
Flabelligera affinis Sars, 1829
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3203.1.1 |
|
persistent identifier |
https://treatment.plazi.org/id/8C476837-FFEC-FFEA-FF79-F92DFC2EFA6A |
|
treatment provided by |
Felipe |
|
scientific name |
Flabelligera affinis Sars, 1829 |
| status |
|
Flabelligera affinis Sars, 1829 View in CoL
Figure 1 View FIGURE 1
Flabelligera affinis Sars 1829:31–34 View in CoL , Pl. 3, Figs. 16–19 View FIGURE 16 View FIGURE 17 View FIGURE 18 View FIGURE 19 ; Cunningham & Ramage 1888:677–678, Pl. 46, Fig. 40A–B; de Saint- Joseph, 1894:96–101, Pl. 5, Figs. 121–123 (partim); Newbigin 1900:190 (partim); Moore 1909b:143; Gunther 1912:11– 12; Haase 1915:183–186, Pls. 1–2, Textfig. 1; McIntosh 1915:107–114, Pl. 89, Fig. 2 View FIGURE 2 , Pl. 96, Fig. 10 View FIGURE 10 , Pl. 104, Figs. 7–7b View FIGURE 7 (syn.); Chamberlin 1919b:398–399; Fauvel 1927:113, Figs. 40a–f; Monro 1930:160, Fig. 63; Rioja-LoBianco 1931:88– 90, Pl. 24, Figs. 1–3 View FIGURE 1 View FIGURE 2 View FIGURE 3 ; Monro 1936:164; Støp-Bowitz 1948a:8–13, Fig. 1 View FIGURE 1 & map; Støp-Bowitz 1948b:30–33, map; Wesen- berg-Lund 1950:35; Pettibone 1954:289–290, Fig. 33e–g (partim); Pettibone 1956:563; Hartmann-Schröder 1971:370– 372, Fig. 130; Amouroux 1974:179–180; Hartmann-Schröder 1996:417–419, Fig. 203; Jirkov & Filippova 2001:360–361, Figs. 1–7o View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 ; Oug et al. 2011:6, Figs. a, b.
Chloraema dujardini de Quatrefages 1849:282–285 View in CoL , Pl. 9, Figs. 1–2 View FIGURE 1 View FIGURE 2 , Pl. 10, Figs. 1–5 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 .
Chloraema pellucidum Sars 1869:253 View in CoL nomen nudum.
Chloraema pellucidum Sars View in CoL in Sars 1872:409; 1873:252–261, Pl. 16, Figs. 9–20 View FIGURE 9 View FIGURE 10 View FIGURE 11 View FIGURE 12 View FIGURE 13 View FIGURE 14 View FIGURE 15 View FIGURE 16 View FIGURE 17 View FIGURE 18 View FIGURE 19 View FIGURE 20 .
Type material. Northwestern France. Syntypes of Chloraema dujardini de Quatrefages, 1849 ( MNHN-465 ), damaged, St. Vaast (Saint-Vaast-la-Hogue, 49º35'19" N, 01º15'58" W), Normandy GoogleMaps .
Additional material. Norway. Two specimens ( ZMUB-2253 ), Tromsø, no further data, Danielsen, coll. (largest specimen used for description). Two specimens ( ZMUB-2263 ), damaged, Holmer Graa, on corals, Friele & Hansen, coll. Three specimens ( ZMUB-18636 ), heavily contracted, tunic transparent, Tfjinsgaarden, Stat. 40, App., coll. An anterior fragment ( ZMH-23598 ), Liefdefjorden, Spitzbergen, SPE-92/68, 7 Aug. 1992, Fucusstation, Taucherprobe, Plante & Wagner, coll . Greenland. Several specimens ( USNM-148624 ), Walrus Grounds, Murchison Sound , NW Greenland (77°45' N), first haul otter trawl, 45 fathoms, 7 Aug. 1938, number 127, R.A. Bartlett coll. (some specimens without tunic; one with body broken by pharynx contraction, has about 20 branchial filaments). Several specimens ( USNM-148624 ), Walrus Grounds, Murchison Sound, NW Greenland (77°38' N), otter trawl, 60 fathoms, 8 Aug. 1938, number 144, R.A. Bartlett coll. One mature female ( USNM-165293 ), residue of dredge, see oil tin No. 1, 9 Aug. 1943, number 3, R.A. Bartlett, coll. (22 chaetigers, no cephalic cage; 2–3 transverse rows of clavate papillae) . Iceland. One specimen ( MNHN-184 a), most tunic removed, Patieksfjorde, R/ V Pourquoi-pas?, Stat. 10 bis, 5 m, 22 Aug. 1929 (60 mm long, 5.5 mm wide, cephalic cage 2.5 mm long, 58 chaetigers). One specimen ( MNHN-184 b), R/ V La Manche Expedition, 1892 (no further data) (noto- and neuropodial lobes distinct) . Ireland. One specimen ( MNHN-457 ), mature female, Clew Bay, West Ireland , no more data, P. Fauvel, coll . France. One specimen ( MNHN-457 b), St. Anne, Cherbourg , Aug. 1916, P. Fauvel, coll. An anterior fragment ( MNHN-457 c), Urville , Regnéville-sur-Mer , Basse Normandie, 18 Jul. 1913, P. Fauvel, coll. One specimen ( MNHN-457 d), Cherbourg , 19 Apr. 1924, P. Fauvel, coll. (tunic with two large nematodes and a large rounded vesicle, probably made by another type of parasite). Three specimens ( MNHN-507 a), brownish, compressed, many neurochaetae broken, Tathiou, Apr. 1895, no further data. One specimen ( MNHN-507 ), without posterior region, anterior end exposed, distorted, eroded, Urville (Urville-Nacqueville, 49º40'29” N, 01º44'15” W), 18 Jul. 1913, P. Fauvel, coll. One specimen ( MNHN-507 b), splendid, St. Anne , Cherbourg, Aug. 1916, P. Fauvel, coll. One specimen ( MNHN-507 c), twisted, partially dehydrated, anterior end exposed, Cherbourg, 19 Apr. 1924, P. Fauvel, coll. (large nematode emerging from the tunic dorsally, and a globular blister or cyst, ventrally, in chaetiger 11). Arctic GoogleMaps Canada. One specimen ( USNM-11181 ), gift by Chas Lütken, ZMUC. Several specimens ( USNM-246975 ), damaged, Stat. 720 (69°21.5' N, 81°40.5' W), B21/P72, Fore Basin, 8–9 m, 10 Oct. 1955, Grainger , coll. (two specimens with two different nematodes in the tunic, one black and white, the other pink) GoogleMaps .
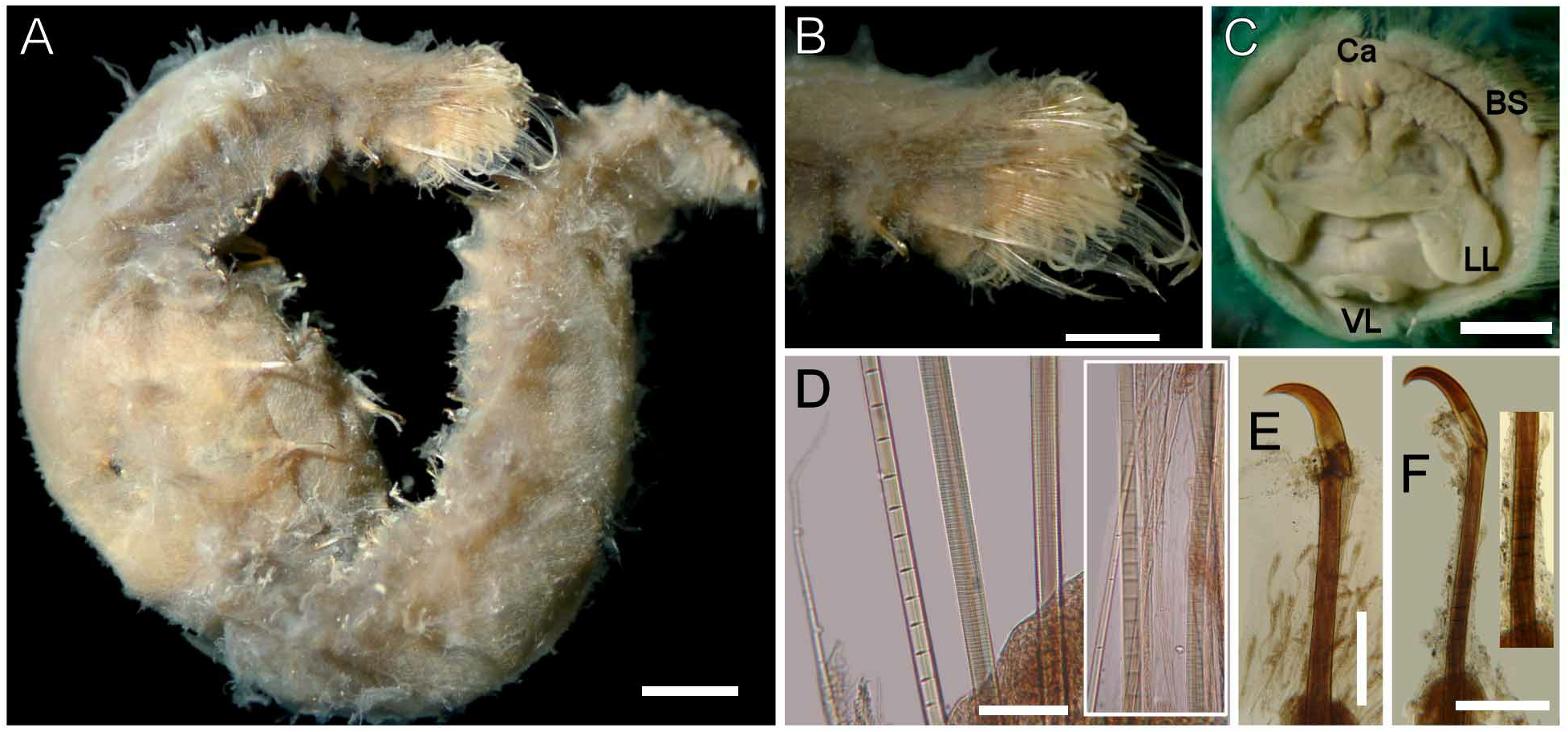
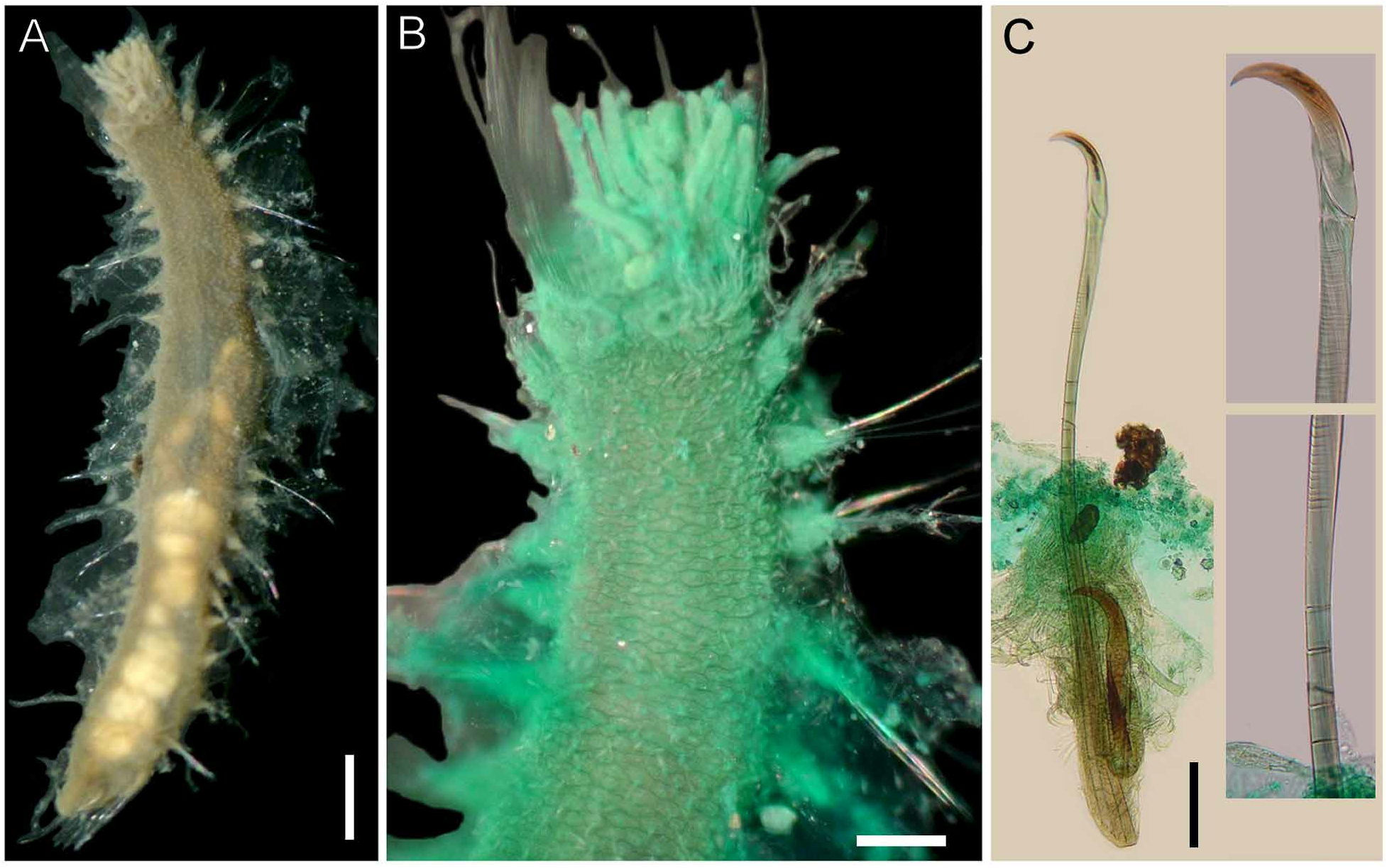
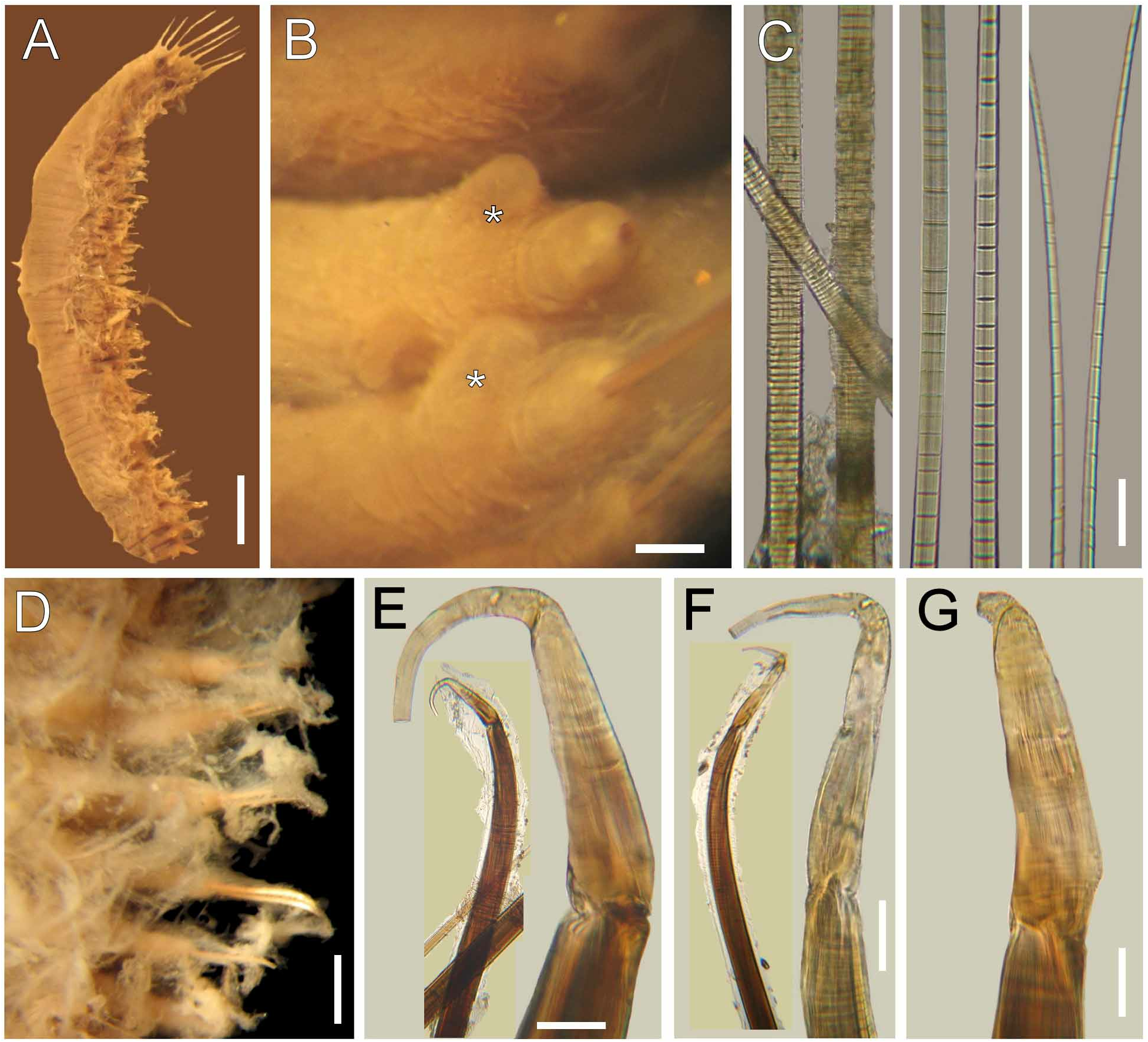
Description. Body pale (ZMUB-2253), tapering posteriorly ( Fig. 1A, E View FIGURE 1 ); 57 mm long, 7 mm wide, cephalic cage 5 mm long, 52 chaetigers. Tunic thick, transparent, densely papillated, somehow damaged, with cover of fine sediment particles; papillae long, clavate, mucronate.
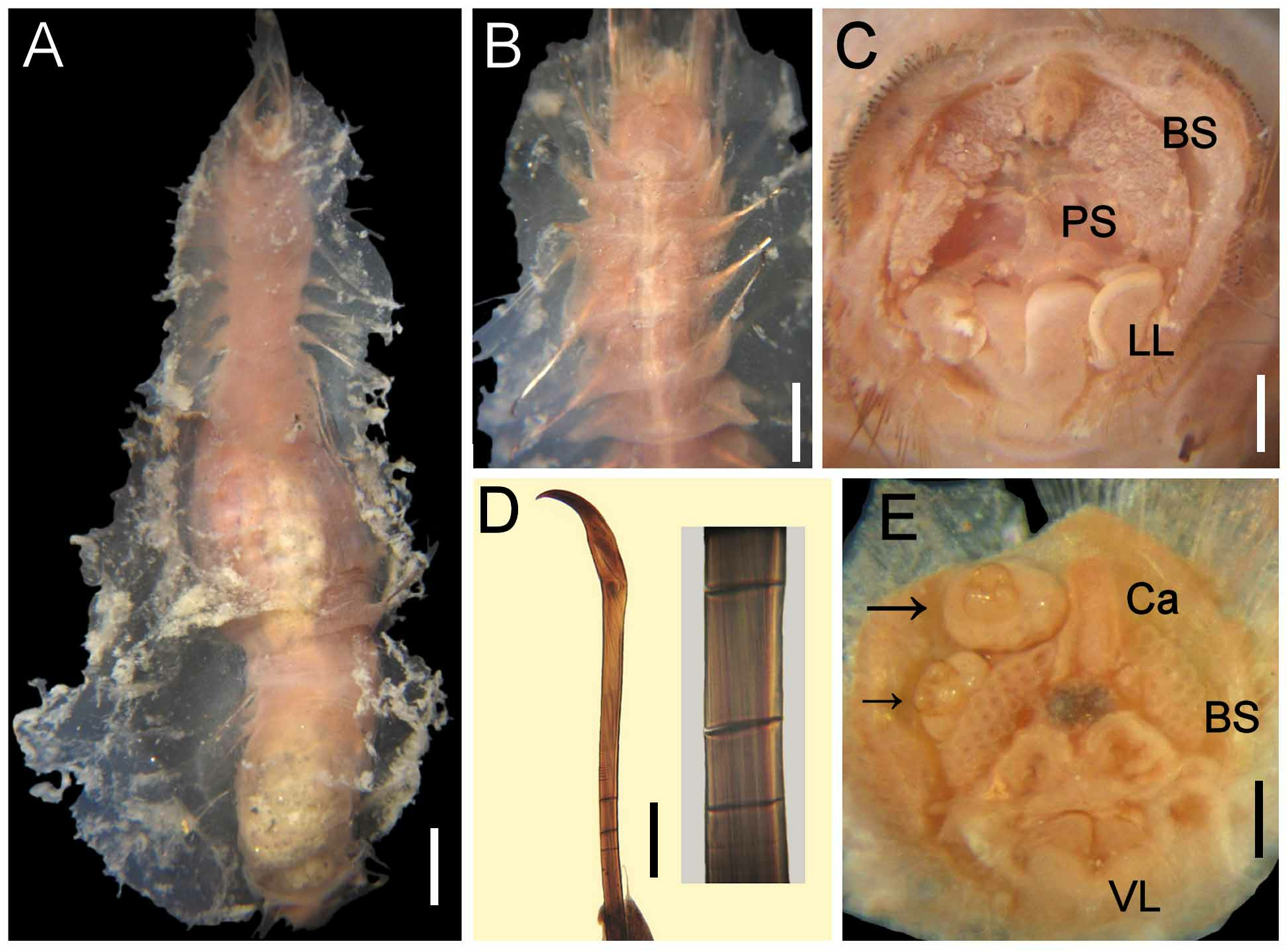
Cephalic hood not exposed. Prostomium elevated, rounded, with four dark, sometimes faded eyes. Caruncle well developed, wide, exceeding the branchial plate margin, distally truncate ( Fig. 1D View FIGURE 1 ). Palps pale, thick, twice as long as branchiae, about four times thicker than branchiae, often exposed ventrally ( Fig. 1C View FIGURE 1 ); palp bases rounded, elevated. Lateral lips well developed, fused to dorsal lip; ventral lip triangular, without ventral projection.
Branchiae in two lateral groups, each with filaments in concentric rows, each group with about 60 filaments. Longest branchiae about half as long as palps. Nephridial lobes paired, lateral to caruncle, about the median distance of branchial groups ( Fig. 1D View FIGURE 1 ).
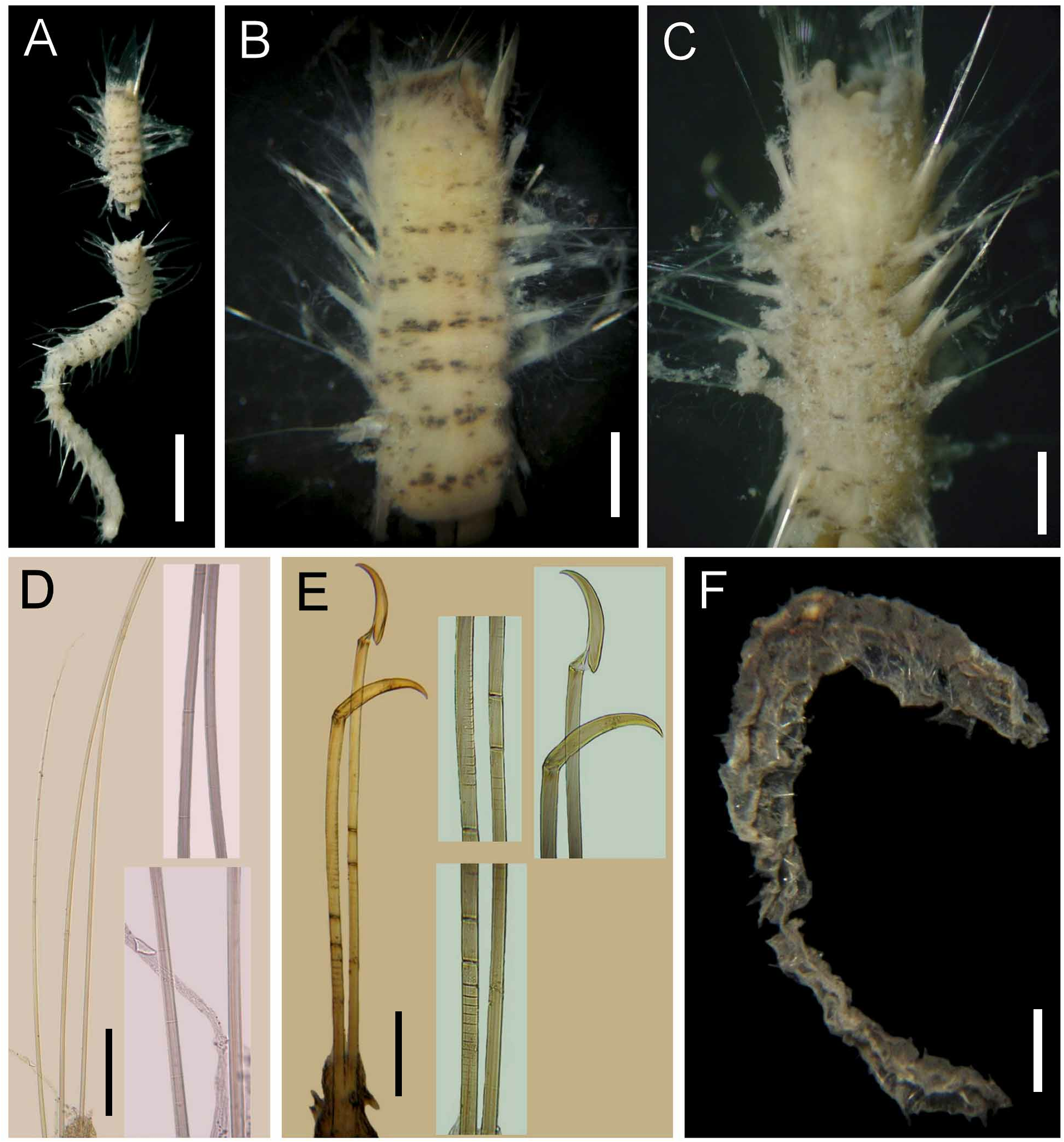
Cephalic cage chaetae about 1/5–1/9 body length, slightly shorter than body width ( Fig. 1B View FIGURE 1 ). Chaetiger 1 involved in cephalic cage, with about 34 noto- and 20 neurochaetae per side. Anterior dorsal margin of chaetiger 1 papillated. Anterior chaetigers without especially long papillae. Chaetiger 1 longer than chaetigers 2–3. Chaetal transition from cephalic cage to body chaetae abrupt, chaetiger 2 with multiarticulated neurohooks. Gonopodial lobes present in chaetigers 5 and 6.
Parapodia well developed, lateral; median neuropodia ventrolateral. Notopodial lobes projecting, basally rounded, distally depressed, truncate, without different papillae. Neuropodial lobes projected, digitate, without different papillae. Noto- and neuropodia widely separated.
Median notochaetae arranged in short transverse rows; all notochaetae multiarticulated capillaries, as long as 1/3 body width,7–8 per bundle, articles best developed distally, very short throughout all the chaetae ( Fig. 1F View FIGURE 1 ), some chaetae with articles ill-defined. Neurochaetae multiarticulated capillaries in chaetiger 1. Multiarticulated neurohooks from chaetiger 2 ( Fig. 1G View FIGURE 1 ), one per ramus (most with a replacement hook). Handle articulation medially placed, articles irregular (Fibonacci sequence); three articles: small, medium-sized and long. Other articles anchylosed, small, continued to bending region. Crest slightly wider than handle, yellowish in anterior chaetigers, with darker distal half in median and posterior chaetigers; width:length ratio 1:4.
Posterior end truncated; pygidium with anus terminal, not covered by tunic, thick muscular ring, without anal cirri.
Variation. Two adult specimens of very different sizes (ZMUB-2253) show that the cephalic cage is sizedependent. The number of chaetigers was the same (52) but one specimen was three times longer than the other (22 mm vs 57 mm). The articulation of the neurohook handle is a consistent feature. The specimens have 2–3 distinct, larger articles and although the syntypes are very small, and the number of articles might be regarded as sizedependent, the number of large articles is not.
Remarks. Flabelligera affinis Sars, 1829 resembles F. orensanzi n. sp. They differ because in F. affinis the tunic does not include sediment particles and the neurohooks have pale, tapering crests, whereas in F. orensanzi n. sp. the tunic often has a slight sediment cover and neurohooks crests are medially widened and dark red.
The original description of F. affinis by Sars (1829) indicated his materials were from rocky and muddy bottoms and that it had a leathery dirty-green tunic. Rathke (1843) described Siphonostoma vaginiferum from muddy bottoms, and indicated it has an opaque, rugose tunic. Rathke’s approach is herein followed to restrict F. affinis to the Flabelligera specimens with a transparent tunic, whereas those having an opaque tunic are regarded as belonging to a distinct species, F. vaginifera (see below). Chloraema pellucidum Sars in Sars, 1872, was regarded as a junior synonym of F. affinis by Haase (1915) and later authors. However, it was originally recognized as an independent species by Michael Sars, who had also described F. affinis . Because F. affinis has been restricted to include specimens with a mostly transparent tunic, C. pellucidum is regarded as conspecific. The syntypes of C. dujardini (MNHN-465) are damaged juveniles, one broken into two pieces, and many neurohooks are broken in both specimens. Their bodies are long, fusiform, compressed, tapered posteriorly; the tunics are damaged, but without sediment particles, transparent, leaving chaetal tips uncovered. They are 8.5–10.0 mm long, 1 mm wide, cephalic cage 1 mm long, 29–33 chaetigers. Newbigin (1900:190, 195) concluded that F. affinis includes F. dujardini , F. diplochaitus (Otto) , and F. claparedi (de Saint-Joseph) . He was correct for the first species only because F. diplochaitus is herein regarded as a distinct species, whereas F. claparedei has been transferred to Flabelliderma Hartman, 1969 ( Salazar-Vallejo 2007).
Distribution. As currently recognized, F. affinis is distributed from Arctic to cold and temperate, boreal localities. It has been recorded from many widespread localities; such records by Fauvel & Rullier (1957:376, 1959a:180, 1959b:958) for Western Africa, as well as those for Southern Africa by Day (1961:505, Fig. 8a View FIGURE 8 ), or by Monro (1933:1056–1057, Text fig. 5) for Panama, are all questionable. Likewise, the records by Chamberlin (1920:21) for the Canadian Arctic, or those by Ushakov (1955:306/ 1965:283, Fig. 113A–D), and by Hobson & Banse (1981:58, Fig. 11e View FIGURE 11 ), for the Northern Pacific Ocean, belong elsewhere.











No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Flabelligera affinis Sars, 1829
| Salazar-Vallejo, Sergio I. 2012 |
Chloraema pellucidum
| Sars, G. O. 1873: 252 |
| Sars, G. O. 1872: 409 |
Chloraema pellucidum
| Sars, M. 1869: 253 |
Chloraema dujardini
| de Quatrefages, A. 1849: 285 |
Flabelligera affinis
| Oug, E. & T. Bakken & J. A. Kongsrud 2011: 6 |
| Jirkov, I. A. & A. V. Filippova 2001: 360 |
| Hartmann-Schroder, G. 1996: 417 |
| Amouroux, J. M. 1974: 179 |
| Hartmann-Schroder, G. 1971: 370 |
| Pettibone, M. H. 1956: 563 |
| Pettibone, M. H. 1954: 289 |
| Stop-Bowitz, C. 1948: 8 |
| Stop-Bowitz, C. 1948: 30 |
| Monro, C. C. A. 1936: 164 |
| Rioja-LoBianco, E. 1931: 88 |
| Monro, C. C. A. 1930: 160 |
| Fauvel, P. 1927: 113 |
| Chamberlin, R. V. 1919: 398 |
| Haase, P. 1915: 183 |
| McIntosh, W. C. 1915: 107 |
| Gunther, K. 1912: 11 |
| Moore, J. P. 1909: 143 |
| Newbigin, M. I. 1900: 190 |
| Cunningham, J. T. & G. A. Ramage 1888: 677 |
| Sars, M. 1829: 34 |