
Echinoderes yamasakii, Sørensen & Rohal & Thistle, 2018
|
publication ID |
https://doi.org/ 10.5852/ejt.2018.456 |
|
publication LSID |
lsid:zoobank.org:pub:DE1B1DEE-9871-4803-9F67-025F2B439560 |
|
DOI |
https://doi.org/10.5281/zenodo.3818844 |
|
persistent identifier |
https://treatment.plazi.org/id/000C173B-417F-4BE9-8EF5-75D584F3AF19 |
|
taxon LSID |
lsid:zoobank.org:act:000C173B-417F-4BE9-8EF5-75D584F3AF19 |
|
treatment provided by |
Valdenar |
|
scientific name |
Echinoderes yamasakii |
| status |
sp. nov. |
Echinoderes yamasakii sp. nov.
urn:lsid:zoobank.org:act:000C173B-417F-4BE9-8EF5-75D584F3AF19
Figs 21–23 View Fig View Fig View Fig , Tables 14–15
Diagnosis
Echinoderes with middorsal spines on segments 4 to 8 and spines in lateroventral positions on segments 6 to 9. Tubes present in lateroventral positions on segments 2 and 5, and in midlateral positions of segment 10. Very minute glandular cell outlets type 2 present in subdorsal positions on segment 2 and in laterodorsal positions on segments 8 and 9. Tergal plate of segment 11 terminates in conspicuously long, pointy, acicular tergal extensions, constituting 13 to 16% of trunk length. Males with three penile spines; females with relatively short and delicate lateral terminal accessory spines, and papillae in ventromedial positions of segment 7.
Etymology
The species is named after Hiroshi Yamasaki in recognition of his many contributions to kinorhynch taxonomy, morphology and phylogeny, and in particular his recent descriptions of the first deep-sea Echinoderes .
Material examined
Holotype
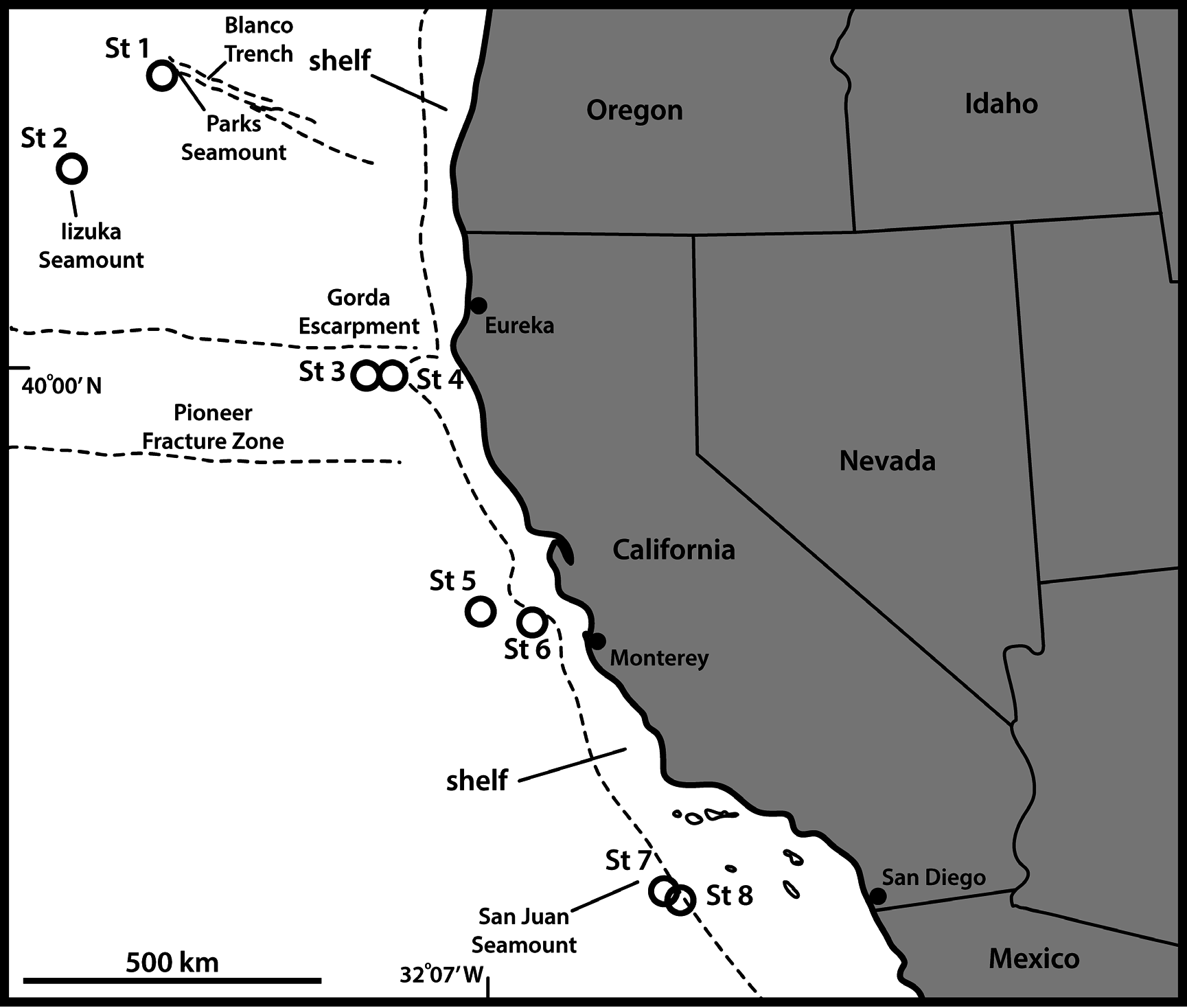
UNITED STATES OF AMERICA: ♀, US West Coast, California, off Monterey , 36°40′52″ N, 122°49′37″ W, St. 6, 2.719 m deep, from mud, 24 Sep. 2008, mounted in Fluoromount G on a glass slide (NHMD-226471). See Fig. 1 View Fig for localities and Table 1 View Table 1 for detailed station data.
GoogleMapsParatypes
UNITED STATES OF AMERICA: 3 ♀♀, 1 ♂, same collecting data as for holotype; 1 ♀, 3 ♂♂, St. 7; 2 ♀♀, 1 ♂, St. 8. All paratypes are mounted in Fluoromount G on glass slides (NHMD-226472–226482).
Additional non-type material
UNITED STATES OF AMERICA: 9 ♀♀, 12 ♂♂, St. 6; 1 ♀, St. 8. All are mounted for SEM and stored in the first author’s personal reference collection.
Description
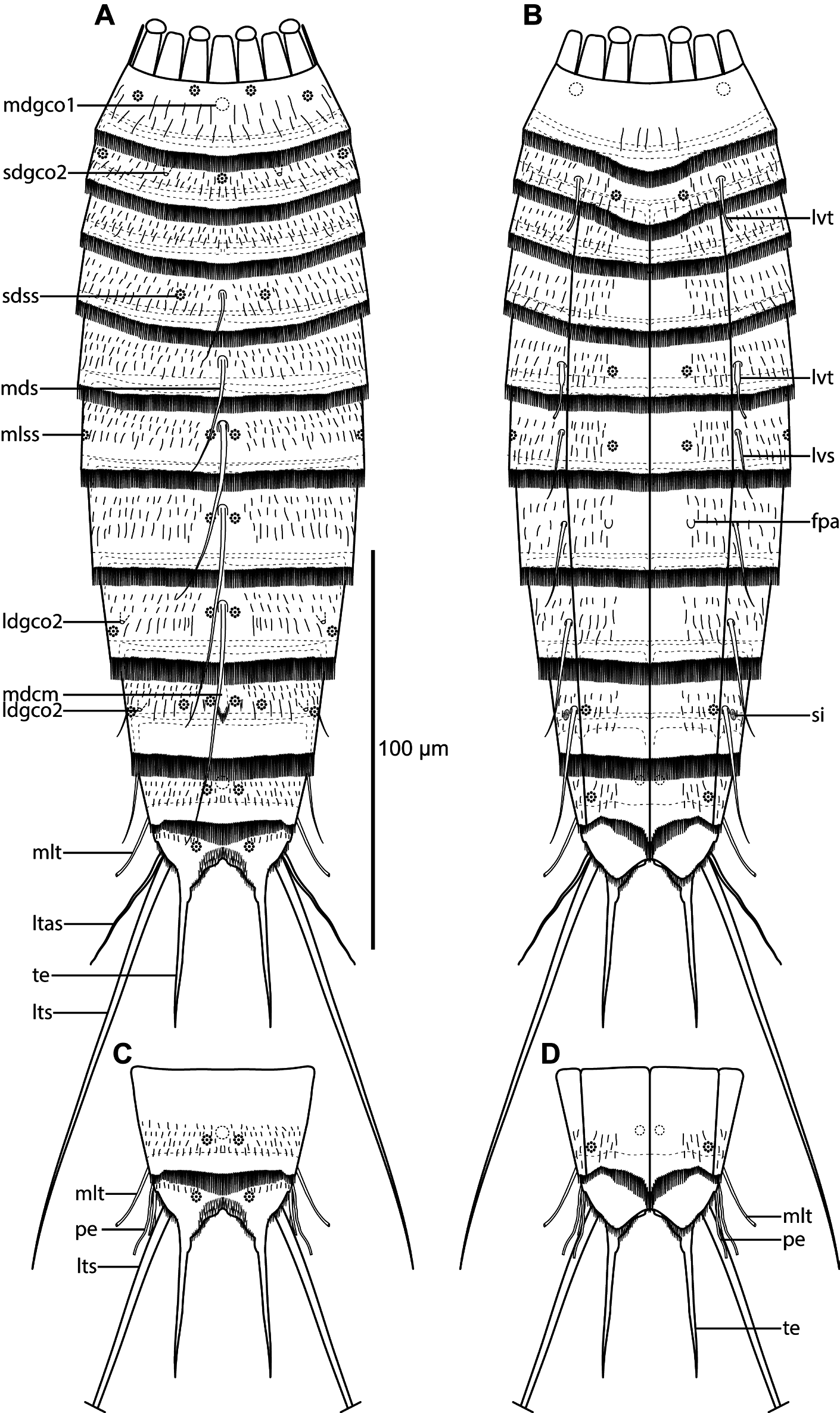
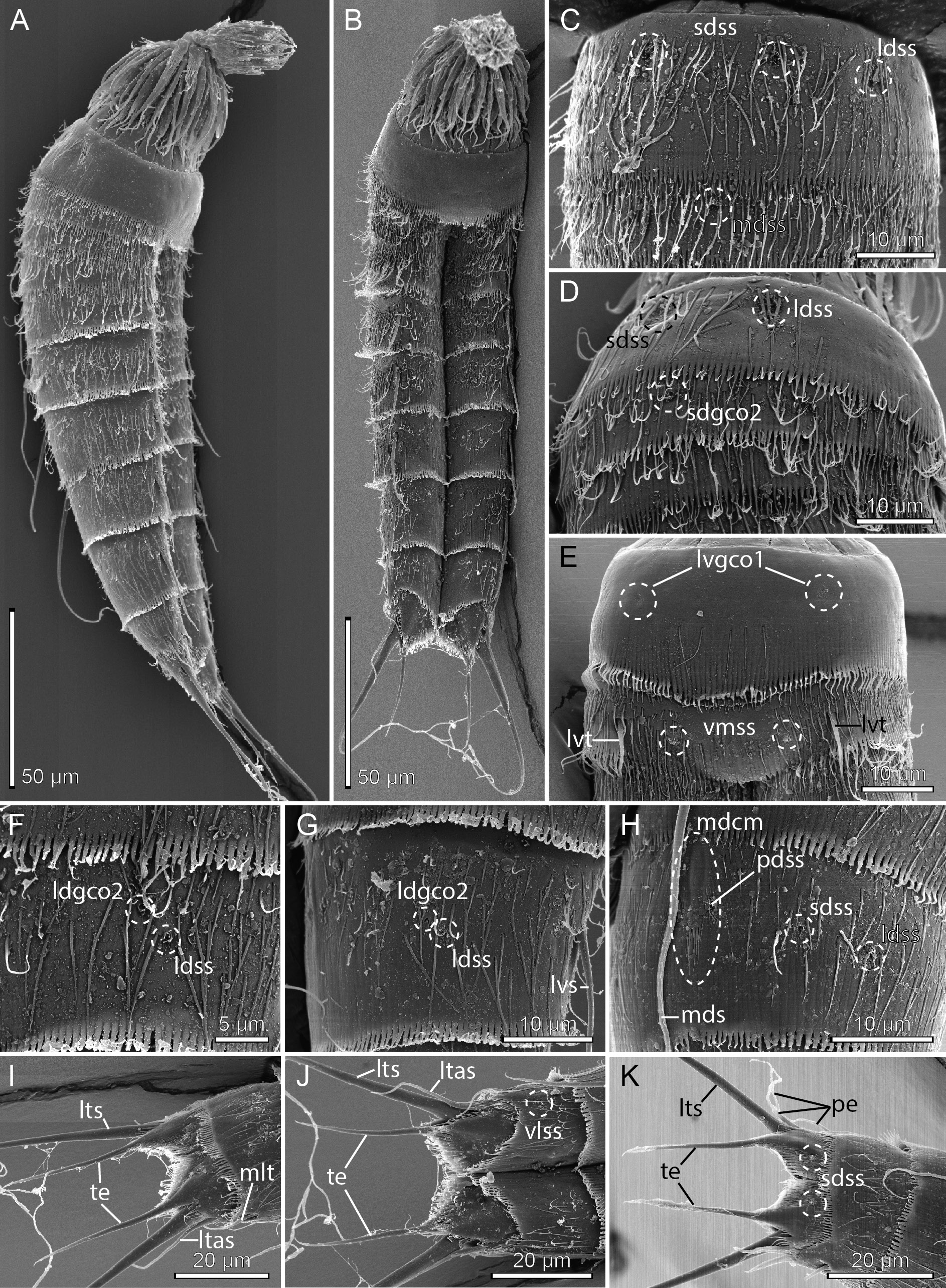
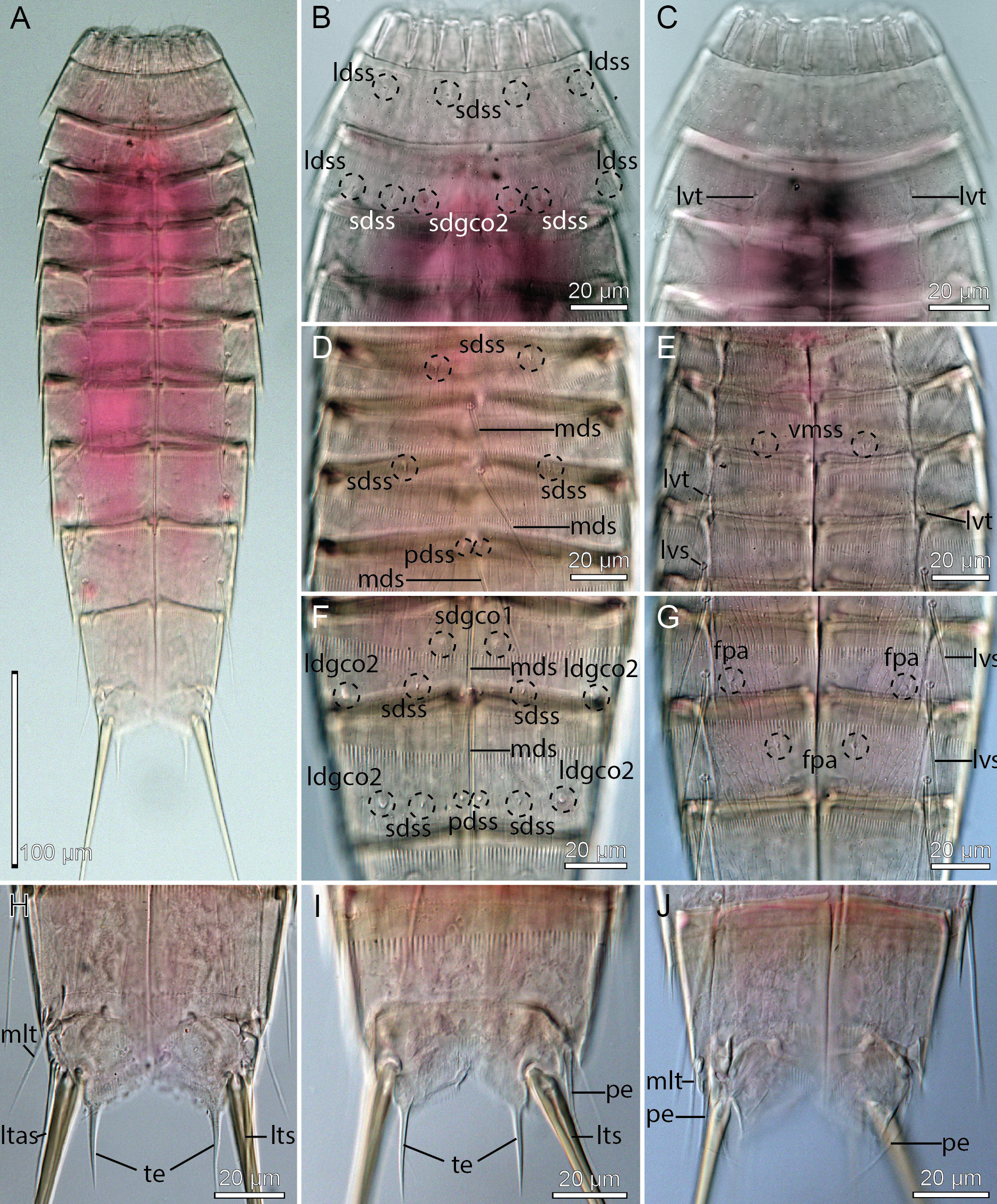
Adults with head, neck and eleven trunk segments ( Figs 21 View Fig A–B, 22A, 23A–B). The trunk is relatively short, terminating in a very large tergal extension. Lateral terminal spines are fairly short, but thick at their bases, from 40% to 57% of trunk length ( Figs 21 View Fig A–B, 22A), and lateral terminal accessory spines in females even shorter and less rigid ( Fig. 21 View Fig A–B). The cuticle appears very thin, which made it difficult to observe cuticular structures such as glandular cell outlets type 1 and sensory spots in LM. Observation of sensory spots was also obscured in SEM due to the dense hair covering and their very minute size. Hence, some glandular cell outlets type 1 and sensory spots might have been overlooked in the following description. For a complete overview of measurements and dimensions, see Table 14. Distribution of cuticular structures, i.e., sensory spots, glandular cell outlets, spines and tubes, is summarized in Table 15.
The head consists of a retractable mouth cone and an introvert. Inner oral styles are present, but their exact number and arrangement could not be determined. The external mouth cone armature consists of nine outer oral styles, each with four or five basal fringe tips. The introvert sectors are defined by ten primary spinoscalids in Ring 01. Each primary spinoscalid consists of a basal sheath and a distal end piece with a blunt tip. The basal sheaths have marginal extensions forming two layers of transverse fringes. End pieces are smooth. The arrangement of scalids follows the same pattern as in Echinoderes hviidarum sp. nov. described above; hence, see Fig. 11 View Fig for a complete overview. All spinoscalids consist of a basal sheath and a pointed end piece. The basal sheaths terminate in fringed margins in spinoscalids of Rings 02 and 04, whereas the sheaths of Ring 03 have a median spike only. The spinoscalids of Rings 05 to 07 are composed as those in the preceding ring, but with shorter end pieces.
The neck has 16 placids, measuring 13 µm in length, except for the slightly longer midventral ones, measuring 15 µm in length. The midventral placid is also broadest, measuring 13 µm in width at its base, whereas all others are narrower, measuring 7 µm in width at their bases. Four dorsal and two ventral trichoscalid plates are present; trichoscalid plates are fairly small and rounded to oval.
Segment 1 consists of a complete cuticular ring. Sensory spots are located in subdorsal and laterodorsal positions ( Figs 21A View Fig , 22B View Fig , 23 View Fig C–D); subdorsal ones are located very close to the anterior segment margin, whereas laterodorsal ones are slightly more posterior; sensory spots are small and rounded, with a few papillae. Glandular cell outlets type 1 are present in middorsal ( Fig. 22B View Fig ) and lateroventral ( Fig. 23E View Fig ) positions. Dorsal and lateral sides are densely covered with cuticular hairs, whereas the ventral side has a short midventral transverse row with 4–6 hairs only ( Fig. 23E View Fig ). The posterior segment margin is nearly straight, with a short ventral extension, terminating in a pectinate fringe with long and flexible fringe tips.
Segment 2 consists of a complete cuticular ring. The pachycyclus of the anterior segment margin is relatively thin and not interrupted. Very minute glandular cell outlets type 2 are present in subdorsal positions ( Figs 21A View Fig , 22B View Fig , 23D View Fig ) and well-developed tubes in lateroventral positions ( Figs 21B View Fig , 22C View Fig , 23E View Fig ); tubes are gradually tapered towards their tips, without distinct bases or lateral wings ( Fig. 23E View Fig ). Sensory spots are located in middorsal ( Fig. 23C View Fig ), laterodorsal (slightly more lateral than those on segment 1) and ventromedial positions ( Fig. 23E View Fig ). Glandular cell outlets type 1 not observed on this or any following segment, down to segment 9. Secondary pectinate fringe present near anterior segment margin of this and the following segments, but it is usually covered by the preceding segment. On this and all following segments, the cuticular hairs are bracteate, densely covering dorsal and lateral areas. Posterior segment margin as on preceding segment.
Segment 3, and the remaining segments, consist of one tergal and two sternal plates ( Figs 21B View Fig , 22A, C, E View Fig ). The pachycyclus of the anterior segment margin is well-developed and interrupted at the tergosternal and midsternal junctions and middorsally. No sensory spots were observed. Cuticular hairs densely cover the tergal plate and the lateral halves of the sternal plates. The posterior segment margin is nearly straight, with fringe tips as on preceding segment.
Segment 4 with a middorsal acicular spine ( Figs 21A View Fig , 22B View Fig ). Sensory spots are present in subdorsal positions. Pachycycli, pectinate fringe of posterior margin and cuticular hairs as on preceding segment.
Segment 5 with a middorsal acicular spine and well-developed tubes in lateroventral positions ( Figs 21 View Fig A–B); tubes have weakly developed, yet visible bases and lateral wings. Sensory spots present in ventromedial positions. Cuticular hairs, pachycycli and pectinate fringe of posterior margin as on preceding segment.
Segment 6 with acicular spines in middorsal and lateroventral positions ( Figs 21 View Fig A–B, 22E). Sensory spots present in paradorsal, midlateral and ventromedial positions. Cuticular hairs, pachycycli and pectinate fringe of posterior margin as on preceding segment.
Segment 7 with acicular spines in middorsal and lateroventral positions ( Figs 21 View Fig A–B, 22D–E). Sensory spots present in paradorsal positions. Females with papillary structures in ventromedial positions ( Figs 21B View Fig , 22E View Fig ); the structures are prominent in LM, but not visible in SEM. Cuticular hairs, pachycycli and pectinate fringe of posterior margin as on preceding segment.
Segment 8 with acicular spines in middorsal and lateroventral positions ( Figs 21 View Fig A–B, 22D–E). Very minute glandular cell outlets type 2 present in laterodorsal positions ( Figs 21A View Fig , 22D View Fig , 23F View Fig ). Sensory spots present in paradorsal and laterodorsal ( Fig. 23F View Fig ) positions. Cuticular hairs, pachycycli and pectinate fringe of posterior margin as on preceding segment.
Segment 9 with acicular spines in lateroventral positions ( Figs 21B View Fig , 22E View Fig ). Middorsal position with smooth area, flanked by conspicuous fringe-like extensions and a pair of paradorsal sensory spots (see mdcm on Fig. 23H View Fig ); the structure is visible in SEM only. Additional sensory spots present in subdorsal, laterodorsal ( Fig. 23 View Fig G–H) and ventrolateral positions. Very minute glandular cell outlets type 2 present in laterodorsal positions (21A, 22D, 23G). Small rounded sieve plates are present in lateral accessory positions ( Figs 21B View Fig , 22E View Fig ). Cuticular hair covering on tergal plate less dense than those on preceding segments. Pachycycli and pectinate fringe of posterior margin as on preceding segment.
Segment 10 with tubes in midlateral positions ( Figs 21 View Fig , 22F, H View Fig , 23I View Fig ); tubes appear fairly stout and are much easier to observe in LM ( Fig. 22F, H View Fig ) than tubes on segment 10 usually are in most other species of Echinoderes ; differentiated bases and lateral wings apparently not present. Sensory spots are present in paradorsal and ventrolateral ( Fig. 23J View Fig ) positions. A single glandular cell outlet type 1 was observed with SEM in middorsal position and a pair in paraventral positions. The cuticular hairs are scattered over the tergal plate and lateral parts of the sternal plates, but not as densely as on the preceding segments. The posterior segment margin of the tergal plate is straight, whereas the margins of the sternal plates are broadly concave, with shorter fringe tip around the tergosternal junctions and longer tips towards the paraventral positions. Pachycycli as on preceding segment.
Segment 11 with medium to short lateral terminal spines ( Figs 21 View Fig A–B, 22A). Males with three pairs of penile spines ( Figs 21 View Fig C–D, 22H, 23K); dorsal and ventral penile spines are thin, but also rather long and flexible tubes, whereas the median one is slightly thicker. Females with short and relatively flexible lateral terminal accessory spines ( Figs 21 View Fig A–B, 22G, 23I–J). Sensory spots present in subdorsal positions ( Fig. 23K View Fig ). The tergal plate has a few short cuticular hairs on the most anterior part, but otherwise the segment is completely devoid of hairs. Long hair-like extensions are present in the mid- to subdorsal areas. Tergal extensions are very long, extending into stout spinous tips ( Figs 21 View Fig , 22 View Fig G–H, 23I–K); tergal extensions measure 33 to 35 µm, equal to 13 to 16% of trunk length. Sternal extensions shorter and rounded.
Remarks for Echinoderes lupherorum sp. nov. and E. yamasakii sp. nov.
Echinoderes lupherorum sp. nov. and E. yamasakii sp. nov. can easily be distinguished from most other congeners by their long and acicular tergal extensions of segment 11. Only nine other species of Echinoderes have conspicuous spinous tergal extensions like this. These include E. microaperturus Sørensen et al., 2012 , E. spinifurca Sørensen et al., 2005 , E. sylviae Landers & Sørensen, 2018 , E. cernunnos , E. multiporus Yamasaki et al., 2018 , E. higginsi Huys & Coomans, 1989 , E. rhaegali Grzelak & Sørensen, 2017 , E. arlis and E. filispinosus Adrianov, 1989 (see Adrianov & Malakhov 1999; Huys & Coomans 1989; Sørensen et al. 2005, 2012; Grzelak & Sørensen 2018, in press; Yamasaki et al. 2018b; Landers & Sørensen in press). Of these, the latter have no middorsal spines at all, and E. multiporus , E. higginsi , E. rhaegali and E. arlis have middorsal spines on segments 4, 6 and 8 only opposed to middorsal spines on segments 4 to 8 in E. lupherorum sp. nov. and E. yamasakii sp. nov. Furthermore, E. cernunnos has a rather special appearance, with large glandular cell outlets type 2 on segments 5 and 7, four pairs of glandular cell outlets type 2 on segment 2, and the tergal plate of segment 11 divided into two ( Sørensen et al. 2012); hence, this species can hardly be confused with any of those mentioned here.
This narrows us down to five rather similar and putatively closely related species, that besides the two new ones include E. microaperturus , E. spinifurca and E. sylviae . Besides the spinous and acicular tergal extensions, these species share nearly the same spine/tube formula (the exact position of the tubes on segment 2 appears to vary between the species, between being either lateroventral or ventrolateral). All five species have the tubes on segment 10 being relocated from the common laterodorsal positions to the rarer midlateral positions (erroneously referred to as laterodorsal in the description of E. spinifurca , but illustrated in midlateral positions). Furthermore, all of them appear to have female papillae on the sternal plates of segments 6, 7 and/or 8, three of them ( E. microaperturus , E. lupherorum sp. nov. and E. yamasakii sp. nov.) have minute glandular cell outlets type 2 in subdorsal positions on segment 2, and four of them (the same three and E. sylviae ) in laterodorsal positions of segments 8 and 9 as well ( Sørensen et al. 2005, 2012; Landers & Sørensen in press). These characters, together with the general overall similarity between the species, suggest that they might represent a group of closely related species within Echinoderes , similar to the Echinoderes coulli group (see, e.g., Sørensen 2014, Yamasaki 2016b). Hence, for now we will refer to E. lupherorum sp. nov., E. yamasakii sp. nov., E. microaperturus , E. spinifurca and E. sylviae as the Echinoderes spinifurca group. Landers & Sørensen (in press) recently pointed out how the tergal extensions in E. microaperturus , E. spinifurca and E. sylviae differ from spinous tergal extensions in other species. In congeners with spinous tergal extensions, the pointed processes are a result of gradually tapering extensions from the tergal plate ( Landers & Sørensen 2018). This is opposite to the acicular tergal extensions in species of the Echinoderes spinifurca group, where the extensions from the tergal plate narrow abruptly around the bases of the terminal needles (see, e.g., Figs 19 View Fig H–I, 20J, 22F–H, 23I–K).
Within the Echinoderes spinifurca group, the species are most easily distinguished by the length of their tergal extensions (absolute length as well as tergal extensions relative to trunk length), and the presence of glandular cell outlets type 2 on segment 2. Echinoderes yamasakii sp. nov. stands out as the species with the longest tergal extensions (34 µm on average and 14.3% of trunk length). The conspicuously long tergal extensions make it easy to distinguish it from the species with the second longest extensions, E. spinifurca . Landers & Sørensen (in press) showed that the length of the tergal extensions in E. spinifurca ranges from 25 µm on average in the Atlantic Fort Pierce population, to 28 µm on average in the slightly longer populations of the Gulf of Mexico, resulting in tergal extension vs trunk length ratios (TE/TL in the following) of 9.0% and 12.4%, respectively. E. spinifurca furthermore lacks the glandular cell outlets type 2 ( Sørensen et al. 2005; Sørensen & Landers in press) that are present in E. yamasakii sp. nov.
The remaining three species have shorter tergal extensions: Echinoderes lupherorum sp. nov. – 26 µm on average, but only a TE/TL ratio of 7.2% due to their relatively large trunk sizes; E. sylviae – 16 µm on average and a TE/TL ratio of 6.6% (see Landers & Sørensen in press); and E. microaperturus – 14 µm on average and a TE/TL ratio of 5.0% (measurements made on type material deposited at the NHMD). This puts the three species in a group with relatively low TE/TL ratio, which make their general appearance very similar. Echinoderes sylviae is most easily discriminated by its lack of subdorsal glandular cell outlets type 2 on segment 2. E. lupherorum sp. nov. and E. microaperturus are indeed very similar. They differ slightly in their distribution of sensory spots, but the most straightforward way to distinguish the species is by the relative length of their middorsal spines. Echinoderes microaperturus has conspicuously short middorsal spines that hardly extend beyond the posterior margins of the segments they attach to. By contrast, E. lupherorum sp. nov. has fairly long middorsal spines that always extend beyond the segment margins, and often beyond the posterior margin of the following segment as well.


| US |
University of Stellenbosch |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |