: 1>2>3>4.
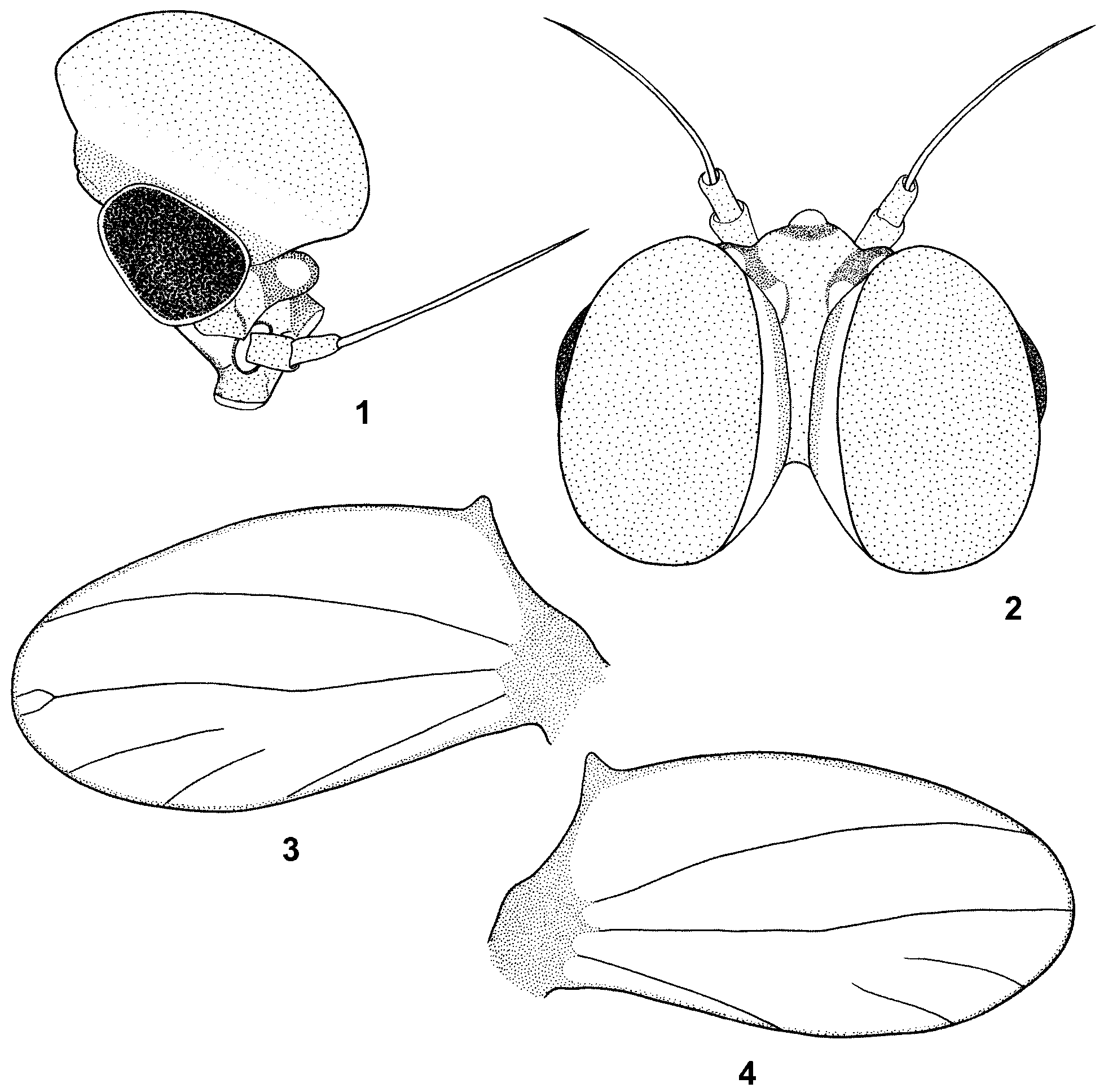
General body color light, yellowish-brown to brown. Head brown with darker smudges laterally. Antennae brown, flagellum slightly paler. Ocelli whitish-brown at the apex with brown bases. Eyes black, surrounded by a light ring, occasionally with some yellowish smudges basally. Turbinate eyes oval in dorsal view with convex external margins. Facetted surface yellowish-brown to light brown. Distal part of stalk with narrow yellow ring; central part slightly darker, dirty-yellow, basal brown ring well distinguished ( Figs. 1, 2 View FIGURES 1 – 4 ). Thorax darker than head, yellowish-brown to brown, with light to dark brown maculation along sutures. Pronotum and distal part of mesonotum yellowish-brown to brown, proximal part distinctly brown. Thoracic sterna light brown to brown. Wings hyaline, transparent. Fore wings with slightly opaque pterostigma. Hind wings with costal process small, acute and rounded at apex with three simple longitudinal veins with two free veins in the third field, second vein occasionally with distal bifurcation, third vein relatively short, length about 0.38–0.40 of wing ( Figs. 3, 4 View FIGURES 1 – 4 ). Fore legs light brown, femora with diffuse smudges distally, middle and hind legs yellow to yellowish-brown.
Abdominal terga with small brownish lateral smudges. Tergum I brown, terga II–VI whitish-yellow with a pair of diffuse central brownish smudges, terga VII–X distinctly darker, yellowish-brown, with narrow elongate central spots. Sterna whitish-yellow to dirty yellow with elongate whitish central spot on segments VII– VIII.
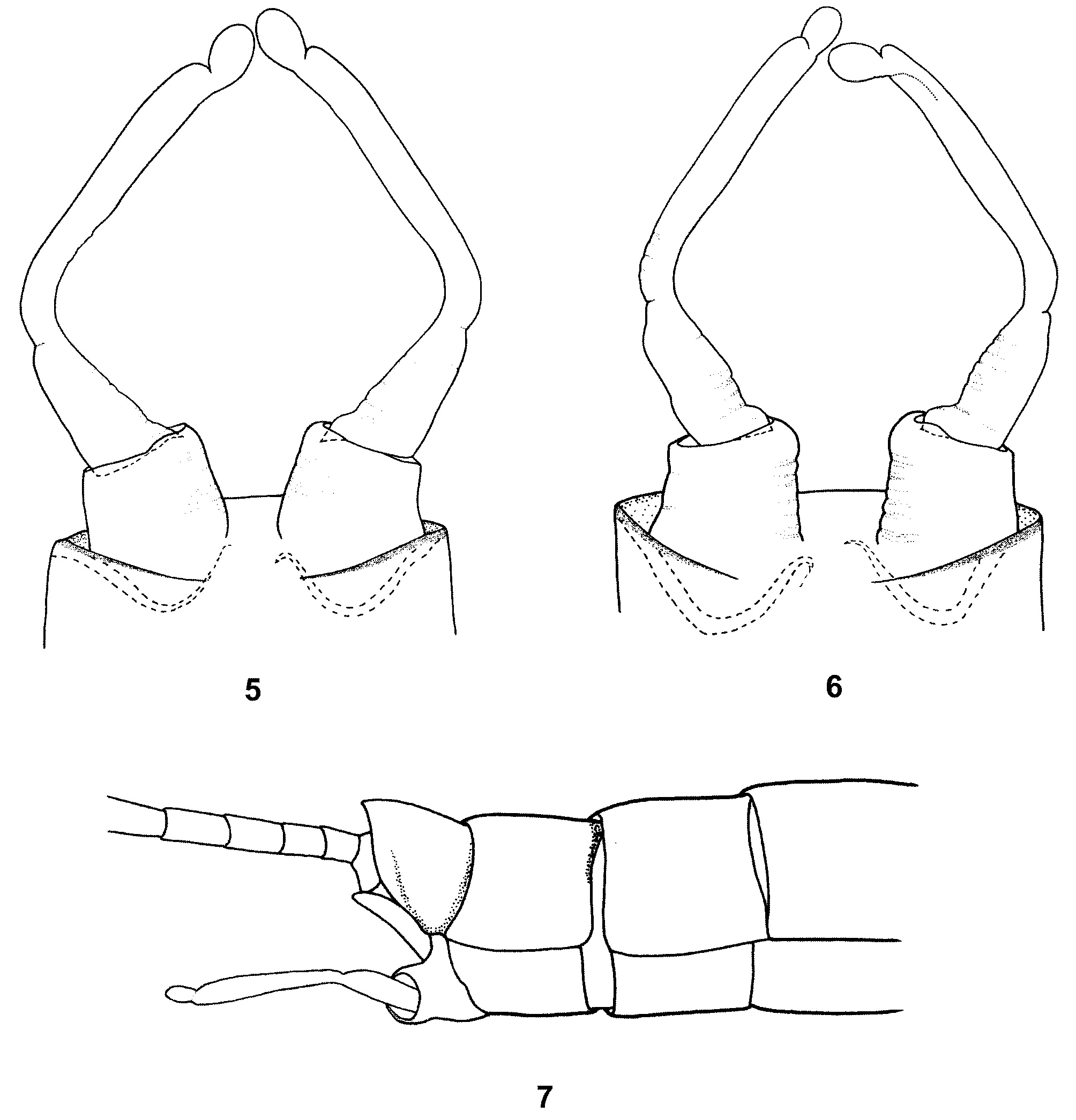
Genitalia pale, yellowish-white to dirty yellow. Forceps distinctly bent up at approximately 45° angle in lateral view ( Fig. 7 View FIGURES 5 – 7 ). Basal segment about as long as wide without a posteromedial projection, segment 1 elongate conical with slightly convex external margin, segment 2 narrow at the base, slightly expanded distally (widened part as long as 2/3–3/4 of the segment length), segment 3 elongate, slightly longer than wide ( Figs. 5, 6 View FIGURES 5 – 7 ). Cerci whitish-grey to dirty yellow, first 4–7 segments light brown.
Female imago. Size: body length: 6.5 mm; fore wings length: 6.2 mm; cerci length: 13.2–13.4 mm.
General body color yellowish-brown to brown. Head uniformly brown. Eyes black with yellowish maculations. Ocelli yellowish with black bases, antennae brown.
Thorax yellowish-brown to brown with dark brown smudges on mesonotum. Sterna whitish-yellow to light brown. Legs yellowish-brown to light brown, fore legs darker. Wings transparent, hyaline, fore wings with distinct yellowish-brown venation, hind wings similar to male, veins darker.
Abdominal terga uniform, yellowish-brown to light brown, sterna paler. Cerci light brown to whitish-yellow, darker at bases.
Mature larva. Size: body length: 5.4–7.5 mm (male), 5.8–8.5 mm (female); cerci length: 2.0– 3.3 mm.
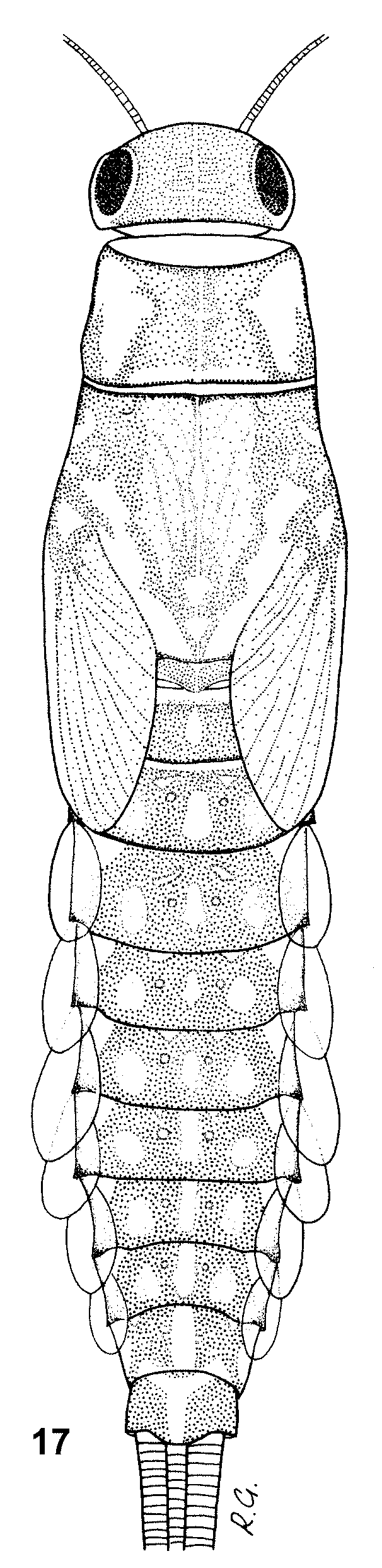
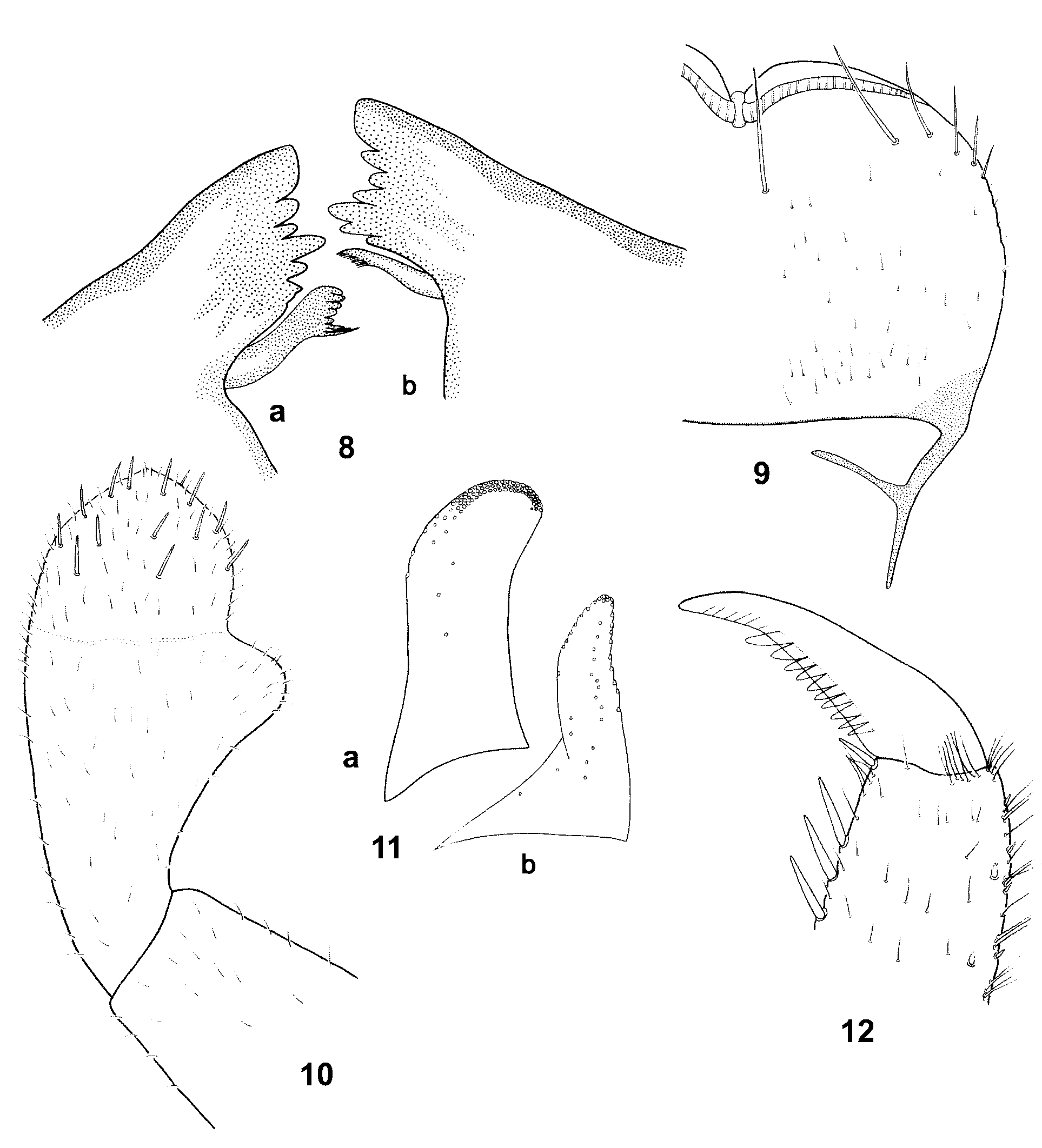
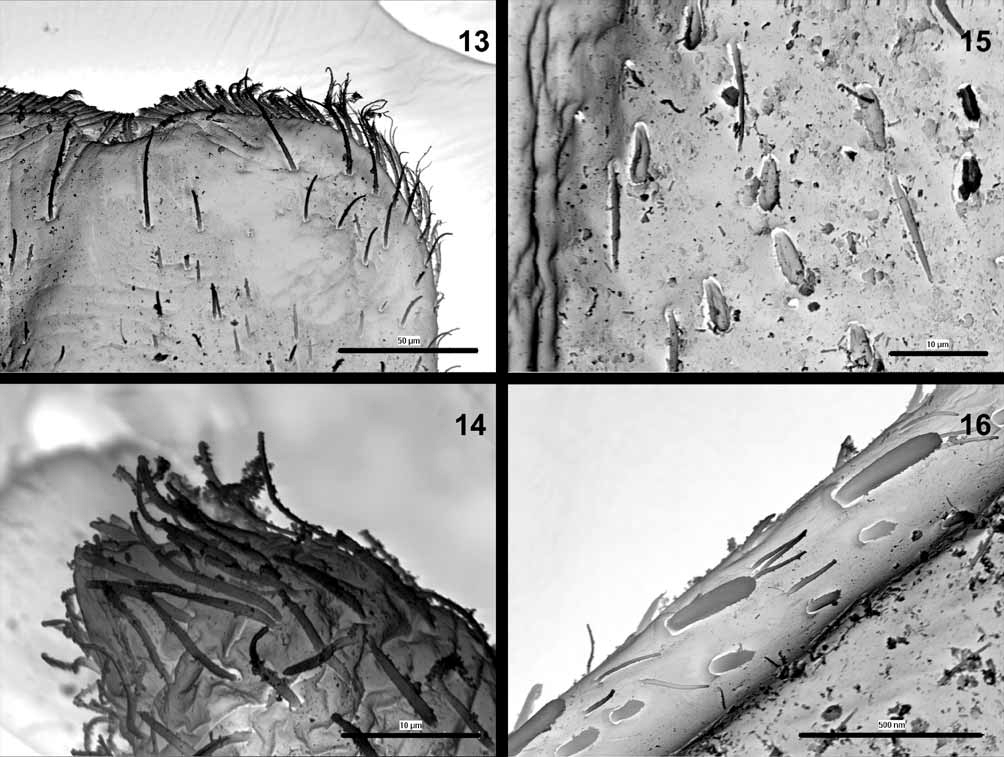
General body color dark, light brown to dark brown. Head yellowish-brown to brown, frons and clypeus paler. The whole head surface covered with short setae and solitary bases of setae. Surface of clypeus with numerous setae. Eyes black, developing turbinate eyes light brown to brown, surrounded by a narrow yellowish ring. Antennae whitish-yellow to light brown, flagellum paler than pedicel and scape, with numerous fine hairs. Pedicel surface with short setae only, scape surface with solitary setae concentrated distally. Labrum relatively wide (width/length ratio 1.52–1.55), with 1 + 4–7 long submarginal bristles and a few pointed lateral setae ( Figs. 9 View FIGURES 8 – 12 , 13 View FIGURES 13 – 16 ). Mandible incisors as in Figs 8 View FIGURES 8 – 12 a, b. Tip of maxillary palpus conical with barely visible, small apical spine, surface with numerous fine hairs ( Fig. 14 View FIGURES 13 – 16 ). Segment 2 of labial palpus relatively broad with slightly concave inner margin, about twice as broad at apex than at base, distomedial lobe as wide as about 1/ 3 of segment 3 at the base. Segment 3 slightly asymmetrical, slightly longer than 1/2 length of segment 2. The length/width ratio of segments 2 and 3 combined is 1.8–2.1 ( Fig. 10 View FIGURES 8 – 12 ). Glossae and paraglossae wide, robust. Paraglossae with 3 regular rows of long bristles apically, basal row consisting of at least 16 bristles ( Figs. 11 View FIGURES 8 – 12 a, b).
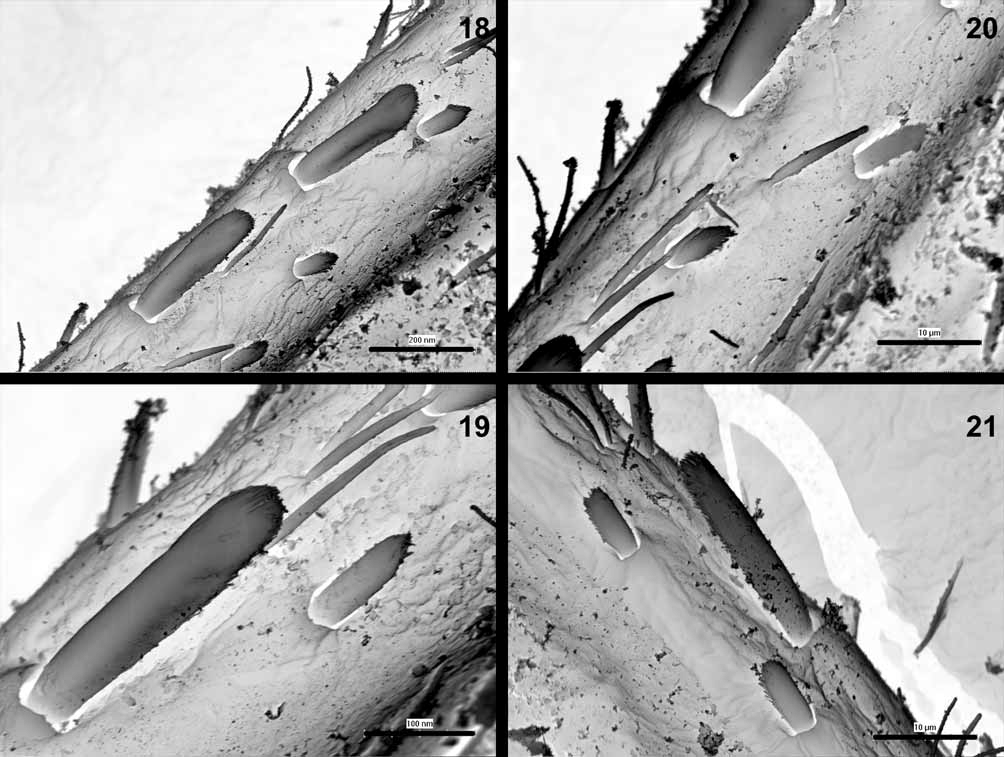
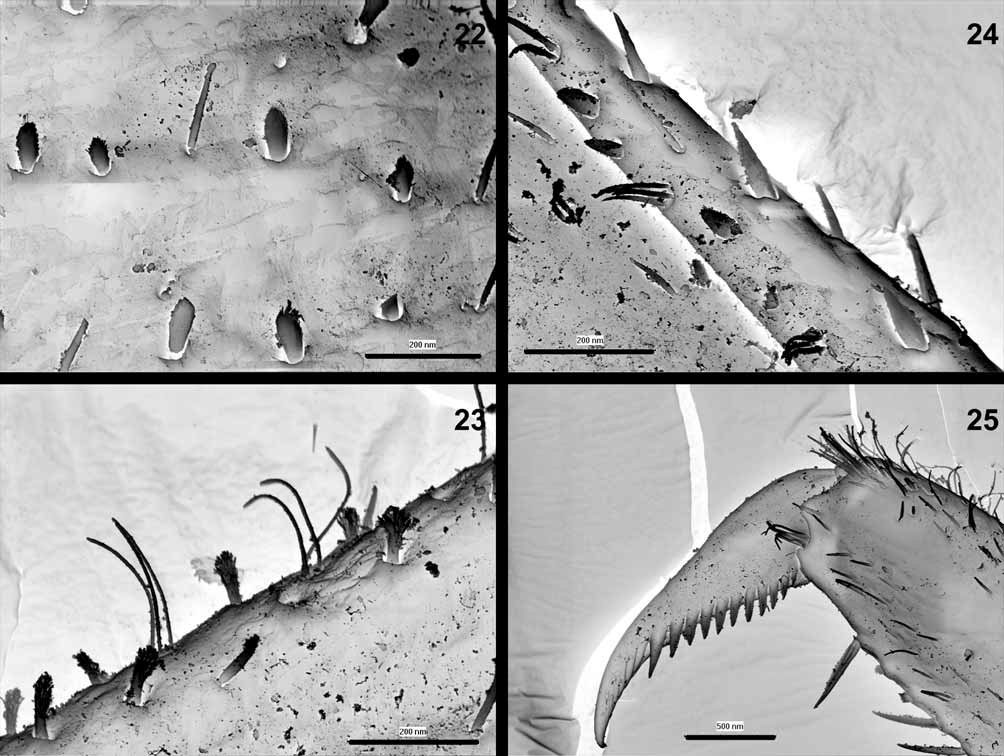
