
Boroecia borealis ( G.O. Sars, 1866 )
|
publication ID |
https://doi.org/10.11646/zootaxa.4394.3.1 |
|
publication LSID |
lsid:zoobank.org:pub:9358DA65-1111-4702-A898-52519D53B181 |
|
DOI |
https://doi.org/10.5281/zenodo.6490271 |
|
persistent identifier |
https://treatment.plazi.org/id/D723883B-FFB7-FFE5-34BC-FB67FC69BF18 |
|
treatment provided by |
Plazi |
|
scientific name |
Boroecia borealis ( G.O. Sars, 1866 ) |
| status |
|
2.1. Boroecia borealis ( G.O. Sars, 1866) View in CoL
( Figures 1–3 View FIGURE 1 View FIGURE 2 View FIGURE 3 )
Synonymy:
1866 Conchoecia borealis Sars : 119.
1896 Conchoecia borealis— Brady & Norman: 685-6, Pl. 61, 1–19. 1906 Conchoecia borealis— Vavra: 48, Pl. 4, 64.
1928 Conchoecia borealis— Sars: 22, Pl. XI–XII.
1980 Conchoecia borealis— Deevey & Brooks: 114, Fig. 36 A–L, 37 A–G. 1973 Boroecia borealis —Poulsen: 166–169, Fig. 84 A– I. 1981 Boroecia borealis— Angel: 543–61.
2008 Boroecia borealis— Bashmanov & Chavtur: 341–347. 1994 Boroecia borealis —Richter: 61–65.
1995 Boroecia borealis —Chen & Lin: 124, Fig. 146, 1–7. 2008 Boroecia borealis— Angel et al.
2015 Boroecia borealis— Chavtur et al.: 168–85, Fig. 12 View FIGURE 12 –20.
The species was originally described by G. O. Sars (1866) from material collected in the vicinity of 68°N, 12°E off the Lofoten Islands. We nominate this approximate position as the type locality for the species. However, in the absence of good material from locations close to this type locality, we cannot designate any syntypes.
Male. Dimensions of the carapace and the lengths relative to the carapace of the principal setae on the first and second antennae are shown in Table 3 and compared to the characteristics of the other species.
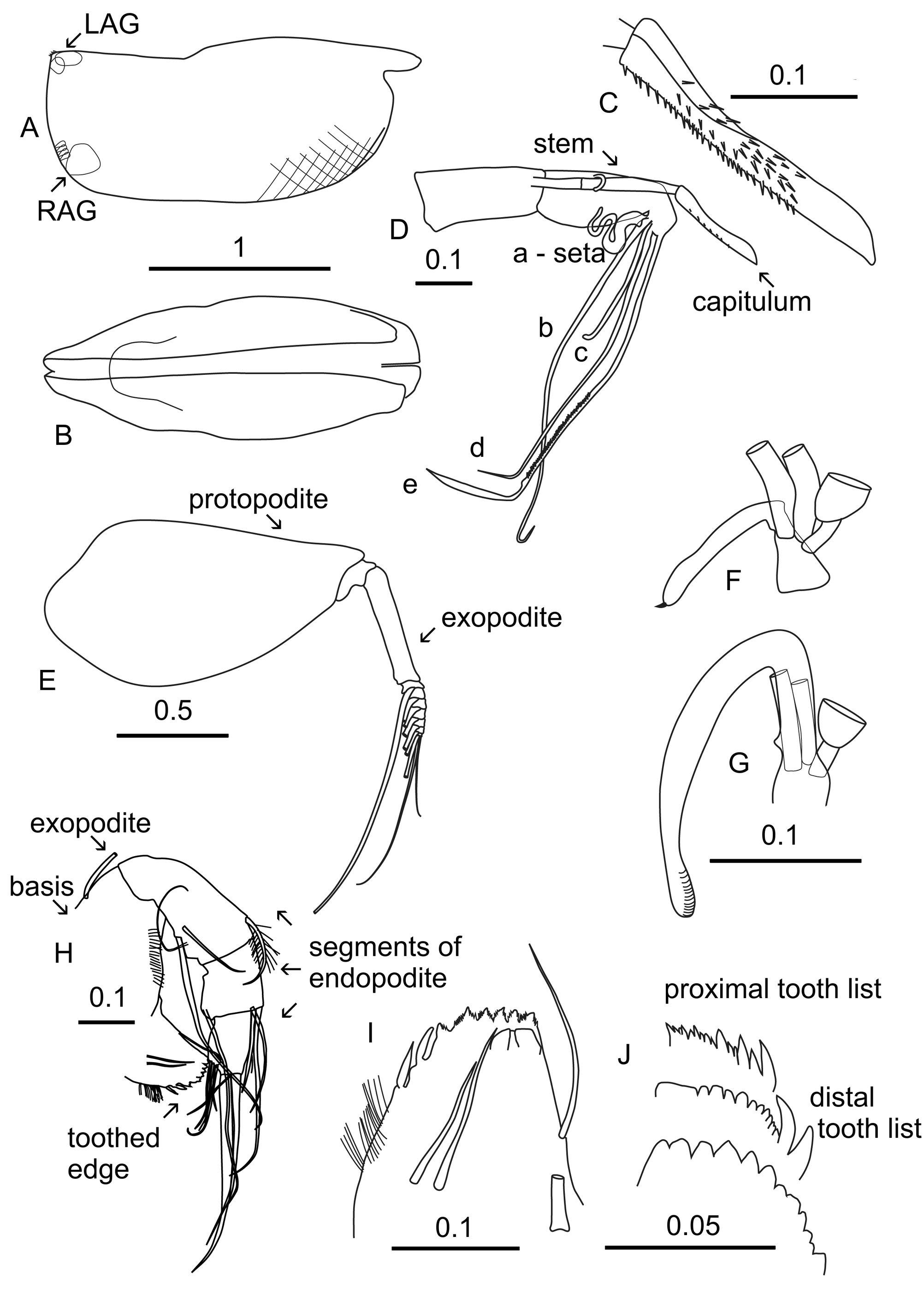
Carapace ( Figures 1 A,B View FIGURE 1 ). Length 2.13 ± 0.071 mm (see Table 1) In the North-East Atlantic, there is a trend for lengths to be greater at higher latitudes (the mean length of specimens from off Svalbard was 2.30 mm compared to 2.05 mm from 44°N) ( Table 4). The shoulder vaults are strongly arched and sharply edged, and the posterior margins of the vaults curve smoothly on to the dorsal margin. There is a criss-crossing diamond pattern of ornamentation, which is clearest in the anterior ventral region of the carapace, but becomes obscure posteriorly. The RAG opens slightly above the PVC and on both valves immediately dorsal to the RAG, there is a large group of lateral glands. Close to the PDC and ~ 0.29 mm below the hinge is a further group of lateral glands that open on the posterior margin on each valve. Along the posterior half of the ventral margin edge glands are well developed. The PDC is a rounded right angle and both valves are armed with 3 or 4 small spines.
Frontal organ ( Figures 1 C,D View FIGURE 1 ). The stem is relatively long (32.57% CL), is articulated about 0.23 mm from distal end and just posterior to where the retaining dorsal seta of antenna 1 locks on to the stem. The capitulum (12.38% CL) ( Figure 1 C View FIGURE 1 ) is clearly articulated and is slightly angled down. Its ventral margin is almost straight, whereas its dorsal margin is slightly concave and arches ventrally close to the pointed tip. It is spinous along the proximal two-thirds of the ventral margin and the proximal half of the dorsal margin and sides.
First antenna ( Figure 1 D View FIGURE 1 ). The length of the limb is 32.62% CL. The first two segments are clearly sutured. The second segment carries a medial seta, which locks the stem of the frontal organ to the limb. The a-seta (17.94% CL) is swollen and convoluted at its base. The b-seta (49.44% CL) carries a long-ribbed process level with the eseta armature; beyond the ribbed process the seta is slightly flanged and the flange carries ~6 short spinules. The cseta is moderately long (16.2% CL); the longest relative length of this seta in all Boroecia species ( Table 3). The dseta (35.0% CL) has ~35 very fine spinules along its trailing edge, and along its leading edge is a line of 11 short spinules followed by 4 longer ones, then 6 short fine spinules line the short, flanged end. The armature of the e-seta (48.98% CL) consists of approximately 44 pairs of closely spaced, fine spines that are aligned close to the seta, pointing proximally, making them difficult to count accurately. Distal to the armature, the seta is hinged before its long broadly flanged terminal section (c.f. Skogsberg 1920, Plate CXXXV, Fig. 4 View FIGURE 4 ).
Second antenna ( Figures 1 E,F,G View FIGURE 1 ). The protopodite is 48.95% CL, and is more than three times the length of the first exopodite segment (15.69% CL). The first segment is more than double the combined lengths of the remaining segments (7.2% CL) that carry the swimming setae. At the distal end of the first exopodite segment is a small terminal seta that wraps around the articulation with the second segment. The processus mamillaris on the endopodite is shaped like an arrowhead with an acute sharp point. The a- and b-setae are bare and nearly straight. The a-seta is ~75% the length of the b-seta. The c- and d-setae are bare and slim and a little shorter than the width of the second segment. There is a small curved e-seta inserted at the base of the g-seta. The g-seta (43.37% CL) is bare and clearly flanged towards its end, the shorter f-seta (33.92% CL) is similar but less obviously flanged. The left hook appendage ( Figure 1 F View FIGURE 1 ) has a short base with a tubercle on the outer corner of the right angle. The distal arm is short and slightly curved terminating in a smooth rounded end, carrying a tiny curved thorn. The right hook appendage ( Figure 1 G View FIGURE 1 ) is much larger; its basal section is longer than the second endopodite segment and has two small inner tubercles. It is angled at about 135°, and the distal process is long, curves back a further 45°, and is slightly thickened centrally. The end is rounded and extensively ridged sub-terminally.
Labrum. It has a shallow rounded notch, and is flanked by ~ 17–20 filaments on each side.
Mandible ( Figure 1 H View FIGURE 1 ). This limb has a general form that is typical of other Conchoeciniids. The first segment of the endopodite has three setae on its inner (ventral) surface, one of which is long and extends well beyond the end of the limb and two shorter setae. At the distal end of its outer margin is a short plumose seta. On the distal end of the second segment are three setae on the outer dorsal side, and a further two on the inner dorsal side. The terminal segment has 2 long unequal claw setae, with a thin short seta between them, and four slim short setae on its inner ventral side. The length of the longest claw seta (17.2% CL) is nearly 70% of the length of the outer edge of exopodite.
The toothed edge of the basale ( Figure 1 I View FIGURE 1 ) has the usual two spine teeth on its inner edge, one sharp pointed and one spindle-shaped with a blunt end. Both these teeth are bare, thus lack any secondary spines (in most other genera one or both of these teeth are armed with secondary spines). There are then six subserrate triangular teeth: the first tooth next to the spine setae is offset from the others (another generic characteristic). There is a broad inner tooth that overlaps the three outermost small teeth. The pars incisiva on the coxale ( Figure 1 J View FIGURE 1 ) consists of nine bluntly pointed teeth, the innermost being broadly flat edged, the others diminish in size along the list. In some specimens, these teeth are worn and indistinct. The distal tooth list consists of two large teeth followed by about 13 smaller teeth, the innermost of which is broad. The proximal list consists of three alternating sets of large and small teeth followed by 13 smaller and unevenly sized teeth.
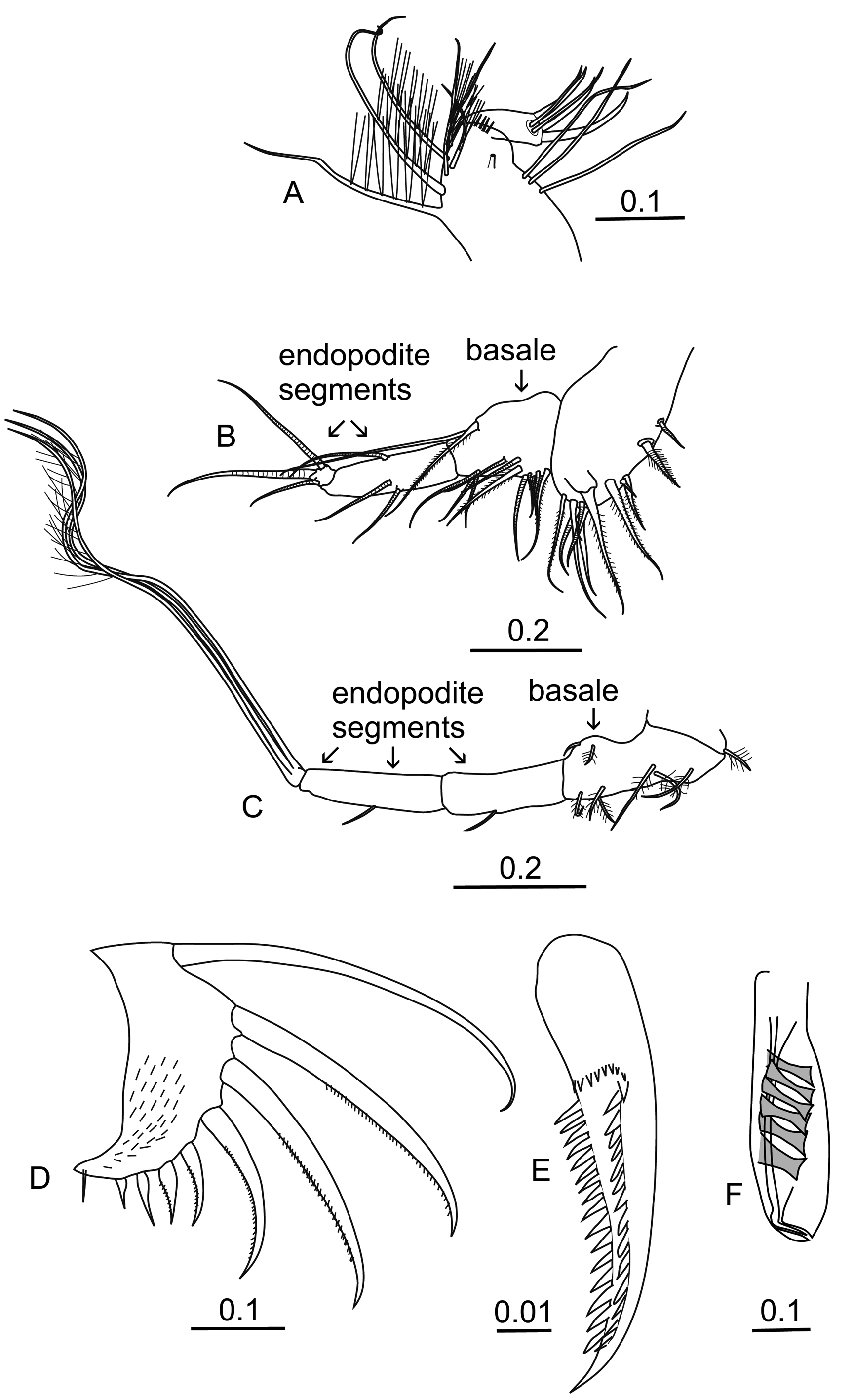
Maxilla ( Figure 2 A View FIGURE 2 ). The first exopodite segment carries six anterior setae, one lateral seta and three posterior setae. On the outer distal edge is a row of three or four short spines. The terminal segment carries the standard three claw setae with two offset slim setae.
Fifth limb ( Figure 2 B View FIGURE 2 ). The epipodial setal formula (dorsal to ventral) is 5 + 5 + 4. On the ventral edge of the basale there is a distal group of five setae (one of which is plumose) and a further three setae distally. Proximally on its lateral surface are two lateral setae that point divergently and a further distal seta. On the dorsal surface are two setae: a long plumose seta and close to the end of the segment an even longer bare seta that reaches beyond the tip of the limb. The first endopodite segment has a pair of ventral medial setae and a single medial dorsal seta. The second endopodite segment carries the three terminal claw setae that are typical of all halocyprids, the central one is stouter and longer than the other two. The longest seta is 8.6% CL.
Sixth limb ( Figure 2 C View FIGURE 2 ). The epipodial seta formula is 7 + 5 + 5. On the ventral surface of the basale there are two proximal setae and another three more distally; all these ventral setae are plumose. Laterally it carries at least one more plumose seta. On its dorsal surface, the bare terminal seta is very short as is the plumose seta. The first endopodite segment has only a single medial seta on its ventral surface, whereas the second segment has a single medial seta on both the dorsal (missing from the specimen drawn) and the ventral surface. The third endopodite terminal segment has the usual three long sub-equal terminal setae (30.9% CL), whose lengths are just over 60% of the dorsal length of the limb. The two dorsal setae have long setules along the distal halves, whereas the ventral seta appears to be bare.
Copulatory appendage ( Figure 2 F View FIGURE 2 ). The anterior margin is almost straight, whereas the posterior margin is convexly curved so that the maximum width is at about 2/3s the length. There are 6 oblique muscles and the end is bluntly rounded.
Caudal furca ( Figure 2 D View FIGURE 2 ). There are the usual 8 pairs of hook setae. The first longest pair have the long, slim, curved tips, which represent a generic character. The lengths of the first pair of hook setae are about 20% of the carapace. Another generic characteristic is the marked disparity in the lengths of the 4th and 5th pairs; the fifth pair is less than half the length of the fourth pair ( Table 2). The 7th and 8th pairs of hook setae are bare, this is a generic character except for B. alaska (in all other Conchoecinae genera examined either both the 7th and 8th pairs are spinose, or just the 8th pair are bare). There is an unpaired seta, whose length is similar to the 5th pair of hooks. The inner surfaces of the two caudal lamellae are covered with fine setules.
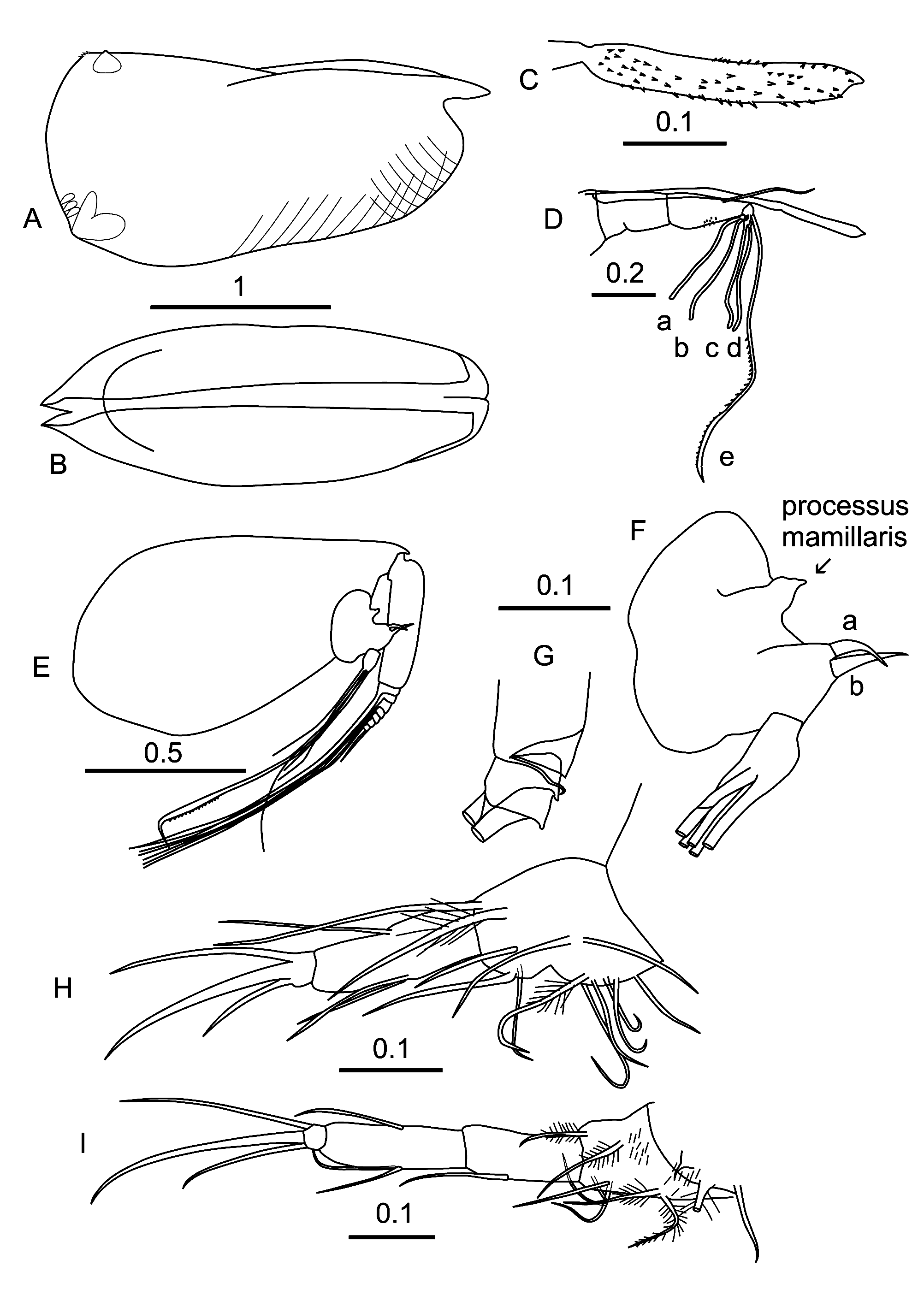
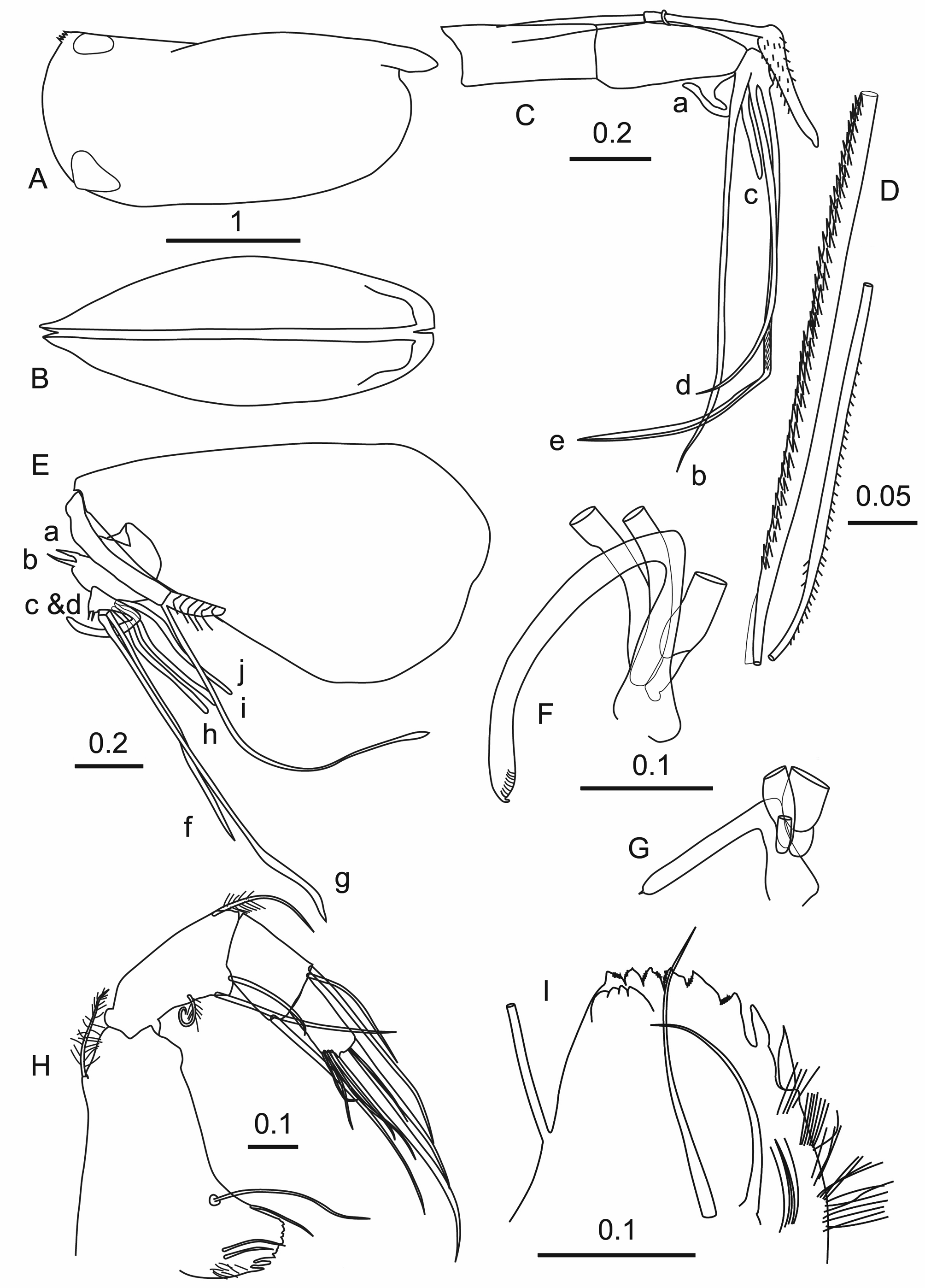
Female. Carapace ( Figures 3 A,B View FIGURE 3 ). The lateral outline of the carapace is similar to the male’s. However, it is longer (mean 2.53 ± 0.084 mm, see Table 1); the carapaces tend to be slightly longer at higher latitudes ( Table 4). Relative to the length, the carapace height is large (47.67% CL) with the maximum height at about 3/4s the length. The carapace tapers anteriorly and the width is less than in the male (width=37.92% CL). The gland positions and types are similar to those of the male and the shoulder vaults are clearly sharply edged. There is a clear criss-cross ornamentation on the anterior half.
Frontal organ ( Figures 3 C,D View FIGURE 3 ). The stem (21.08% CL) is longer than the first antenna and is unsutured. The suture between the stem and the capitulum is also not clearly defined. The capitulum ( Figure 3 C View FIGURE 3 ) is slightly curved and relatively short (10.75% CL). It is only slightly thicker than the shaft, and sparsely spinose; the spines along its ventral surface are coarser than those on the dorsal surface.
First antenna ( Figure 3 D View FIGURE 3 ). The segments of the first antenna are weakly differentiated by partial sutures; the limb’s overall length is 18.67% CL. The dorsal seta (14.27% CL) is bare and long; it almost reaches level with the tip of the capitulum. The a–d-setae (14.20% CL) have short shafts and are about a third the length of the e-seta (36.76% CL). The e-seta is slightly flanged distal to the ends of the a–d-setae, and the trailing edge of the flange is lined with short fine spinules.
Second antenna ( Figure 3 E View FIGURE 3 ). The protopodite is 43.46% CL. The first exopodite segment is 15.68% CL and its eight distal segments that carry the swimming setae together form 6.87% CL. The first exopodite segment has a very short terminal seta that is wrapped around the suture with the second segment ( Figure 3 F View FIGURE 3 ). Both endopodites have a triangular processus mamillaris with a terminal nipple-like tip. The a- and b-setae are similar to those of the male, but the c-, d-, and e-setae are absent. Both the f-seta (37.05% CL) and g-seta (26.84% CL) are slightly flanged terminally, and the flanges are lined with fine spinules. The h–j-setae are 21.10% CL.
Mandible and maxilla. These limbs are not sexually dimorphic and are the same as in the male. The relative length of terminal seta of the mandible is almost identical at 17.4% CL.
Fifth limb ( Figure 3 H View FIGURE 3 ). The setation of this limb is similar to the male’s. Its longest terminal seta is 7.7% CL.
Sixth limb ( Figure 3 I View FIGURE 3 ). The epipodial setal formula is 7 + 5 + 5. On the ventral surface of the basale five plumose setae (2 + 2 + 1) are inserted, and dorsally is a plumose seta and a short bare seta. The first endopodite segment has a single medial ventral seta, but no dorsal seta. The second endopodite segment has two medial seta on ventral side and one seta on dorsal surface. The last segment has three terminal claw setae, central one is the longest (14.9% CL, which is about 2/3s the length of the dorsal edge of the limb) and the shortest ventral seta is just over half its length.
......continued on the next page Longestseta Geographical variation. Carapace length data of B. borealis specimens collected from different localities in the North-East Atlantic ( Table 4) are extracted from material mostly from along 20°W ( Angel & Fasham 1975), but also from repeated sampling at 44°N 13°W (Angel 1977, 1984). Further data come from material collected at Hudson Station L3–17 ( 10/6/2001), where the southward flow of the East Greenland Current results in much lower water temperatures off Newfoundland. The table shows that there is a slight diminution in the size of this species from higher to lower latitudes in the North Atlantic, but the Hudson data suggest that this is related to water temperature. A few of the specimens collected at Station “M” (65°N 20°W) in the Norwegian Sea, where water temperatures are cooler, were also marginally larger than those from 60°N. The growth factors (i.e. the ratios between the mean carapace lengths of successive instars) are consistently 1.41–1.50 except between the final juvenile instar and adult males. The increase in biomass is probably much the same because the adult male is relatively wider. Similar growth factors have been reported in other halocyprid species (Angel 1977, Ikeda 1992, Drapun 2002).
In their web-based atlas of Atlantic planktonic ostracods Angel et al. (2008) show plots of published records of the zoogeographical range of this species, which indicate that it is endemic to the North Atlantic. However , recent records from high latitudes in the Arctic show that it occurs sporadically in Arctic regions well beyond those directly influenced by inflows of North Atlantic Water. For example, two adult females occurred in samples collected from the fringes of the Canadian Deep to the north of the Bering Strait (Healy 05/2 station 14, 22 July 2005, 200– 500m, 74° 18.5'N; 159° 55'W). These adults were persisting well beyond their predominantly recorded distribution suggesting that future slight changes in oceanic conditions resulting from climate change may result in changes in the geographical range of this and perhaps also other species. GoogleMaps

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SubOrder |
Halocypridina |
|
Family |
|
|
SubFamily |
Conchoeciinae |
|
Genus |