
Liladownsia fraile, Woller & Fontana & Mariño-Pérez & Song, 2014
|
publication ID |
https://doi.org/10.11646/zootaxa.3793.4.6 |
|
publication LSID |
lsid:zoobank.org:pub:1AFFE199-B80B-4DB0-B913-BF88132430B8 |
|
DOI |
https://doi.org/10.5281/zenodo.5082411 |
|
persistent identifier |
https://treatment.plazi.org/id/F9168457-7856-1668-FF11-3219A149FE8F |
|
treatment provided by |
Felipe |
|
scientific name |
Liladownsia fraile |
| status |
sp. nov. |
Liladownsia fraile sp. nov. Fontana, Mariño-Pérez, Woller & Song
http://lsid.speciesfile.org/urn:lsid: Orthoptera .speciesfile.org:TaxonName:463901
Taxonomic description (of male except where specified). General. Liladownsia fraile is a medium-large showy grasshopper. Body is primarily colored with blackish-steel blue, yellow, red, and sometimes orange depending on color form ( Figs. 2–4 View FIGURE 2 View FIGURE 3 View FIGURE 4 and 10,11). Both sexes are very colorful, and are characterized by wrinkled teguments, reticulate tegmina, and diffuse hair on the body, which makes them appear almost silky ( Figs. 2A&B View FIGURE 2 , 3A View FIGURE 3 , and 10 View FIGURE 10 ). The shape of the pronotum ( Figs. 2C View FIGURE 2 and 3B View FIGURE 3 ) and hind femora is very peculiar. Prozona (especially in females) with a bulging appearance and almost smooth while metazona is markedly wrinkled and raised posteriorly. The overall look of the pronotum resembles the lowered hood of a monk/friar ( fraile in Spanish) ( Figs. 2–4 View FIGURE 2 View FIGURE 3 View FIGURE 4 and 10 View FIGURE 10 ). Hind femora are thin in comparison to body size and general body structure ( Figs. 2A&B View FIGURE 2 , 3A View FIGURE 3 , and 10 View FIGURE 10 ). Contrary to the generally more complicated structure of the overall exterior, the male genitalia are quite simple. The external genitalia do not have any special features with the cerci subconical, absent furcula, and the supra-genital plate subtriangular ( Fig. 5 View FIGURE 5 ). The internal phallic complex also lacks any peculiarities ( Figs. 6 View FIGURE 6 and 7 View FIGURE 7 ).
Coloration. Two general color forms appear to exist across both sexes: a lighter color form with blackish-steel blue and yellow more dominant overall ( Figs. 2A View FIGURE 2 , 4A View FIGURE 4 , 10B View FIGURE 10 , and 11A&D View FIGURE 11 ) and a darker color form with blackishsteel blue, yellow, red, and sometimes orange more dominant overall ( Figs. 2B View FIGURE 2 , 3A View FIGURE 3 , 4B&C View FIGURE 4 , 10A View FIGURE 10 , and 11 B,C View FIGURE 11 ,&E). Head blackish-steel blue with fastigium, vertex, frontal ridge, and cheeks red, yellow, or orange. Antennae yellow to orange with first two segments blackish ( Figs. 2–4 View FIGURE 2 View FIGURE 3 View FIGURE 4 and 10 View FIGURE 10 ). Pronotum black on anterior margin, lateral sides of prozona, along and behind sulcus; prozona mainly red, orange, or yellow and metazona posteriorly red ( Figs. 2C View FIGURE 2 and 3B View FIGURE 3 ). Tegmina clear brown with blackish reticulation. Fore and middle femora red and/or yellow and blackishblue steel apically. Fore and middle tibia blackish-blue steel, all tarsi blackish-blue steel. Hind femora with blackish medial area on external side and red or yellow-orange upper and lower marginal areas. Apical portion of hind femora and knees entirely blackish-blue steel; hind tibia blackish-blue steel with reddish-yellow or yellow basal portion. Thorax brownish; abdomen reddish brown and terminalia in both sexes blackish ( Figs. 2A&B View FIGURE 2 , 3A View FIGURE 3 , and 10 View FIGURE 10 ).
Head. Head short, fastigium projecting moderately from eyes, widely- rounded when viewed laterally; frontal ridge well-marked with almost parallel lateral keels. Antennae filiform, semi-flattened with parallel sides; about 24 segments, median segments about 2.5 times as long as wide. Eyes scarcely prominent in females, not spaced very far apart; in males, more prominent, larger, and closer together. Prosternal process cylindro-conical, slightly bent posteriorly ( Figs. 2A&B View FIGURE 2 , 3A View FIGURE 3 , and 10 View FIGURE 10 ).
Pronotum and wings. Pronotum with swollen prozona, more pronounced in female; metazona arising posteriorly from sulcus, posterior margin widely rounded. Prozona as long as metazona in both sexes. Aside from some setae, prozona smooth with some scattered indentations. Metazona wrinkled and highly rugose. Sulcus very deep; median carinae present on prozona, but more prominent anteriorly, and also present on metazona ( Figs. 2C View FIGURE 2 and 3B View FIGURE 3 ). Tegmina brachypterous, tectiform, and widely oval; homogeneously reticulated; longer than pronotum and reaching midpoint of hind femora in both sexes; partially overlapping on dorsum in both sexes. Hind wings vestigial and more or less half the length of the tegmina ( Figs. 2A&B View FIGURE 2 , 3A View FIGURE 3 , and 10 View FIGURE 10 ).