Thorax dark, light to dark brown, pronotum with a pair of large pale subtriangular areas laterally, mesonotal color pattern as in Fig. 17 View FIGURE 17 . Metanotum brownish with central pale spot. Thorax surface with numerous fine hairs and slender spatulate setae rounded apically ( Fig. 15 View FIGURES 13 – 16 ). Sterna pale, yellowish to light brown. Legs yellowish to light brown with distinct large central dark spot on all femora (occasionally with relatively small central spot). Tibiae with brown distal spots (occasionally only with diffuse brownish smudges), tarsi darker distally. Outer margin of femora with a single, sparse row of large spatulate bristles alternating with fine hairs and small submarginal spatulate setae ( Fig. 16 View FIGURES 13 – 16 ). Spatulate bristles relatively short and robust, distinctly rounded at apex, distal portion with shallowly splintered margins ( Figs. 18, 19, 21 View FIGURES 18 – 21 ). Femora with (1) slender spatulate setae with convergent margins ( Figs. 18–20 View FIGURES 18 – 21 ) and (2) relatively wide and apically rounded spatulate setae with subparallel margins ( Fig. 21 View FIGURES 18 – 21 ). Submarginal spatulate setae of distal portion of femora with splintered margins. Inner margin of femora with uniform row of small pointed spines and fine hairs; all surface of femora with spatulate setae and fine hairs ( Fig. 22 View FIGURES 22 – 25 ). Outer margins of tibiae with deeply splintered, slender spatulas alternating with fine hairs ( Fig. 23 View FIGURES 22 – 25 ). Inner margins with irregular row of spines and fine hairs ( Fig. 24 View FIGURES 22 – 25 ). Outer margins of tarsi with small pointed spines and hairs; small submarginal spatulate setae, with distally splintered edges. Inner margins with large pointed spines; splintered submarginal spatulate setae, and fine hairs. Surface of tarsi with numerous hairs and splintered spatulate setae. Tarsal claws elongate, relatively slender with at least 10 teeth, without subapical setae ( Figs. 12 View FIGURES 8 – 12 , 25 View FIGURES 22 – 25 ).
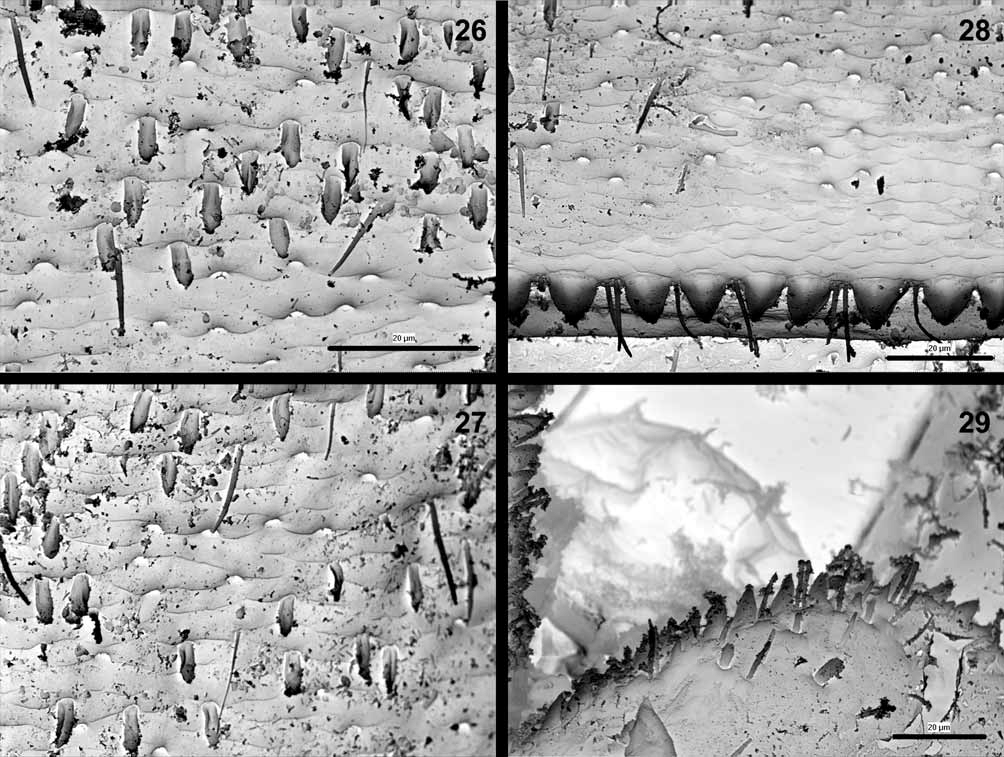
Abdominal terga dark, yellowish-brown to brown, color pattern as in Fig. 17 View FIGURE 17 , muscle insertions pale (cf. Jacob 2003), posterior margins with broad triangular bluntly pointed teeth alternating with setae ( Fig. 28 View FIGURES 26 – 29 ), surface with numerous fine hairs, wide bases of hairs and small spatulate setae. Two distinct types of spatulate setae discernible on abdominal tergal surface: (1) elongate narrow ones with lateral convergent margins, and (2) relatively rare ones with subparallel or divergent lateral margins, rounded at apex ( Figs. 26, 27 View FIGURES 26 – 29 ). Sterna uniform, yellowish-brown to brown with a pair of short transversal divergent strokes near the anterior margins. Inner margin of paraprocts with a single row of at least 10 (usually 12–15) distinct teeth ( Fig. 29 View FIGURES 26 – 29 ), surface with fine hairs and sparse spatulate setae. Gills 1 and 7 slightly asymmetric and subequal in size. Gills 2– 6 asymmetric and distally rounded. Both outer and inner gill margins with small teeth alternating with fine hairs. Cerci uniform, yellowish-brown to brown, paracercus shorter by 1/2.
