
Alderina canariensis, López-Fé, 2006
|
publication ID |
https://doi.org/ 10.1080/00222930601043763 |
|
persistent identifier |
https://treatment.plazi.org/id/2C138783-DC1A-550A-E3B8-FEB1FB7526FE |
|
treatment provided by |
Felipe |
|
scientific name |
Alderina canariensis |
| status |
sp. nov. |
Alderina canariensis sp. nov.
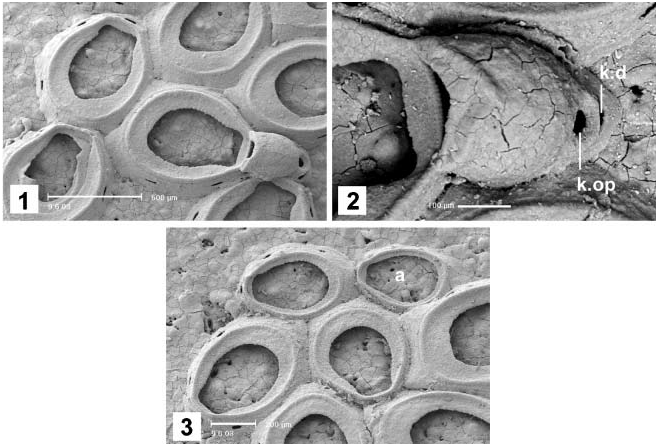
( Figures 1–3 View Figures 1–3 ; Table I)
Material
Many colonies on different pieces of Madrepora oculata .
Holotype: a complete colony with ancestrula and ovicells ( MNCN 25.03 About MNCN /3722).
Other material. Numerous colonies on ‘‘ Madrepora 1’’ (MNCN 25.03/3730) and ‘‘ Madrepora 2’’ (MNCN 25.03/3736).
Description
Colony encrusting, unilaminar. Autozooids oval, with well-developed gymnocyst and cryptocyst. Spines absent except in the ancestrula and first zooids. Opesia extensive, occupying approximately two-thirds of the zooidal length. Ovicell hyperstomial, slightly longer than wide, not closed by the zooidal operculum. The ectooecium has usually a small triangular uncalcified area, but some ovicells have the ectooecium fully calcified. The ovicell is supported by a kenozooid which lies below and has a small gymnocyst and opesia and its own communication organs. Interzooidal communication by basal pore chambers, six per autozooid. Avicularia absent. The ancestrula is similar to an autozooid, but has thinner gymnocyst and cryptocyst and bears 10 marginal spines. The number of spines decreases progressively through early astogeny.
Discussion
The features of the new species, such as the gymnocyst and cryptocyst, the hyperstomial ovicell and the basal dietellae, are clearly calloporid. Among the calloporid genera, Alderina Norman, 1903 is the most appropriate to accommodate A. canariensis sp. nov., because its type species, A. imbellis (Hincks) , has similar autozooids, lacks avicularia and its ovicell is not closed by the operculum. However, some differences exist between A. canariensis and A. imbellis , regarding mainly the marginal spines and the ovicell. In contrast with A. canariensis , A. imbellis is completely devoid of spines, even in the ancestrula ( Hayward and Ryland 1998). The ovicell of A. canariensis sp. nov. has a small proximal area of uncalcified ectooecium, always clearly smaller than the same area of A. imbellis , and in some ovicells the ectooecium is fully calcified; these states coexist in the same colony. Alderina canariensis sp. nov. could also be near to Crassimarginatella , but the ovicell of this genus is closed by the operculum.
The most remarkable difference between A. canariensis sp. nov. and most calloporids (including A. imbellis ) is the ovicell supported by a kenozooid, which has a small opesia ( Figure 2 View Figures 1–3 ). This is the situation termed B type by Bishop and Househam (1987). Although initially proposed for species of the genus Puellina , the classification of hyperstomial ovicells as types A, B and C depending on the degree of modification of the succeeding zooid may be used for many cheilostomes. The B type corresponds to the intermediate degree, in which the succeeding zooid is reduced to a kenozooid but still has a part of the frontal wall. It occurs in some cheilostome genera, such as Puellina and Macropora ( Bishop and Househam 1987; López de la Cuadra and García Gómez 1997), where the three types are present depending on the species. Then, the B type ovicell of A. canariensis sp. nov. may be considered a specific character, which does not preclude the inclusion of this species in Alderina .
Within the Calloporoidea, the ovicell of type B also occurs in two austral uniserial species: Daisyella libita Gordon, 1989 and Pyriporoides judyae Branch and Hayward, 2005 . Nevertheless, the ovicell of both species lack the small proximal area of uncalcified ectooecium which occurs in Alderina . Further, Daisyella libita has a median longitudinal suture ( Gordon 1989) and Pyriporoides judiae has a median longitudinal ridge ( Branch and Hayward 2005), both structures lacking in any species of Alderina . An uniserial species, Daisyella bathyalis Rosso and Taylor, 2002 , has been assigned to Daisyella in the northern Atlantic ( Rosso and Taylor 2002). The ovicell of D. bathyalis has a proximal area of uncalcified ectooecium which occurs not only in Alderina , but also in some species of other calloporid genera like Callopora , Amphiblestrum , or Copidozoum , but it lacks any longitudinal structure, suture, or ridge, which are respectively present in Daisyella and Pyriporoides . Perhaps the generic assignation of Daisyella bathyalis could be reconsidered, but this is beyond the scope of the present work.
It is thus apparent that ovicells of B and C types, associated with a kenozooid, occur independently in different cheilostome genera, perhaps by reduction of the succeeding zooid.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.