Legs. Fore tibia with 3–5 spines on both inner and outer lower margins. Middle tibia with 3–5 spines on both outer and inner lower margins. Upper margins of hind tibia with 11–12 spines on outer side and 10–11 on inner side. Hind femora thin in relation to body size: 5.45 times as long as high in males and 6.5 times in females ( Figs. 2A&B View FIGURE 2 , 3A View FIGURE 3 , and 10 View FIGURE 10 ).
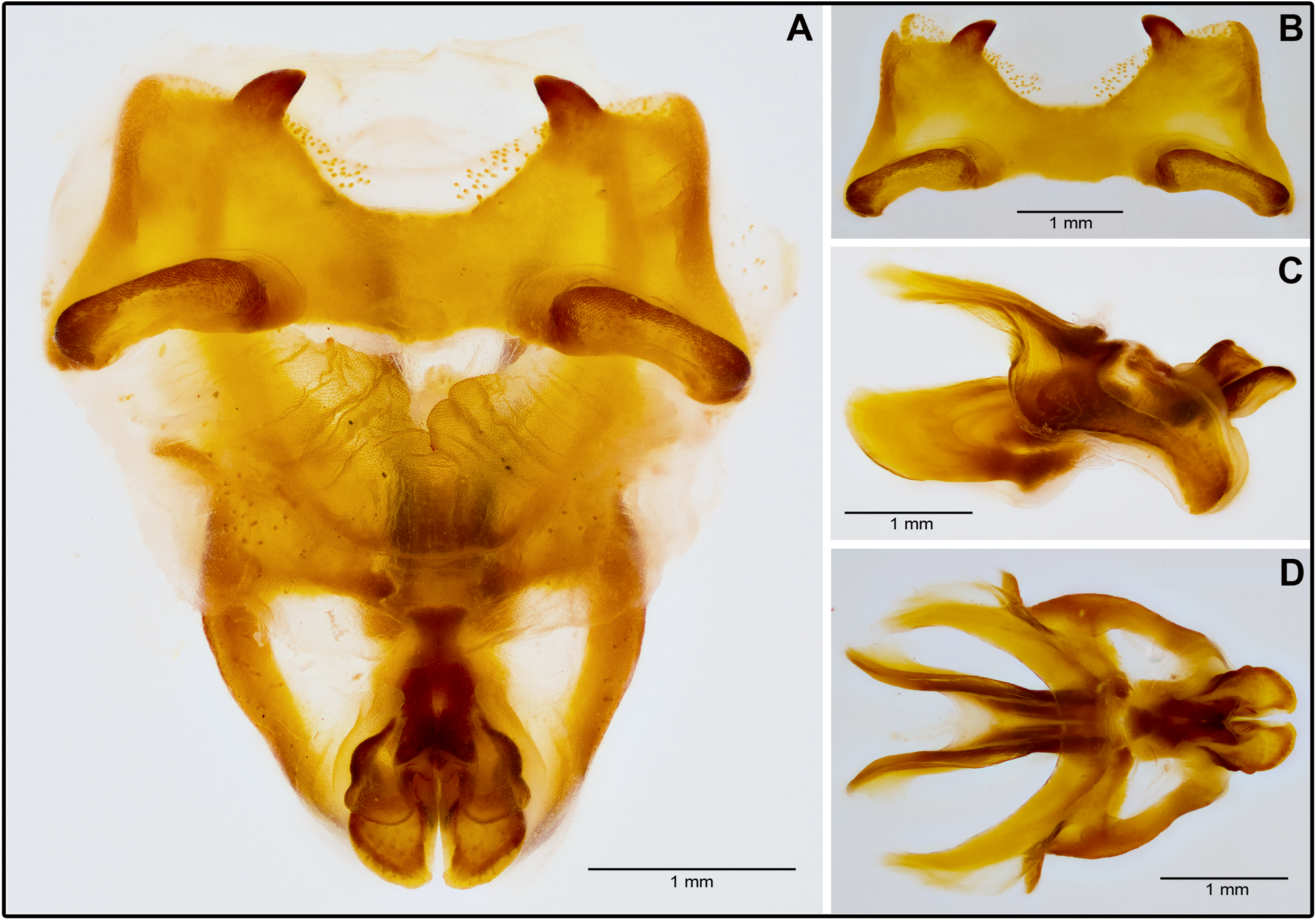
Terminalia. Male. Furcula almost absent; only two vestigial protuberances present. Supra-anal plate subtriangular, with rounded apex and slightly convex lateral sides. Median impression and median keels hardly developed ( Fig. 5B View FIGURE 5 ). Cerci subtriangular, gradually tapering from basal part; apex moderately flattened and rounded ( Fig. 5 View FIGURE 5 ). When viewed laterally ( Fig. 5A View FIGURE 5 ), subgenital plate resembling 1/4 of a sphere, hemielliptical from above; apex truncated and barely concave.
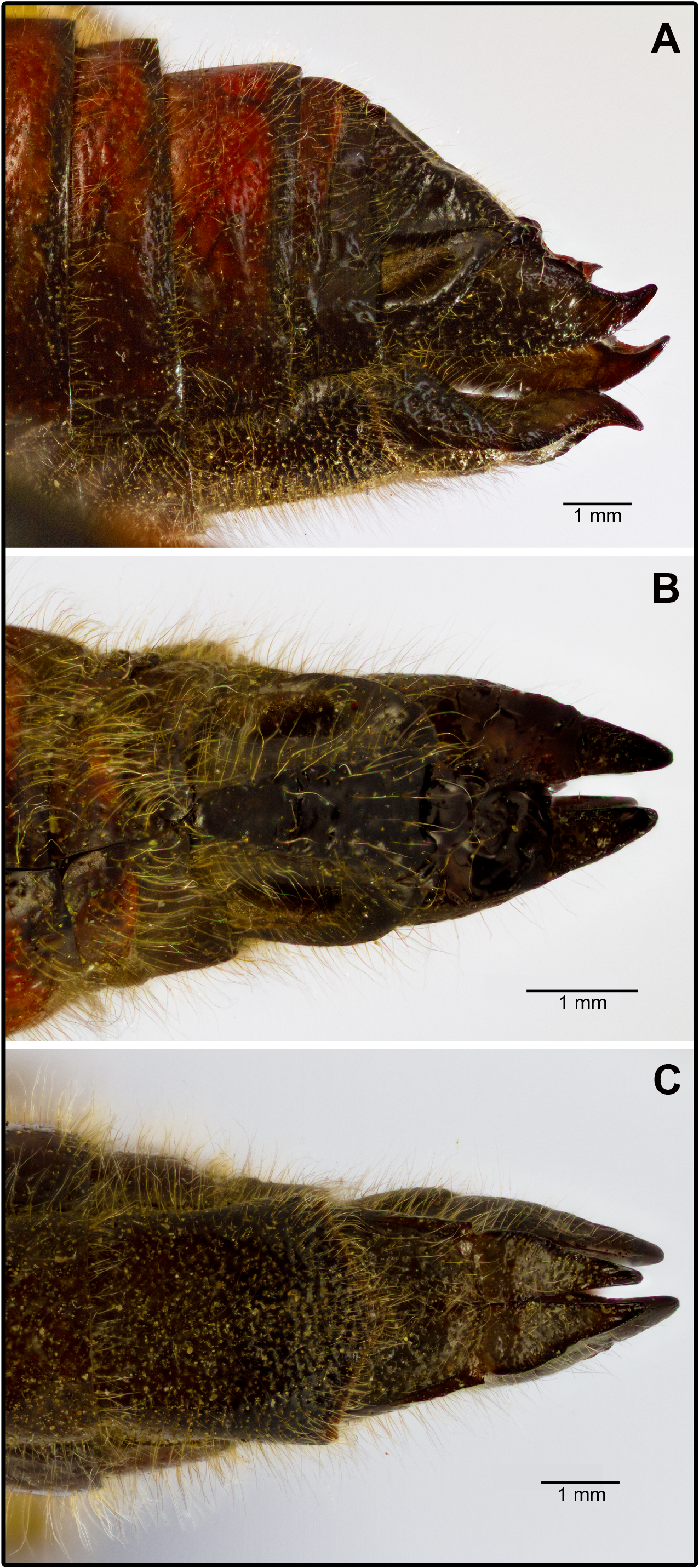
Female. Supra-anal plate subtriangular, apex rounded with concave lateral sides ( Fig. 8B View FIGURE 8 ). Dorsal valvae of the ovipositor ( Fig. 8A&B View FIGURE 8 ) almost twice as tall as ventral ones ( Fig. 8A&C View FIGURE 8 ). Lower margin of ventral valvae uniformly sinuous without teeth or hooks ( Fig. 8C View FIGURE 8 ). Subgenital plate with scarcely projecting subtriangular posterior margin and with concave lateral sides ( Fig. 8C View FIGURE 8 ).
Phallic complex. Epiphallus well-sclerotized; anterior margin of bridge concave; anterior spines short, conical, directed inwards and downwards, arched lophi with superior margin almost straight ( Figs. 6 View FIGURE 6 and 7A&B View FIGURE 7 ); well-elevated over anterior processes from a lateral viewpoint ( Fig. 6A View FIGURE 6 ). Valvae of penis short, subequal in length. Dorsal valvae subtrapezoidal; ventral valvae longer, transversally flattened ( Figs. 6 View FIGURE 6 and 7A,C View FIGURE 7 ,&D).
Etymology. This specific epithet comes from the common name used for this species by the local people of the type locality. In Spanish, “fraile” translates to monk or friar, referring to the swollen prozona, which resembles the hood of a monk’s robe. For the same reason some locals also refer to it as “chapulín de capucho” (hooded grasshopper). Based on this, we propose the following common name for this amazing grasshopper: Lila Downs’ friar grasshopper.
Male measurements (in mm) (n=4). Body length 24.09–29.49 (27.70 ± 2.49); pronotum length 7.15–7.88 (7.44 ± 0.35); prozona length 3.79–4.08 (3.97 ± 0.13); metazona length 3.21–3.79 (3.46 ± 0.24); hind femur length 13.87–15.33 (14.78 ± 0.69) and tegmina length 7.30–11.68 (10.18 ± 2.03).
Female measurements (in mm) (n=5). Body length 32.70–41.46 (37.23 ± 3.85); pronotum length 10.07–12.84 (11.53 ± 1.15); prozona length 5.54–6.71 (6.1 ± 0.44); metazona length 4.52–6.27 (5.4 ± 0.74); hind femur length 18.39–20.14 (19.27 ± 0.67) and tegmina length 10.07–12.99 (11.18 ± 1.19).
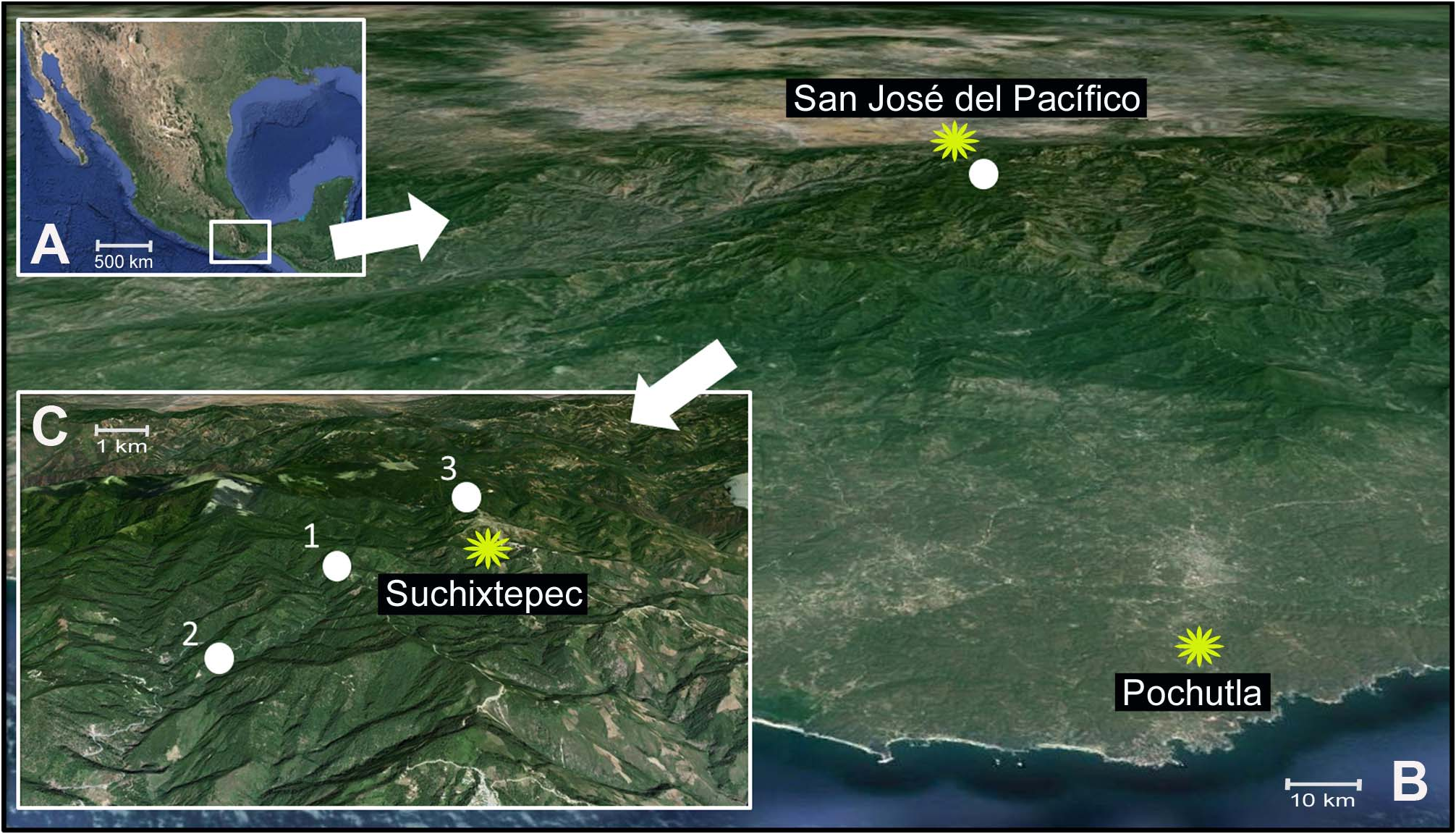
Holotype. Male. Mexico, Oaxaca, Near Suchixtepec. 26 km after San José del Pacífico. 16.0779500, - 96.4945833 ( WGS84 ). 2,321 m.a.s.l., edge of a pine forest. 10-XII-2011. ( Paolo Fontana, Ricardo Mariño-Pérez, Derek A. Woller, and Paola Tirello) ( Fig. 1 View FIGURE 1 B-1). GoogleMaps
Additional specimens examined. Eight paratypes ( three males and five females) with the same collecting information as the holotype ( Fig. 1 View FIGURE 1 B-1); two paratypes (females): Mexico, Oaxaca, On Km 165, near Suchixtepec. 29 km after San José del Pacífico. 16.0586944, -96.4996667 ( WGS84 ), 2127 m.a.s.l., pine-oak forest. 12-XII- 2013. ( Paolo Fontana, Ricardo Mariño-Pérez and Salomón Sanabria-Urbán) ( Fig. 1 View FIGURE 1 C-2) GoogleMaps ; two paratypes ( one male and one female: 1.3 km Northwest of Suchixtepec. 16.104083, -96.471722 ( WGS84 ). 2,663 m.a.s.l., pine-oak forest. 12-XII-2013. ( Paolo Fontana, Ricardo Mariño-Pérez and Salomón Sanabria-Urbán) ( Fig. 1 View FIGURE 1 C-3); two paratypes ( one male and one female: 6 km Northeast of Suchixtepec at Rio Molino, Sierra del Sur, 2,591 m.a.s.l., 5- V-1962. ( J. Stuart Rowley). Also, some nymphs were collected and examined from the holotype locality ( Fig. 1 View FIGURE 1 C- 1) and the locality 29 km after San José del Pacífico ( Fig. 1 View FIGURE 1 C-2). GoogleMaps
Type depository. Male holotype and five paratypes ( one male and four females) at UCFC, five paratypes ( two males and three females) at CNIN-UNAM, two paratypes ( one male and one female) at MNHN, and two paratypes ( one male and one female) at CPF .
Ecology. The habitat of the localities is in the boundaries of oak, pine-oak, and pine forests ( 1,900 –3,000 m.a.s.l.) ( Fig. 9C View FIGURE 9 ) within the southern parts of the Sierra Madre del Sur mountain range in Oaxaca, Mexico ( Fig. 1 View FIGURE 1 ). These vegetation types have been observed on diverse classes of rock: igneous, sedimentary and metamorphic, and do not tolerate draining deficiencies. Associated soil is of moderate acidity (pH 5.5–6.5) with abundant litter and organic matter in the superficial horizon and often in deeper horizons as well. Soil texture varies from clay to sand and the color is typically red, although sometimes it is also possible to find yellow, black, brown, or grey. According to the Köppen–Geiger climate classification system, the climate of the region in which these forests can be found is primarily Aw (equatorial, winter dry), but also: Am (equatorial, monsoonal), Bsh (arid, summer dry, hot arid), Cfa (warm temperate, fully humid, hot summer), Cfb (warm temperate, fully humid, warm summer), Cwa (warm temperate, winter dry, hot summer), and Cwb (warm temperate, winter dry, warm summer) ( García, 1973; Kottek et al., 2006). The mean precipitation per year ranges from 350 to over 2,000 mm, but is usually in the range of 600–1,200 mm. Temperatures vary from 10 to 26° C, but are quite often in the range of 12 to 20° C ( Rzedowski, 1981).
The forests are comprised of various species of oaks ( Quercus conspersa , Q. laeta , Q. laurina , Q. rugosa , and Q. ocoteifolia among others in addition to nunemrous species of pines ( Pinus ), hornbeams ( Carpinus ), Styrax , and Ternstroemia. All of these trees can range from 4 to 20 m in height, sometimes up to 30 m. The shrubby layer is comprised of the following genera: Bejaria , Comarostaphylis , Gaultheria , Lyonia , Litsea , Myrica , Calliandra , and Symplocos while herbaceous layer contains Salvia , Arenaria , Lobelia , and Lupinus . Climbing plants, epiphytes and rock-growers, are also quite common in these types of forests ( Rzedowski, 1981).
During both collecting expeditions (2011 and 2013) we found adults ( Figs. 2 View FIGURE 2 , 3 View FIGURE 3 , and 10), but also nymphs ( Figs. 4 View FIGURE 4 and 11 View FIGURE 11 ) representing almost all developmental stages ( Fig. 11 View FIGURE 11 ), indicating that this species persists, at least, until the end of January, throughout some of the coldest times of the year in Mexico, which suggests, based on its size and overlapping presence of almost all life stages, that this grasshopper has a lifespan lasting several months. This idea was seemingly confirmed by the fact that the two adult specimens discovered in MNHN were collected in May. According to multiple conversations with local residents, this grasshopper is abundant and easy to find throughout the area. Based on the currently known localities ( Fig. 1 View FIGURE 1 ), it is possible that the geographic range of this grasshopper is confined to the southern parts of the Sierra Madre del Sur mountain range ( Fig. 1C View FIGURE 1 ). Additionally, in 2013, a male and female were observed copulating in the typical manner of other Melanoplinae ( Otte, 1970) while the pair was hanging from a shrubby plant in the Lamiaceae family about two meters from ground level.
In 2011, both nymph and adult specimens were collected on a single (possible) host plant, Salvia elegans (Pineapple sage) ( Fig. 9A&B View FIGURE 9 ), which is also part of Lamiaceae , possesses showy red flowers, and is native to the pine-oak forests ( Fig. 9C View FIGURE 9 ) of Mexico and Guatemala. During the 2013 expedition we collected further specimens of the new species in the same kind of habitat, but on a wider range of plants, mostly members of Lamiaceae as well.
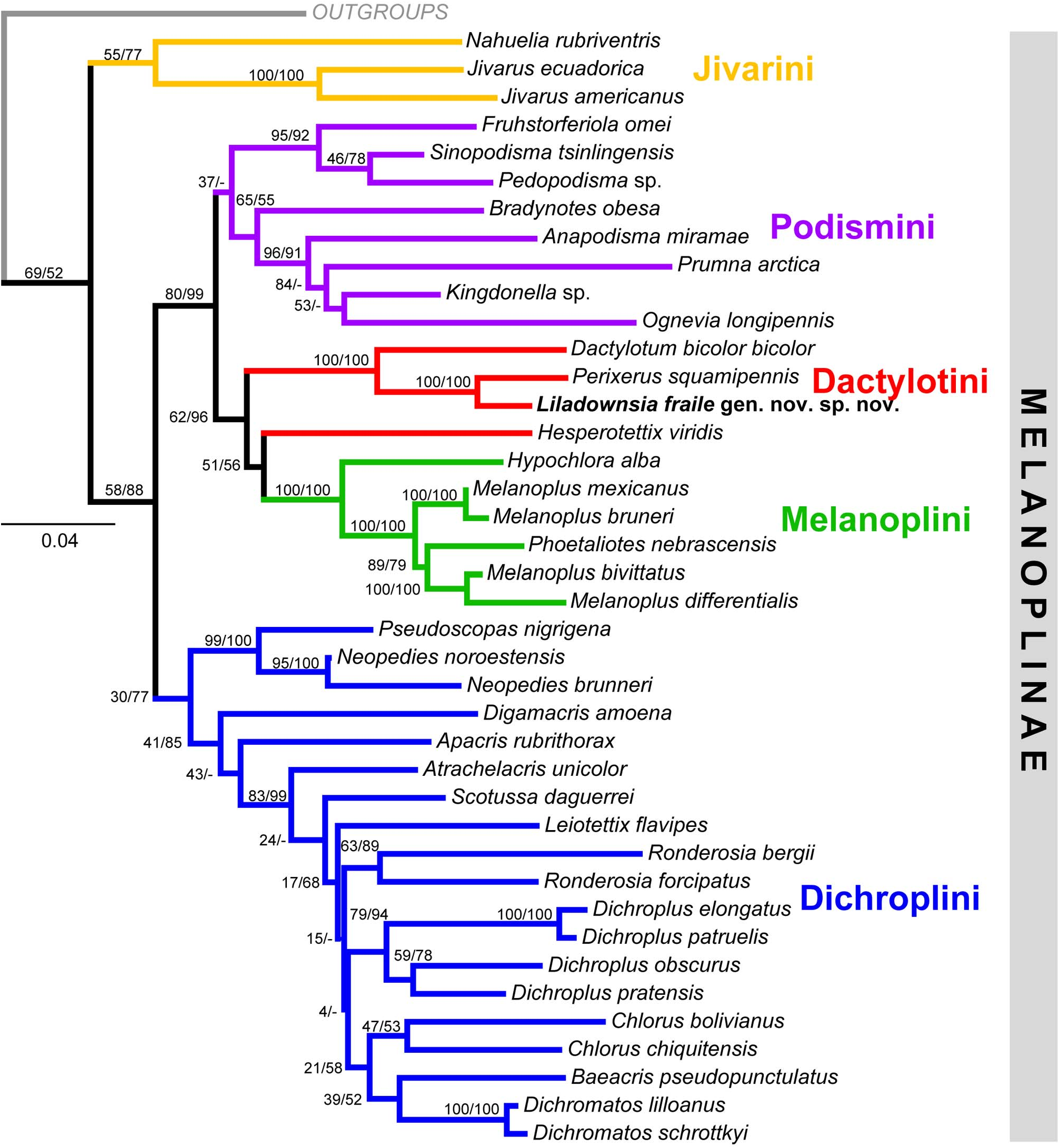
Phylogenetic analysis. Our matrix based on COI and COII consisted of 2,289 aligned nucleotides and 42 taxa. The PartitionFinder analysis found that the best-fit data-partitioning scheme was partitioning the alignment into two subsets by treating the first codon position of COI and the third codon position as a single partition with the remainders combined as the second partition. For the Bayesian analysis, PartitionFinder recommended GTR+G as the best model of nucleotide substitution for the first partition, and GTR+I+G for the second partition.
Both ML and Bayesian analyses recovered nearly identical topologies and we present the ML phylogram for discussion ( Fig. 12 View FIGURE 12 ). Nodal supports varied from weak to strong depending on the nodes. Monophyletic groups were recovered for the subfamily Melanoplinae overall as well as for four of the five included tribes: Jivarini , Podismini , Melanoplini , and Dichroplini . The analysis did not recover monophyly for Dactylotini because H. viridis was placed basally to Melanoplini . Additionally, the remaining three dactylotine taxa that were included, D. bicolor bicolor , P. squamipennis , and L. fraile , formed a clade and a sister relationship between P. squamipennis and L. fraile . was robustly recovered. Based on this phylogenetic relationship, as well as the previous findings ( Chapco 2006 and Chintauan-Marquier et al. 2011), we find that Hesperotettix does not belong to Dactylotini , but to Melanoplini . Thus, at the tribal level, the following relationships were recovered: (( Jivarini , (( Podismini , ( Dactylotini , Melanoplini )), Dichroplini )).








No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Melanoplinae |
|
Tribe |
Dactylotini |
|
Genus |