Male and female subimagos unknown.
Etymology. The species is named for Zdenka, wife of the senior author, to acknowledge her distinguished field assistance during sampling of mayflies on Rhodos.
Types. HOLOTYPE: male imago, GREECE, Rhodos (Rodos) Island, stream at the Kremastis Bridge in Kremastí, about 14 km SW of Rhodos City, about 3 km NE of the Diagoras International Airport and about 500 m from its mouth to the Aegean Sea at about 10 m a. s. l., May 28, 2005. PARATYPES: 6 male imagoes, 1 female imago, 275 larvae, same locality and collection date as holotype.
Further material examined (no types). 256 larvae, GREECE, Rhodos (Rodos) Island, same locality and collection date as holotype; 8 larvae, Greece, Rhodos Island, Argiros Potamos (stream), about 30 km SW of Rhodos City, 1 km NE of Kalavárda and about 1 km from its mouth to the Aegean Sea at about 10 m a. s. l., May 26, 2005.
All material (preserved in 70% alcohol) was collected by Zdenka and Tomáš Soldán. Genitalia of two male imagoes on slides. Holotype, most paratypes and other material examined deposited in collection of the Biological Centre, Academy of Sciences of the Czech Republic, Institute of Entomology, Č eské Budĕjovice, Czech Republic; 50 larvae and 1 male imago (all paratypes) deposited in the State Museum of Natural History, National Academy of Sciences of Ukraine, Lviv, Ukraine.
The comparative material of B. buceratus was collected in Central Bohemia (the Berounka River basin), Slovakia and Ukraine (the Latorica, Tisza and Dniester River basins) at numerous localities. Larvae and adults of B. nexus were collected at several localities of the Dyje (Thaya) River basin in Moravia ( Czech Republic), Slovakia and Ukraine (the Latorica and Tisza River basins). For descriptions of some localities see Landa and Soldán (1989), Soldán and Zahrádková (2000), and Horsák (2001). Material of B. spei was not studied, morphological characters and other data are based on the original description by Thomas and Dia (1985, 2007).
Differential diagnosis and affinities. B. zdenkae sp. nov. can be easily distinguished from the other three representatives of the B. buceratus species-group ( B. (B.) buceratus Eaton, 1870 , B. (B.) nexus Navás, 1918 , and B. (B.) spei Thomas & Dia, 1985 ) by the following combination of morphological characters: in male imago: (1) facetted surface of turbinate eyes yellowish-brown to light brown (yellowish brown to orange brown in B. buceratus , dark brownish in B. nexus and flesh-colored in B. spei ), (2) tricolor combination of stalk of turbinate eyes (stalk paler, light orange in B. buceratus , eyes unicolorous in B. nexus , stalk flesh-colored in B. spei with diffuse and intermittent basal ring), (3) third vein of hind wings relatively short, about 0.38–0.40 wing length (about half of wing length in B. buceratus , 1/2 length to longer in B. nexus ), (4) forceps conspicuously bent up in lateral view (as in B. buceratus , nearly straight in B. nexus and B. spei ), (5) basal segment of forceps without posteromedial projection (posteromedially produced into lobe-like projection in B. buceratus , B. nexus and B. spei ), and (6) segment 1 of forceps elongate conical (conical in B. buceratus and B. spei , oval, bulbous with apparently convex inner and outer lateral margins in B. nexus ); in larva: (7) structure of segments 2 and 3 of labial palpi (segment 3 shorter than or equal to 1/2 of segment 2 in B. nexus ), (8) structure of mandible incisors (cf. e.g., our Figs. 8 View FIGURES 8 – 12 a, b with Fig. 3 View FIGURES 1 – 4 g, d by Thomas and Dia 1985: 242 showing B. spei ), (9) paraglossae with basal row of apical bristles consisting of at least 16 bristles (about 10–12 bristles in B. spei ), (10) outer margins of femora with single sparse row of spatulate bristles, alternating with fine hairs and submarginal small spatulate setae (as in B. spei ), (11) structure of small spatulate setae of femora (some of them conspicuously splintered as in B. spei ), (12) arrangement of small spatulate setae on dorsal surface of femora, (13) posterior margins of abdominal terga with broad bluntly pointed triangular teeth, alternating with setae (teeth rounded, semicircular in B. spei , narrow triangular blunt pointed teeth in B. buceratus ), (14) surface of terga with numerous fine hairs, spatulate setae, and wide bases of hairs and spatulate setae (without spatulate setae in B. spei ), (15) inner margin of paraproct with 10–15 distinct teeth (about 20–25 teeth of different shape in B. spei ; about 20 teeth in B. buceratus and Thomas and Dia 1985: 243, Fig. 12 View FIGURES 8 – 12 ; about 25 teeth in B. nexus, Müller-Liebenau 1969: 149 , Abb. 109), (16) paracercus shorter than cerci by 1/2 (subequal in B. buceratus and B. nexus , about 2/3 of cerci in B. spei ), and (17) specific color pattern of thorax and abdomen in dorsal view (e.g., a single, conspicuous pale band in the middle of pronotum in B. spei , in Thomas and Dia 1985: 243, Fig. 7 View FIGURES 5 – 7 ).
The male genitalia of B. zdenkae sp. nov. are similar to B. buceratus , in the structure and proportions of segments 1–3, and conspicuously bent up forceps (see Müller-Liebenau 1969: 140, Abb. 99–100 or Jacob 2003: 96, Abb. 17a–b). On the other hand, the male imago of the new species differs markedly from B. buceratus in color pattern of the facetted surface of the turbinate eyes and their stalks and in hind wing venation (cf. characters 1–3, see also Müller-Liebenau 1969: 139–140, Abb. 104j or Jacob 2003: 97). On the other hand, B. zdenkae sp. nov. can be easily distinguished from B. nexus by the different arrangement of all critical imaginal characters (1–6, see above and cf. Müller-Liebenau 1969: 146–147, Abb. 101, 107, and 108, or Jacob 2003: 98, Abb. 17c).
Larvae of B. zdenkae sp. nov. can be distinguished from all remaining representatives of the B. buceratus species-group by the combination of characters (9 and 14), and, particularly really the critical characters (8 and 16) separate this species from B. buceratus and B. nexus . Additionally, B. zdenkae sp. nov. differs from B. nexus by the structure of outer margin of femora and paraproct (characters 10 and 15). The latter two characters and the arrangement of triangular teeth on the posterior margin of abdominal terga (character 13), distinguish the new species from B. spei with small rounded, nearly semicircular teeth, cf. Thomas and Dia (1985: 243: Fig. 11 View FIGURES 8 – 12 ). Contrary to B. spei (muscle insertions on head well visible and contrasting), the head of B. zdenkae sp. nov., is rather unicolorous. Labrum of B. zdenkae sp. nov., provided with 1 + 4–7 bristles is relatively wider (width/length ratio 1.52–1.55) vs. 1 + 6–8 bristles and width/length ratio 1.35–1.50 in B. spei . Further differences between B. zdenkae sp. nov., B. buceratus and B. spei can be found also in thoracic and abdominal terga color patterns (see Müller-Liebenau 1969: 141–145 and 147–150, Abb. 102–103, 109 or Jacob 2003: 96–98, Abb. 3l, 18, 19, and Thomas and Dia 1985 for details).
Biology. Larvae of B. zdenkae sp. nov. evidently prefer stream run places of the “lowland” biotopes within the island. These localities (mostly at elevations of 100–250 m a. s. l.) were inhabited by other baetid species, e.g., by apparently dominant Baetis (Nigrobaetis) digitatus ( Bengtsson, 1912) which is the first faunistic record from Rhodos and P. fascicaudale , but no larvae of B. zdenkae sp. nov. have been collected. Larvae of this new species were found in two streams with permanent water of about 1–3 m across and an average depth of 10–30 cm, moderate to fast current velocity and stony bottom overgrown, in some places, with submerged aquatic vegetation. The current velocity was about 20–50 cm.s -1 at the most densely inhabited places, the density of larvae, roughly estimated at about 100–300 ind.m -1, decreased rapidly with decreasing current velocity, places with slower current were inhabited rather sparsely. Water temperature ranged from 17 ºC early in the morning to 23–26.5 ºC in the afternoon at sun-exposed, densely inhabited places (type locality, up to 28.5 ºC in pools with very rare occurrence of larvae). As to substrate roughness, larvae preferred gravel places (lithal) covered with cobble and pebble. Their densities in places covered with granules (2.0 mm in diameter) and very coarse (1.0 mm) or coarse (0.5 mm) sand were very low showing rather incidental occurrence. Larvae were not found on fine substrates like medium to very fine sand or silt. If submerged aquatic plants occurred at the places preferred, larval densities among vegetation were comparable with those of stony habitats. However, substrate requirements of larvae do not seem to be pronounced. Stream banks are regulated at numerous places in both biotopes inhabited. For instance, the Argiros Potamos near Kalávarda is a very shallow stream with an artificial concrete streambed in some places.
At the type locality, larvae of B. zdenkae sp. nov. are clearly dominant forming at least 75% of the mayfly standing crop, they seem to be very rare at the locality of the Argiros Potamos (at most 10% of the mayfly standing crop in May, Caenis luctuosa ( Burmeister, 1839) and a new species of the B. lutheri species-group were dominant). At the type locality, the larvae were collected together with those of C. luctuosa , P. fascicaudale and B. (Rhodobaetis) cf. rhodani ( Pictet, 1843) .
The life cycle of B. zdenkae sp. nov. remains unknown in detail. However, judging from the size of larvae in May 2005 and timing of mating flight we might suppose a seasonal, bivoltine winter life cycle, the MBws type according to the classification by Clifford (1982). Adult emergence by mid May indicates a possibility of two generations a year (second generation might fly from late August to early October). Owing to favorable conditions (generally mild climate, relatively high water temperatures) there is a chance to complete development of larvae originating from the eggs laid in May of the same year. Taking into account approximately linear growth of larvae during winter followed by its acceleration after spring warming of water, and the different size categories of larvae collected at the same time in May, several egg cohorts are likely, as in numerous other mild climate Baetis species (cf. Sowa 1975 or Clifford 1982)
Subimagines emerge from the water surface and places overgrown with submerged vegetation during daylight hours, from about 10.00 a.m–16.00 p.m. local time. Baetis zdenkae sp. nov. showed early morning mating flight activity with the mating flight generally occurring from about 6.30–8.30 a.m. local time. First males started to appear approximately a half hour before sunrise, and showed a typical “pendular” flight pattern usually in groups of about 3–10 individuals, females were observed about 0.5–1 hour later. Although only a few females were observed flying upstream, their flight activity seemed to last at least one hour after males had disappeared. A single female was observed ovipositing on a submerged stone, hence the way of oviposition seems to be similar to other species of the genus Baetis .
Generally, the biology of B. zdenkae sp. nov. resembles that of B. spei which was characterized as a rhithro-potamal species by Thomas and Dia (1985), while the remaining representatives of B. buceratus species-group occur at rather different (mostly potamal) habitats. However, larvae of B. spei showed a wider range of vertical distribution than those of B. zdenkae sp. nov. being collected at altitudes of 5–950 m a. s. l. at places 2–48 km from source in well oxygenated water. They were moderately abundant to abundant at streams about 3–10m across, including also a temporary stream. The larvae were characterized as thermophilous (in contrast to those of B. bisri Thomas & Dia, 1983 and B. baroukianus Thomas & Dia, 1984 living at the same area ( Thomas & Dia 1983, 1984), maximal water temperature ranged from 17–29 ºC (pH 7.4–8.25), conductivity 290–440 µS.cm -1. The life cycle of B. spei is not fully known, mature larvae were collected from May to August ( Thomas & Dia 1985), were collected from May to August ( Thomas & Dia 1985), and adults were collected at the same time ( Thomas & Dia 2007).
Larvae of B. buceratus have been found mostly at the localities of large lowland rivers (epipotamal). They apparently can tolerate a certain degree of pollution being sometimes dominant at these biotopes. Larvae prefer medium to high current speed and stony substrata (gravel and larger stones) but often occur also among submerged plants. Such habitats are described as typical for this species ( Buffagni & Desio 1994; Gonzáles del Tánago 1984). B. buceratus shows a typical bivoltine winter life cycle (MBws) with overwintering larvae of the first generation. However, there are some data indicating plasticity of the cycle. In Switzerland, larvae live also in epirhithral stream segments although they prefer lower elevations and places with lower current velocity at this area ( Sartori & Landolt 1999). According to Landa (1968) older larvae of the first generation appear from March to May – this may indicate ceasing of larval development during winter months rarely found in species with the MBws life cycle type. According to Haybach (1998, 2006) there is one overwintering generation with flight period in spring and two generations following late summer and autumn. Since the adults apparently fly from July to October ( Landa 1968; Langford 1971; Sowa 1975), two “summer” generations and, consequently, some type of polyvoltinism (MP) cannot be excluded.
Larvae of B. nexus live in lowland waters of different size, being mostly found in rivers, their backwaters and artificial freshwater biotopes (e.g., in the “roach zone”), at places with very low current velocity. They evidently prefer places rich in submerged aquatic vegetation. Azonal species according to Haybach (1998, 2006), habitat preference cannot be evaluated from the rhithron-potamon concept. However, in south Moravia ( Czech Republic), the larvae are usually found in a typical epipotamal river, moreover influenced by the upstream impoundment ( Horsák 2001) like they do in southern France (Cammouseight & Fontaine 1990, see this paper also for further details on ecological requirements of larvae and their quantitative presentation). Seasonal bivoltine winter life cycles (MBws) have been observed in France (Cammouseight & Fontaine 1990), with two flight periods in May and August, and also in Hungary ( Ujhelyi 1966), Germany ( Haybach 1998, 2006) and Poland ( Keffermüller 1972).
Distribution. Since B. zdenkae sp. nov. is so far known only from Rhodos Island it might be considered endemic to this island. However, the true distribution of B. zdenkae sp. nov. still remains poorly known so that its classification is preliminary since these possible “endemics” might inhabit a much larger area as shown, for example, by our finding of presumably “endemic” species B. mirkae group described from Cyprus also on Rhodos ( Soldán & Godunko, 2008). That is why we prefer to wait until further collection records are published. The distribution of B. zdenkae sp. nov. can be compared to that of B. spei , the species known only from Lebanon, only from two river basins (Nahr el Aouali and Nahr ed Damour) as well, so far altogether from 9 localities ( Thomas & Dia 1985).
On the other hand the distribution of B. zdenkae sp. nov. seems to be quite different from the distributions of B. buceratus and B. nexus . The former species (type locality the Kenneth and Holybrook near Reading, Berkshire, England) belongs to widely distributed Westpalaearctic species. It is known from Europe including the British Islands and some Mediterranean islands, e.g., Sardinia ( Belfiore & Gaino 1988) but probably missing in North Africa. Northern limits are situated in Scandinavia – Sweden and Denmark ( Bengtsson 1917; Jensen 1969), southeastern ones in Asia Minor, Turkey and Iraq ( Koch 1988; Kazanci 1984; Al-Zubaidi et al. 1987), eastern limits probably in southern Ural, Caucasus and Central Asia (cf. Kluge 1997). B. buceratus is classified either as a Holomediterranean faunistic element of the expansive type according to Jacob (1972), or a Pontokaspian element according to Haybach (1998) and its area is sympatric to that of B. zdenkae sp. nov.
Baetis nexus View in CoL belongs to widely distributed Westpalaearctic species as well, however its area is restricted to about 50º N. Northern limits are situated in Germany and Poland. This species is known from Europe except for the British Islands, Fennoscandia and most of the Mediterranean, and is missing in North Africa (see Sartori & Soldán (2006) for complete list of distributional records). Eastern limits are uncertain, the species has been found easternmost in the Asian part of Turkey ( Kazanci 1984). The species is considered a typical Pontic faunistic element according to Haybach (1998, 2003).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |