
Afroedura pienaari, Jacobsen, Niels H. G., Kuhn, Arianna L., Jackman, Todd R. & Bauer, Aaron M., 2014
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3846.4.1 |
|
publication LSID |
lsid:zoobank.org:pub:0DD5A603-D65F-4976-BBE9-94DA7110053F |
|
DOI |
https://doi.org/10.5281/zenodo.5620601 |
|
persistent identifier |
https://treatment.plazi.org/id/744387D5-B12B-A105-EBAD-FC50E1BBFA3C |
|
treatment provided by |
Plazi |
|
scientific name |
Afroedura pienaari |
| status |
sp. nov. |
Afroedura pienaari sp. nov.
( Figs. 11 View FIGURE 11 B–C)
Afroedura pondolia subsp. nov. Pienaar et al. 1983
Afroedura langi 'Waterpoort' Jacobsen 1992a, 1997; Kirchhof et al. 2010 Afroedura langi 'Tshipise' Jacobsen 1992a, 1997
Afroedura langi 'Shinokwen' Jacobsen 1992a, 1997
Holotype. TM 81144, adult male, Farm Bristol 760MS, 22°59' S, 29°37' E, Soutpansberg District (2229DC), Limpopo Province, Republic of South Africa, collector R. E. Newbery, 25 July 1981.
Paratypes. TM 81133–81139, 81141–81143, 81145–81146, 81160–81165, same data as holotype; TM 81140, same locality as holotype, collector N. H. G. Jacobsen, 2 February 1979; TM 81175, Farm Crimea 747MS, Soutpansberg District, Limpopo Province (2229DC), collector N. H. G. Jacobsen, 1 February 1979; TM 81147–81153, Farm Robertson 748MS, Soutpansberg District, Limpopo Province (2229DC), collector R. E. Newbery, 24 July 1981; TM 81167–81169, Farm Waterpoort 695MS, Soutpansberg District, Limpopo Province (2229DC), collector R. E. Newbery, 24 July 1981; TM 81170–81173, Farm The Moss 763MS, Soutpansberg District, Limpopo Province (2229DC), collector R. E. Newbery, 7 November 1985.
Additional material examined (all from Limpopo Province). TM 81154–81157, 81174, Farm Zoutpan 459MS, Soutpansberg District (2229CD); TM 81107, Musekwas location 194MT, Dzanani District (2230CC); TM 81103–81104, 81106, 81111 Gumela, Sibasa District (2230CB) TM 81102, 81105, 81110, Lavhalisa, Dzanani District (2230CA); TM 30414, 35 km southwest of Tshipise, Soutpansberg District (2229DD); TM 81342, Vhuswinzhe, Dzanani District (2230CC); TM 81101, 81108–81109, near Shinokwenfontein, KNP (2231AC); TM 77353–77355, Bobomeni, KNP (2231BC); TM 77356–77360, Mashipange, KNP (2230DB); TM 77336–77345, 3 km W of Baobab Hill, KNP (2231AC); TM 77329–77335 Pafuri border gate, KNP (2231AC); TM 77346–77352, 5 km NW of Hutwini, KNP (2231AC); TM 77361–77362, 77363–77367, Madziringwe stream, S of the Trails Camp, KNP (2231CA); TM 77368–77372 Nyala Drive, 4 km W of Bobomeni, KNP (2231AC); TM 77309–77304, Shipudza Spring, KNP (2231CA); TM 77315, 77318–77323, Shidzivani, KNP (2231CA); TM 77324–77328, Xantangalani, KNP (2231CA); TM 47314 Vivo area, Soutpansberg District (2329AB); TM 81129–81132 Farm Calitzdorp 221LS, Soutpansberg District (2329BA).
Etymology. The species is named for the late Dr. U. de V. Pienaar (1930–2011), former Chief Warden of Kruger National Park and former Chief Director of South African National Parks, in recognition of his work in furthering herpetofaunal knowledge of the Kruger National Park.
Diagnosis. A medium-sized Afroedura (maximum SVL 53 mm) differing from all other congeners by the following combination of characters: two pairs of enlarged subdigital lamellae per digit; tail faintly verticillate and flattened near base, with four subcaudal rows and 6–7 supracaudal rows per verticil; dorsal scales smooth, 86–111 scale rows at midbody; internasal scales absent; 13–19 precloacal pores in males.
Description. (based on holotype TM 81144) Adult male; SVL 48.0 mm; TailL 57.0 mm; mass before preservation 2.2 g. A medium sized, dorsoventrally depressed Afroedura , with a flat, ovate head, wider than the neck. Rostral 2.5 times wider than high; nostrils pierced between rostral, first upper labial and three nasal scales; nasorostrals raised and in broad contact behind rostral. Scales on snout variable, large and flattened, decreasing in size posteriorly, to being smallest on the crown of the head; 10 scales between nasals and eye and 16 between eye and ear; four supraciliary spines above eye; supralabials nine. Mental wedge-shaped, longer than wide; one postmental. Infralabials seven.
Dorsal scales uniform, rounded, juxtaposed and flattened paravertebrally, becoming larger and oblique laterally. Midbody scales in 93 rows. Ventral scales flat, smooth, imbricate and irregular to hexagonal. Limbs robust; two pairs of enlarged scansors under each digit and four inferomedian scales under 4th toe. Precloacal pores 18, arranged in a shallow ‘V’-shape, with a single median poreless scale interrupting the series. Tail faintly verticllate, widening slightly posterior to the cloaca before tapering. Caudal verticils with seven dorsal and four ventral scale rows. Supracaudals almost square to slightly longer than broad, with a rounded to apical posterior margin; subcaudals almost square with a rounded posterior margin and imbricate; three post cloacal spurs at base of tail.
Color. Gray-brown to brown dorsally with 6–7 irregular dark brown to blackish crossbars occasionally bordered posteriorly by a black edge, followed by a white marking, the latter sometimes evident only as a median spot. Dorsum variegated and mottled with brownish black and off-white. Head mottled dorsally; a dark stripe extending from the nostrils through the eye to the rear of the head, linking to the second crossbar in the neck region. Limbs brownish with pale and dark spotting and marbling. Original tail with 8–13 crossbars, with interstitial areas spotted or marbled. Venter whitish to pinkish mesially.
Variation. There is significant variation in specimens of this species in several features ( Tables 4, 5), in part reflecting the very large sample size and its relatively large geographic range. A single internasal is present in TM 81142. 10–14 scales between nasals and eye and 16–22 between eye and ear, 2–4 supraciliary spines. Supralabials 8–12. One or two postmental scales present. Infralabials 7–9. Midbody scale rows 86–98 in type series, up to 111 in other specimens examined. 3–7 enlarged inferomedian scales under 4th toe. Precloacal pores in male paratypes in continuous or interrupted series of 14–18 (12 to 19 in other specimens examined, although 96% of all male specimens have at least 14 pores); no precloacal pores in females. 0–4 postcloacal spurs on either side of tail base. Original tails 50.0–58.4% of total length. 32.2% of specimens have regenerated tails (n=31).
Distribution. Recorded from along the northern Soutpansberg and into the northwestern Kruger National Park, Limpopo Province ( Fig. 6 View FIGURE 6 ). Kirchhof et al. (2010) reported this species from Lajuma, on the southwestern Soutpansberg, in an area that would be expected to harbour A. broadleyi , however, their specimens had the typical precloacal pore counts of A. pienaari and on this basis we accept their identification.
Natural history. Afroedura pienaari sp. nov. is a nocturnal, rupicolous gecko that inhabits crevices and fissures in and between boulders on rocky outcrops and under flakes of exfoliating rock beneath overhangs. It appears to be mostly solitary, but is occasionally found in pairs and rarely three indivduals may occupy the same crevice. Found on north and south-facing slopes in Makhuleke Sandy Bushveld (SVl 1), Limpopo Ridge Bushveld (SVmp 2), Soutpansberg Mountain Bushveld (SVcb 21) and Soutpansberg Summit Sourveld (GM 28) ( Mucina & Rutherford 2006) at elevations of 800–1200 m a.s.l. Kirchhof et al. (2010) reported this species from cracks and fissures in vertical rock walls and from caves and found several individuals in an electrical junction box.
Two soft shelled eggs are laid at a time. These are placed on the undersides of rocks, to which they adhere and then harden. Communal nesting takes place in favorable crevices.
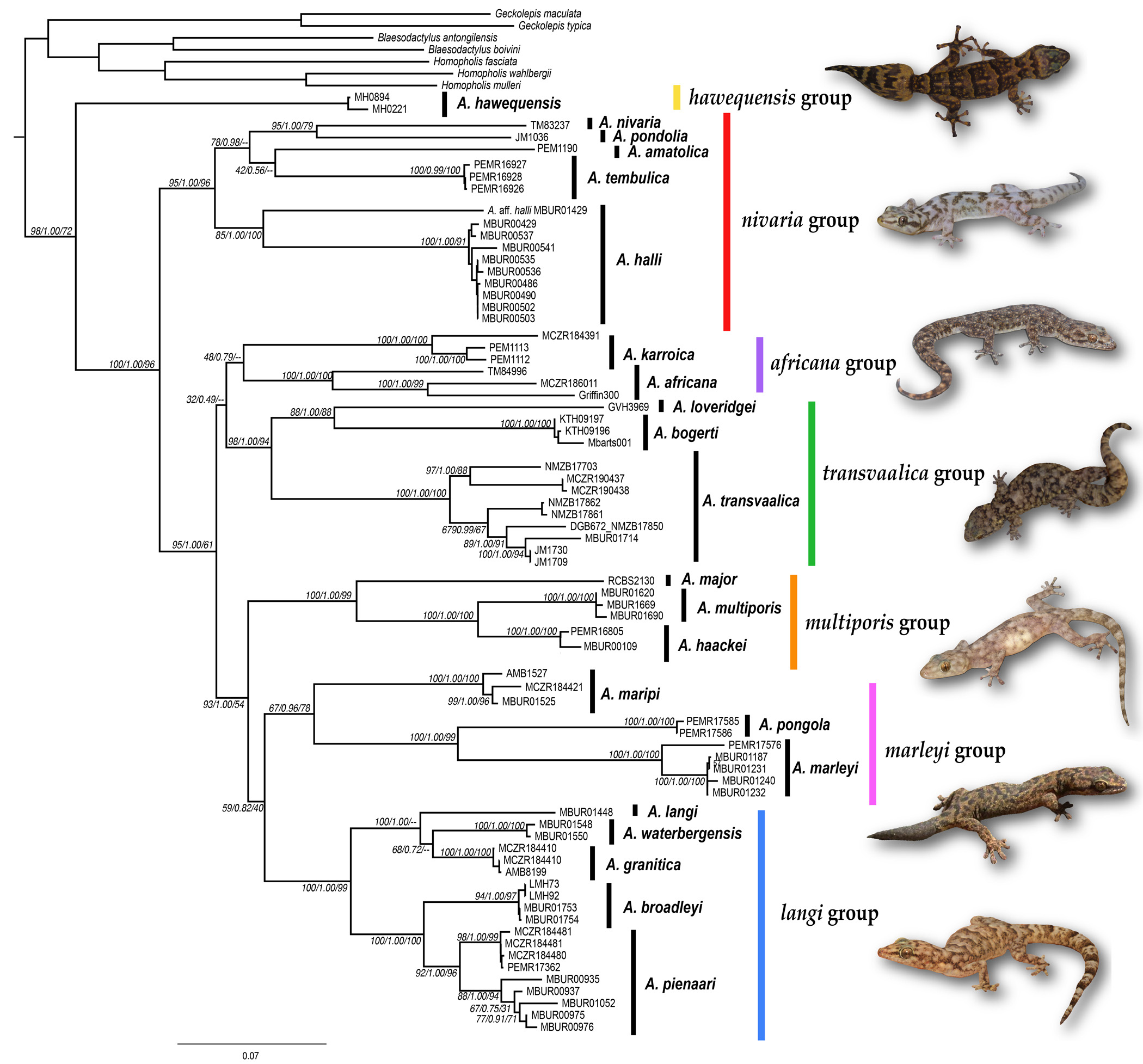
Remarks. Jacobsen (1990, 1992a) initially considered each of three discrete populations of this gecko to represent putative species. The western and eastern forms are similar in size but are separated by smaller geckos, with generally lower scale counts, in the central part of the range ( Table 5). Genetic divergences between populations of Afroedura pienaari sp. nov. are also pronounced ( Fig. 1 View FIGURE 1 ). However, further sampling may reveal continuity between populations and existing populational differences in morphological features are largely overlapping, so we here treat all of the northern Soutpansberg/Limpopo Valley A. langi complex geckos as a single species.
Waterpoort Tshipise KNP
Midbody scales n = 31 n = 9 n = 66
86–107 86–102 86–111 93.9 + 4.71 90.67 + 4.8 96.44 + 5.29
Eye–Ear scales n = 33 n = 9 n = 62
13–19 15–17 13–19
16.58 + 1.15 15.78 + 0.67 16.05 + 1.45
Supralabials n = 38 n = 10 n = 59
8–12 8–10 8–11
9.6 + 0.8 9.4 + 0.7 9.6 + 0.81
Infralabials n = 42 n = 10 n = 62
7–10 7–9 6–10
8.21 + 0.68 7.7 + 0.67 8.02 + 0.78
Caudal verticils (supracaudals/subcaudals) 7/4–5 6–8/4 6–7/4
Precloacal Pores n = 16 n = 7 n = 27
14–19 12–15 14–19
16.94 + 1.53 14.14 + 1.21 16.89 + 1.40
Adult male SVL (mm) n = 16 n = 5 n = 27
51.0 42.5 48.0
44.56 + 3.08 39.60 + 2.16 43.96 + 2.61
Adult female SVL (mm) n = 16 n = 2 n = 31
52.0 39.0 53.0
47.12 + 3.72 34.5 + 6.36 44.47 + 4.76 This species may be distinguished from the geographically proximate A. broadleyi sp. nov. and from A. waterbergensis sp. nov. by its greater number of precloacal pores and from A. leoloensis sp. nov. by its lower number of pores. In addition, it is substantially larger than A. leoloensis sp. nov., A. langi , and A. waterbergensis sp. nov. (see Table 4) and has a significantly greater number of supralabials than A. langi (P <0.001, t134 d.f.) and more scales from the eye to ear than A. broadleyi sp. nov. and A. langi (p <0.001, t172 d.f. and p <0.001, t127 d.f., respectively). This species has largely overlapping scale counts with A. granitica sp. nov., which is larger (58 versus 53 mm maximum SVL and even greater mean differences; Table 4). The great majority of male specimens, however, have 14 or more precloacal pores (up to 19), averaging more than 16, whereas A. granitica sp. nov. has a mean of only 14 pores. Afroedura pienaari sp. nov. also has fewer eye to ear scales than A. granitica sp. nov., although not statistically so.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.