
Arabella aracaensis, Steiner & Amaral, 2009
|
publication ID |
https://doi.org/ 10.1080/00222930903219988 |
|
persistent identifier |
https://treatment.plazi.org/id/625387E4-6C14-FFAB-9601-CB5C0852F970 |
|
treatment provided by |
Felipe |
|
scientific name |
Arabella aracaensis |
| status |
sp. nov. |
Arabella aracaensis View in CoL sp. n.
( Figures 4–6 View Figure 4 View Figure 5 View Figure 6 )
Material examined
Holotype: ZUEC-POL-103, Araçá Beach , São Sebastião Channel, State of São Paulo, Brazil, 45°24′28″W; 23°48′55″S, 3 December 2002, intertidal, mud, coll. Tatiana M. Steiner. GoogleMaps
Paratype series: ZUEC-POL-2176 (1 specimen), Araçá Beach , São Sebastião Channel, State of São Paulo, Brazil, 45°14–30′W; 23°41–53′S, 3 December 2002, intertidal, mud, coll. Tatiana M. Steiner ; ZUEC-POL-2175 (1 specimen), Araçá Beach , São Sebastião Channel, State of São Paulo, Brazil, 45°14–30′W; 23°41–53′S, 3 December 2002, intertidal, mud, coll. Tatiana M. Steiner ; ZUEC-POL-2176 (1 specimen), Araçá Beach , São Sebastião Channel, State of São Paulo, Brazil, 45°14–30′W; 23°41–53′S, 3 December 2002, intertidal, mud, coll. Tatiana M. Steiner ; ZUEC-POL- 2179 (1 specimen), Araçá Beach , São Sebastião Channel, State of São Paulo, Brazil, 45°14–30′W; 23°41–53′S, 21 January 1997, intertidal, coll. A. Cecília Z. Amaral ; ZUEC-POL-2178 (1 specimen), Araçá Beach , São Sebastião Channel, State of São Paulo, Brazil, 45°14–30′W; 23°41–53′S, 25 October 1996, intertidal, coll. A. Cecília Z. Amaral.
Material used for SEM: from ZUEC-POL-2178 (parapodia 40 to 47, 70 to 76). Additional material: 2 slides from ZUEC-POL-2178 (ZUEC-POL-2178A1 – parapodia 6, 25, 62, 80, 115, 146; ZUEC-POL-2178A2 – jaws); 2 slides from ZUEC-POL-2179 (ZUEC-POL-2179A1 – parapodia 7, 21, 30; ZUEC-POL-2179A2 – jaws). SEM: parapodia 2, 7 and 50.
Description
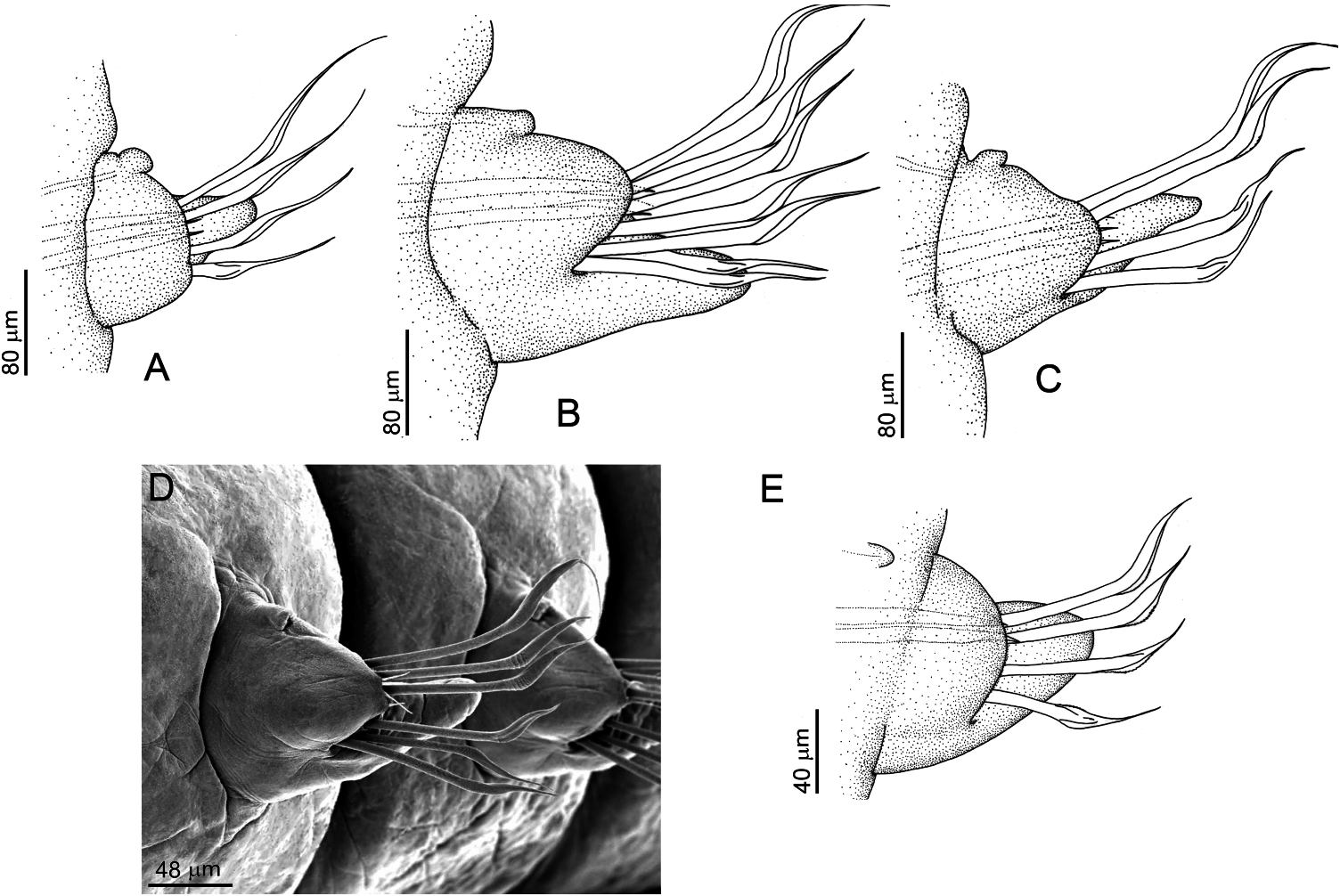
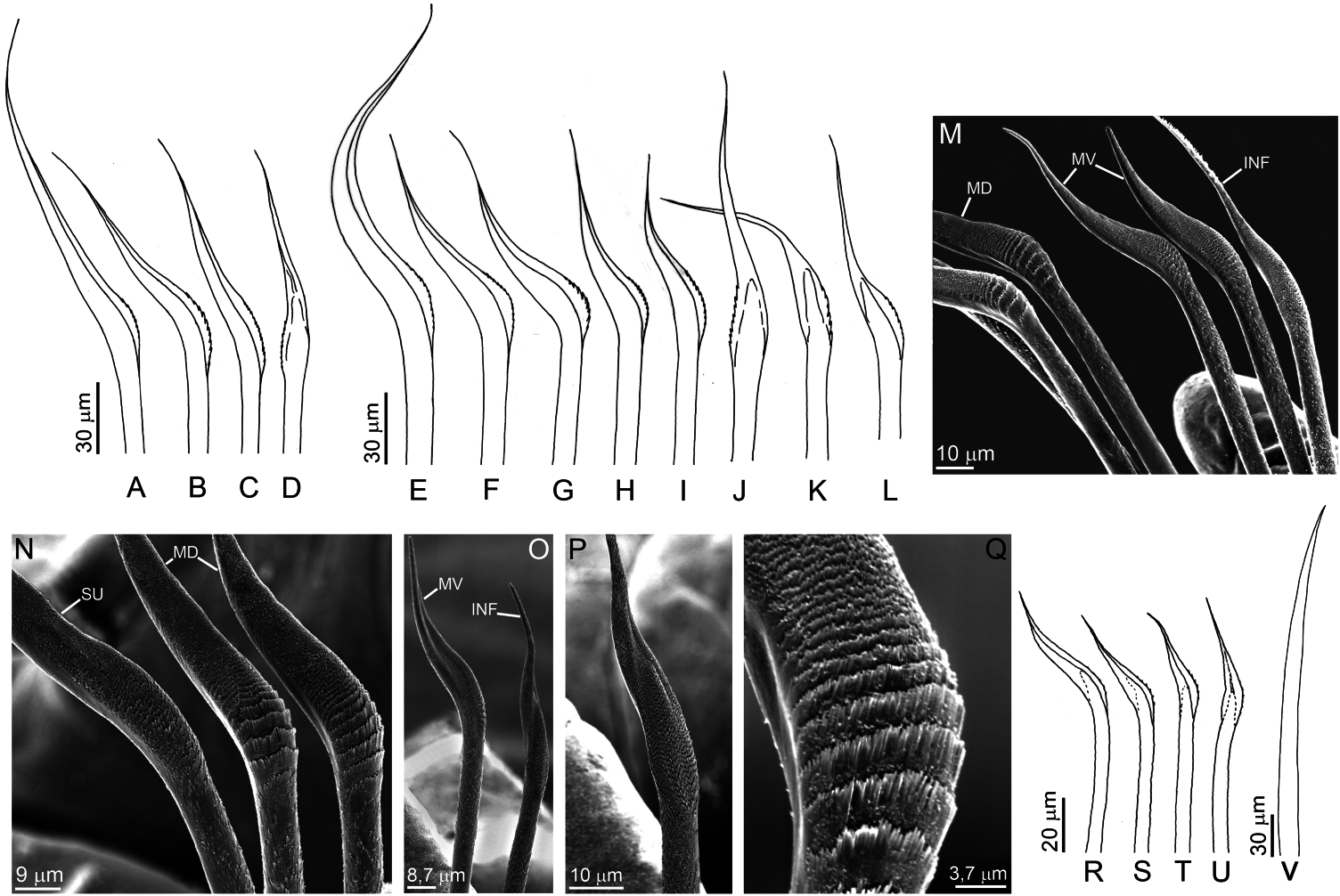
Holotype 23 mm long, 0.8 mm wide, 90 setigers, with one posterior fragment, with pygidium, 18 mm long, 80 setigers. Smallest specimen is incomplete, 5 mm long, 0.6 mm wide, 19 setigers (ZUEC-POL-ST2179). Largest specimen is incomplete, 17.5 mm long, 1 mm wide, 55 setigers (ZUEC-POL-2175). One almost complete specimen, 56 mm long, 0.8 mm wide, 166 setigers (ZUEC-POL-2178). Median and posterior segments with a small median brown band, more conspicuous dorsally. Width uniform along the body, narrowing near pygidial region. Prostomium conical, inflated, not compressed dorsoventrally ( Figure 4A–C View Figure 4 ), slightly wider than long, without pigmentation. Two pairs of eyes, arranged in line at base of prostomium; inner pair smaller and located superficially in the tissue. Peristomial rings slightly wider than base of prostomium, both equal in length and shorter than the following setigers. Mandibles joined by short ligament; distal plates with concentric growth rings ( Figure 4F View Figure 4 ). Paired maxillary support four times longer than MI ( Figure 4D View Figure 4 ); unpaired support wide at the base, tapering distally, two-thirds length of paired support. MI slightly asymmetrical ( Figure 4E View Figure 4 ), distally falcate, with six to eight basal teeth; left MII with seven to eight teeth, right MII longer than left one and with about ten teeth plus one additional tooth on anterior margin ( Figure 4D View Figure 4 , arrow); MIII and MIV with an additional broad dorsal chitinous plate ( Figure 4D View Figure 4 , cp); anterior tooth of MII to MIV longer; next tooth shorter than following ones, except left MII where teeth are same length. Anterior tooth of MII to MIV extends laterally, through a ridge ( Figure 4D View Figure 4 , rd) that ends in another tooth of same length ( Figure 4D View Figure 4 , at). First two to three parapodia smallest, next parapodia increasing in size gradually along anterior region, and maintaining size in median and posterior regions. First setigers with short, truncate pre-setal lobule ( Figure 5A View Figure 5 ); following setigers with rounded pre-setal lobule ( Figure 5B–D View Figure 5 ). Post-setal lobule always conical and twice as long as pre-setal. Last parapodia slightly shorter. Notopodium represented by small papilla with one to two short slender inner aciculae. Simple geniculate limbate setae with one serrated blade, divided into four groups, 1 and 2 supra-acicular, 3 and 4 subacicular: (1) 1 superior, longer than others, and blade finely serrated ( Figure 6A, E, N View Figure 6 ); (2) 1–2 mediodorsal ( Figure 6B, F, G, M, N View Figure 6 ); (3) 1–2 medioventral ( Figure 6C, H, I, M View Figure 6 ), shorter than mediodorsal and distal end more sloped. The slope is remarkable in the mediodorsal, which has a coarsely serrated blade ( Figure 6Q View Figure 6 ); (4) 1, rarely 2, modified ventral setae ( Figure 6D, J–L, M, O, P View Figure 6 ), with blade finely serrated, as wide as other setae of bundle. Setae of anterior parapodia slightly longer than rest of body, with narrower blades ( Figure 6A–D View Figure 6 ). Acicula amber-coloured ( Figure 6V View Figure 6 ), about three per parapodium, emerging slightly beyond the lobule. Narrow conical pygidium, as long as 2–3 last setigers ( Figure 4G View Figure 4 ); one pair of digitiform lateral pygidial cirri, as long as the pygidium.
Remarks
The shape of the prostomium ranged from triangular ( Figure 4A View Figure 4 ) to conical ( Figure 4B View Figure 4 ). The colour of eyes in fixed animals ranged from black to light brown.
The jaws analyzed belong to specimens ZUEC-POL-2178, ZUEC-POL-2175 and ZUEC-POL-2176, and the smallest specimen ZUEC-POL-2179. The general morphology of the jaws is the same in all specimens. The asymmetry of MI is related to the size, with the left MI slightly larger than the right MI, except in ZUEC-POL-2176, in which the inverse occurs. Similarly, all specimens have a large right MII, except in ZUEC-POL-2176 where left MII is larger than right MII. In ZUEC-POL-2178 the asymmetry of MI is not easily perceptible.
The anterior tooth of the large right MII, as well as the MIII, MIV and MV, is large, easily visible, and is a character observed in all specimens. The only maxilla that does not have this large tooth is the short left MII, where all the teeth have similar lengths.
In the small specimen ZUEC-POL-2179 some differences were observed. The demarcation between the two peristomial rings is less apparent dorsally. Both lobules are slightly larger and robust ( Figure 5E View Figure 5 ); the notopodial papilla has only one slender aciculum, and the neuropodium has one or two aciculae. The number of setae is about four per parapodium, and the blades have a short limb ( Figure 6R–U View Figure 6 ). The unpaired support of MI was not observed. The asymmetry of MI is evident ( Figure 4E View Figure 4 ). The shape and the pattern of dentition of MI, as well as the other maxillae, is basically the same, and the number of teeth was: MI = 7 + 6, MII = 9 + 6, MIII = 5 + 5, MIV = 4 + 3, MV = 1 + 1. Studies have shown that there is not necessarily a direct relationship between the number of teeth and the size of the maxilla: small maxillae can have the same number as large ones ( Kielan-Jaworowska 1966). The number of teeth can also vary according to the age of the animal.
Discussion
The character asymmetry of maxillae was considered as invariable, and the number of teeth as variable and size-dependent. The morphology of the jaws and the number of maxillae show only slight variation. Because of this, the new species was not compared with species that show wide variation in these characters, taking into account the size of the body. Therefore, the new species was compared with species that have asymmetrical maxillae as a constant character.
In general, most descriptions of species of Arabella do not describe in detail the morphology of the maxillae. Descriptions that mention the serrated aspect of the blades are also few. It is possible that the serrated blade is similar in all species of the genus, but this aspect is insufficiently studied. Perkins (1979) described three species of Cenothrix and mentioned the same pattern of serrated blades; the same occurs with A. semimaculata Moore, 1911 , A. asymmetrica Crossland, 1924 and A. protomutans Orensanz, 1990 .
Arabella aracaensis View in CoL sp. n. was compared with many descriptions of A. iricolor ( Fauvel 1923; Pettibone 1963; Orensanz 1974; Gardiner 1975; Lana 1984; George and Hartmann-Schroder 1985; Hilbig 1995; Núñez-Fraga 1990). Basically, this species is characterized by the presence of asymmetrical maxillae and the absence of a modified ventral seta. However, a revision of specimens referred to this species is necessary.
Other species lack the modified ventral seta: A. asymmetrica View in CoL , A. semimaculata View in CoL , A. pectinata Fauchald, 1970 View in CoL , A. cincta Hartmann-Schröder, 1962 View in CoL and A. protomutans View in CoL . Of these, A. protomutans View in CoL shows some similarities to A. aracaensis View in CoL sp. n., including the general morphology of the body, shape of the MI and paired maxillary support. The serrated blades show the same pattern as A. aracaensis View in CoL , except the ventral seta which is smooth. Furthermore, the left MI is bifid, a character that is absent in A. aracaensis View in CoL . Arabella logani Crossland, 1924 View in CoL shows similarities in the shape of the maxillae and the pattern of the serrated blades of setae. However, Colbath (1989) studied the type species, found a bifid left MI, and noted that this character distinguishes this species from A. iricolor , since both have the same type of ventral modified setae. If the MI of A. logani View in CoL is bifid, it is possible that A. protomutans View in CoL is the same species.
Of the other species that have modified ventral setae, none resembles A. aracaensis , either because they have a symmetrical apparatus ( A. atlantica Crossland, 1924 ; A. novecrinita Crossland, 1924 ) or have a ventral seta that is very robust and tapers abruptly ( A. maculosa Verrill, 1900 ; A. multidentata ; A. monroi Colbath, 1989 and A. panamensis Colbath, 1989 ).
According to Orensanz (1990), oenonids have no growth rings on the mandibles, and growth occurs by intussusception: in other words, by gradual transformation and incorporation of the formative elements of the mandibles; rather than by apposition, i.e., growth through adding layers, which would form rings. Because of the intense dark coloration and dull surface of the mandibles, the rings may pass unperceived. However, by the use of optical microscopy with incident light, it is possible verify growth rings clearly. Also, according to Colbath (1989), the maxillae are not mineralized, as in other Eunicida , and are strongly sclerotinized and contain slight traces of metals, which can interfere with observation of the rings.
Distribution
Atlantic Ocean: Brazil (State of São Paulo, São Sebastião Channel, Araçá Beach).
Etymology
The species was named Arabella aracaensis in defence of the preservation of Araçá Beach, the collection locality of the two species described here. This beach supports a highly diverse biota which is threatened with extinction because of the expansion of the Port of São Sebastião in the State of São Paulo.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Arabella aracaensis
| Steiner, Tatiana M. & Amaral, Antonia Cecília Z. 2009 |
Arabella aracaensis
| Steiner & Amaral 2009 |
A. aracaensis
| Steiner & Amaral 2009 |
A. aracaensis
| Steiner & Amaral 2009 |
A. aracaensis
| Steiner & Amaral 2009 |
A. protomutans
| Orensanz 1990 |
A. protomutans
| Orensanz 1990 |
A. protomutans
| Orensanz 1990 |
A. pectinata
| Fauchald 1970 |
A. iricolor
| , Kielan-Jaworowska 1966 |
A. iricolor
| , Kielan-Jaworowska 1966 |
A. cincta Hartmann-Schröder, 1962
| Hartmann-Schroder 1962 |
A. asymmetrica
| Crossland 1924 |
Arabella logani
| Crossland 1924 |
A. logani
| Crossland 1924 |
A. semimaculata
| Moore 1911 |