
Aegla buenoi Marçal & Teixeira, 2021
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5005.3.4 |
|
publication LSID |
lsid:zoobank.org:pub:A7FB5098-295C-496E-84FB-82CD3C524AB1 |
|
DOI |
https://doi.org/10.5281/zenodo.5152333 |
|
persistent identifier |
https://treatment.plazi.org/id/F032E974-6EDE-4BA5-8577-17207631A62A |
|
taxon LSID |
lsid:zoobank.org:act:F032E974-6EDE-4BA5-8577-17207631A62A |
|
treatment provided by |
Plazi |
|
scientific name |
Aegla buenoi Marçal & Teixeira |
| status |
sp. nov. |
Aegla buenoi Marçal & Teixeira View in CoL n. sp.
( Figs 3–5 View FIGURE 3 View FIGURE 4 View FIGURE 5 )
urn:lsid:zoobank.org:act:F032E974-6EDE-4BA5-8577-17207631A62
Type-material. Holotype: male, Brazil, Paraná, Ibaiti, Paranapanema River basin, Cinzas River sub-basin, Carneiro stream, 23°57′19.71″S, 50°14′16.09″W, altitude 656 m, C.A. da Silva, R.H.C. do Nascimento and R. Rockembacher coll., 17.v.2016 ( MZUEL 249 ) GoogleMaps . Paratypes: 1 male and 2 females, same data as holotype ( MZUEL 245 ) GoogleMaps . 3 males and 2 females, Brazil, Paraná, Ibaiti, Paranapanema River basin, Cinzas River sub-basin, tributary of the Carneiro stream, 23°57′27.00″S, 50°14′5.00″W, altitude 620 m, G.M. Teixeira and J.J.S. Rosa coll., 12.vi.2017 ( MZUEL 246 , genetic vouchers: BOLD access AEGPR016-18 , AEGPR017-18 , AEGPR018-18 , AEGPR019-18 , AEGPR020-18 ) GoogleMaps .
Type-locality. Carneiro stream, city of Ibaiti, Paraná state, Brazil.
Geographical distribution. The new species has been collected in two streams within the Cinzas River subbasin, Paranapanema River basin. Both streams are located in the city of Ibaiti, Paraná state, Brazil.
Diagnosis. Rostrum triangular, narrow base, reaching distal apex of compound eyes, carinate. Subrostral process well developed, occupying proximal half of subrostral margin, anterior and posterior margins forming obtuse angle (122°). Extra-orbital sinus deep. Anterolateral spines not reaching basal margin of cornea. Epigastric prominences pronounced and oval. Protogastric lobes pronounced. Cervical groove U-shaped. Cardiac area subrectangular. Proximal dorsal margin of dactylus with rudimentary or absent lobe. Palmar crest disciform, outer surface excavated. Subterminal lobe of carpus well defined, pointed. Ventromesial border of cheliped ischium with 4 or 5 tubercles. Anteromesial region of third thoracic sternite tapered. Anterior angle of second abdominal epimeron and ventral angles of third and fourth abdominal epimeron with corneous scale apically. Anterior margin of second abdominal epimeron almost straight. Uropods wide.
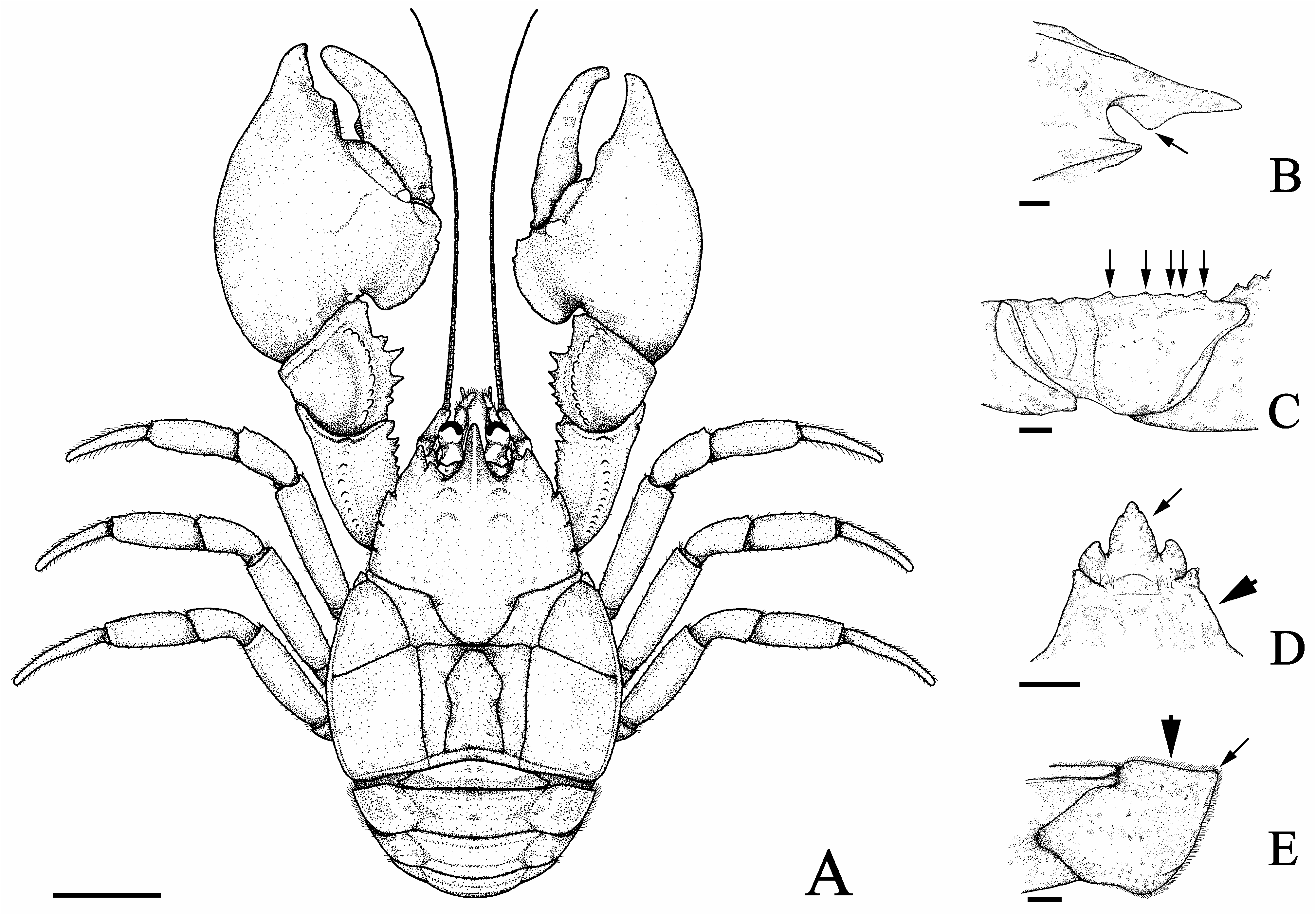
Description of holotype. Carapace moderately convex, gastric region convex, dorsal surface scabrous, covered with punctations, anterior and posterior branchial areas expanded laterally (CW/CLE = 1.02) ( Figs 3A View FIGURE 3 , 4A View FIGURE 4 ). Rostrum triangular, narrow base (RBW/LMR = 0.98), reaching distal apex of compound eyes, carinate along entire length, small corneous scales on lateral margins, tip apparently broken; ventral portion of rostrum much higher than dorsal in profile. Rostral carina beginning at level of protogastric lobes, with row of corneous scales extending next to apex. Subrostral process well developed and covered by small setae, occupying proximal half of subrostral margin, tip rounded, anterior and posterior margins forming obtuse angle (122°) ( Figs 3B View FIGURE 3 , 4B View FIGURE 4 ). Eyestalk and cornea well developed. Orbital and extra-orbital sinuses deep. Orbital sinus U-shaped. Orbital spines well developed, with small terminal corneous scale. Anterolateral spines acuminate apically with small corneous scales terminally, not reaching basal margin of cornea. Epigastric prominences pronounced and oval, with corneous scales. Protogastric lobes pronounced, with corneous scales ( Fig. 4B View FIGURE 4 ). Gastric area elevated in relation to hepatic lobes and rostrum in lateral view. Demarcation between hepatic lobes well defined. Lateral margins of hepatic lobes with small corneous scales. Cervical groove U-shaped. Transverse dorsal linea sinuous along its extension. Areola rectangular (AH/ [(APM+AAD)/2] = 2.54). Cardiac area subrectangular (TDL/PMC = 1.22). Epibranchial area strongly elongated, anterolateral angle with 2 corneous scales, lateral margin with row of small corneous scales and small setae. Lateral margins of anterior and posterior branchial areas with row of corneous scales and small setae.
Chelipeds unequal, left largest.
Major cheliped (left). Dactylus ( Fig. 4C View FIGURE 4 ): dorsal margin and outer surface with small corneous scales, inner surface with setal tufts and scales; dorsal margin without proximal lobe; cutting margin with well-developed lobular basal tooth, followed by row of corneous scales up to distal end; row of small tufts of long setae next to cutting margin; pre-dactylar lobe well developed, rounded, smooth, without corneous scales. Propodus ( Fig. 4C View FIGURE 4 ): outer surface granular, globose aspect; palmar crest disciform with outer surface excavated, margin poorly serrated, covered by corneous scales; fixed finger cutting margin with well-developed lobular basal tooth, followed by row of corneous scales up to distal end; inner and outer surfaces of fixed finger with rows of long setae tufts next to cutting margin. Carpus ( Fig. 4C View FIGURE 4 ): dorsal margin with 2 proximal tubercles, 2 median spines, distal tubercle, double-tipped, each tubercle or spine with terminal corneous scale, subterminal lobe well defined, pointed, with small corneous scales, setae apically; inner surface with 2 distinct tubercles with acute corneous scale terminally; outer surface with carpal ridge high, with small corneous scales. Merus ( Fig. 4D View FIGURE 4 ): dorsolateral edge with distal tubercle, with corneous scale terminally, followed by row of tubercles decreasing in size proximally; ventromesial edge with 2 distal spines, with corneous scale terminally, followed by 3 tubercles of similar size, with corneous scale; ventrolateral border with 2 distal tubercles, with terminal corneous scale, followed by several small tubercles proximally. Ischium ( Figs 3C View FIGURE 3 , 4D View FIGURE 4 ): dorsolateral edge with spine, with corneous scale terminally; ventromesial border with large proximal tubercle, 3 small median tubercles and large distal tubercle, each with terminal corneous scale; ventrolateral border smooth.
Minor cheliped (right) similar to major cheliped except as noted hereafter. Dactylus ( Fig. 4E View FIGURE 4 ): cutting margin with rudimentary lobular basal tooth. Propodus ( Fig. 4E View FIGURE 4 ): cutting margin with rudimentary lobular basal tooth. Carpus ( Fig. 4E View FIGURE 4 ): dorsal margin with 2 proximal tubercles, 2 median spines, distal spine, each with terminal corneous scale; inner surface with 5 tubercles, corneous scale terminally. Merus ( Fig. 4F View FIGURE 4 ): ventromesial edge with distal spine, followed by 5 tubercles, each with terminal corneous scale. Ischium ( Fig. 4F View FIGURE 4 ): ventromesial border with proximal tubercle, distal tubercle, each with terminal corneous scale.
Second, third, and fourth pereiopods similar. Dactyli, propodi, carpi, meri and ischii with several rows of setal tufts and small scales on surface. Carpi and meri with row of tubercles with terminal corneous scale along dorsal margin. Meri and ischii with long setae concentrated along dorsal margin.
Anteromesial region of third thoracic sternite tapered, projecting between coxae of third maxillipeds, with scattered setae. Fourth thoracic sternite with anterolateral angles produced anteriorly, with scattered setae ( Fig. 3D View FIGURE 3 ).
Anterolateral angle of second abdominal epimeron and ventral angles of third, fourth abdominal epimeron well defined, with corneous scale apically. Anterior margin of second abdominal epimeron almost straight ( Fig. 3E View FIGURE 3 ). Uropods well developed, wide (WU/HWT = 1.04). Pleopods 2–5 absent.
Telson divided by longitudinal suture. Anterolateral and posterolateral margins well differentiated.
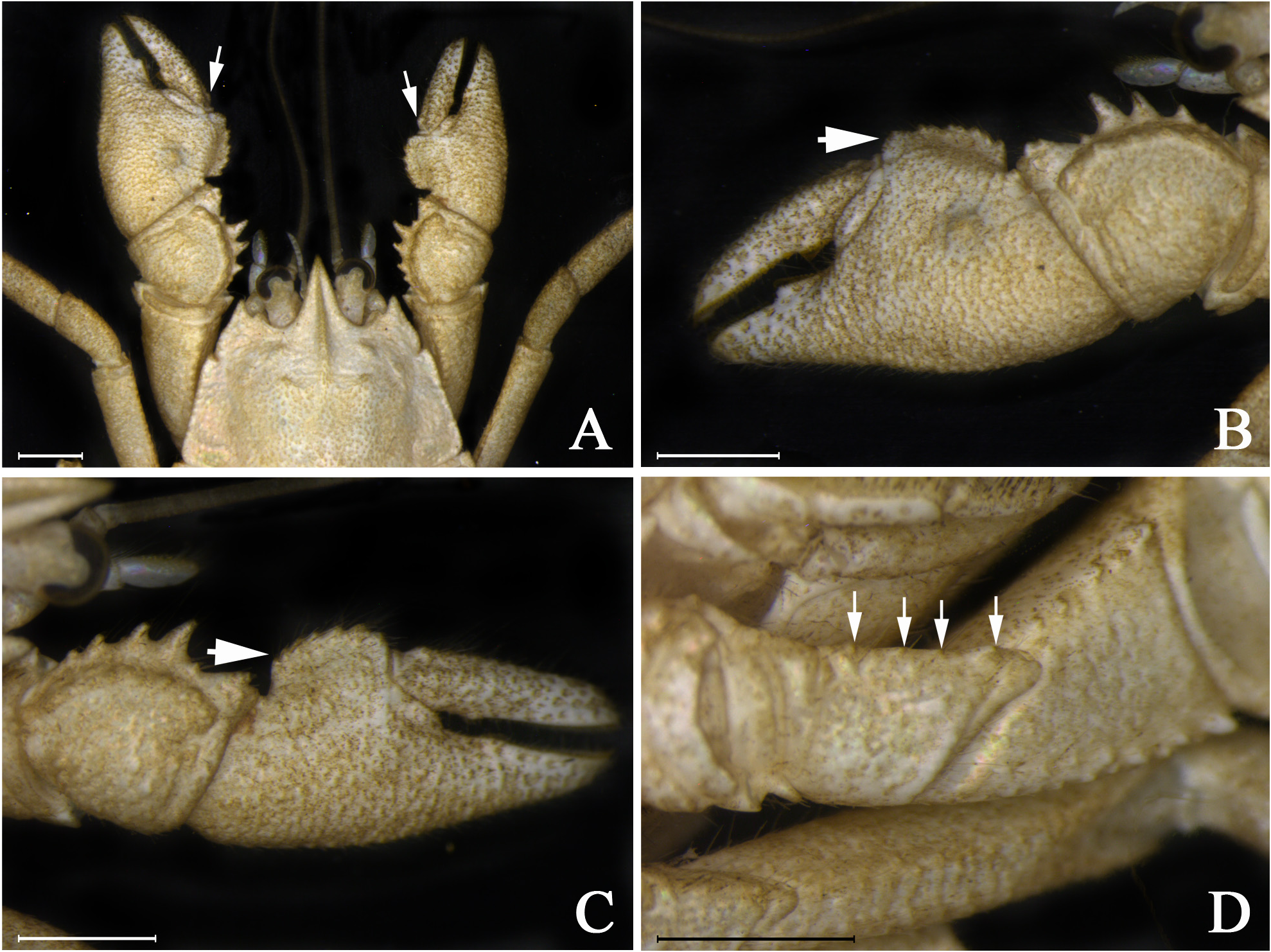
Variations. The rostrum is generally narrow base, but in some paratypes the base is wider than usual (RBW/ LMR = 1.03 ± 0.04; n = 5). In some specimens, the anterolateral spines can reach the basal margin of the cornea. The areola can be subrectangular (AH/[(APM+AAD)/2] = 2.12 ± 0.20; n = 4), trapezoidal (APM/AAD = 1.79 ± 0.10; n = 3), or rectangular (AH/[(APM+AAD)/2] = 2.31; n = 1). The cardiac area varies from subrectangular (TDL/PMC = 1.28 ± 0.06; n = 7) to trapezoidal (TDL/PMC = 1.45; n = 1). The proximal lobe on the dorsal margin of the dactylus of the major and minor chela may be rudimentary (as opposed to absent) ( Fig. 5A View FIGURE 5 ). The palmar crest is rectangular instead of disciform in small individuals (CLE <12 mm) ( Figs 5B–C View FIGURE 5 ). The ventromesial border of ischium may present four small tubercles instead of five ( Fig. 5D View FIGURE 5 ). The anterolateral angle of second abdominal epimeron may be unarmed instead of present a corneous scale as in the holotype. The uropods are narrow (WU/HWT = 0.92 ± 0.03; n = 3) in some specimens. All measurements taken on type-series and morphometric relationships are summarized in Table 1.
Biology. Unknown.
Etymology. The specific epithet is given in honor of Dr. Sérgio Luiz de Siqueira Bueno, for his outstanding contributions to the taxonomy and knowledge of the biology of aeglids.
Molecular data. Sequences of COI generated in this study were deposited in the BOLD database (access numbers AEGPR016-18, AEGPR017-18, AEGPR018-18, AEGPR019-18, AEGPR020-18).Altogether we analyzed 884 base pairs (bp) of COI with no insertions, deletions, or stop-codon. The mean base frequencies were: A = 0.2880, C = 0.1444, G = 0.1537, T = 0.4140.
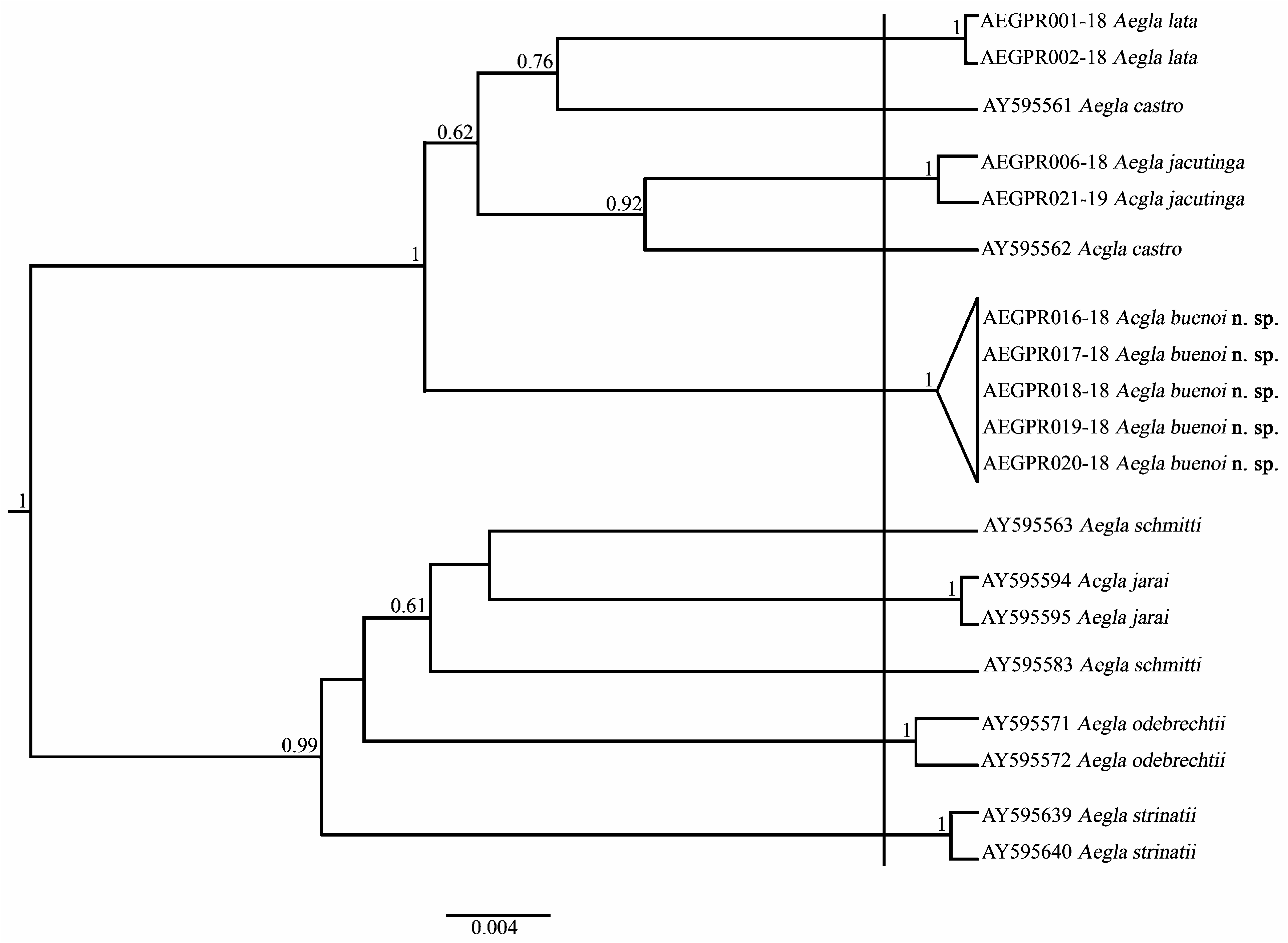
The genetic distance between A. buenoi n. sp. and its congeners ranged from 2.1% to 4.8% ( Table 2). Aegla jacutinga showed the lowest divergence relative to A. buenoi n. sp., and the other species. In contrast, A. strinatii was found to be most distantly related to the new species and exhibited higher interspecific variations than the others, with a minimum K2P divergence of 3.8%. Most of the species showed intraspecific distances lower than 0.4%. Aegla castro and A. schmitti , however, presented intraspecific variations higher than 2.6% ( Table 2).
Regarding species delimitation, the GMYC analysis indicated the presence of 10 independent strains ( Fig. 6 View FIGURE 6 ). Two distinct clusters, with various well-supported subclusters, were obtained. Interestingly, the sequences of A. castro and A. schmitti formed non-monophyletic groups. On the other hand, all sequences of A. buenoi n. sp. were joined in the same cluster, indicating the presence of a single species.

| R |
Departamento de Geologia, Universidad de Chile |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |