
Fissuroderes higginsi Neuhaus & Blasche, 2006
|
publication ID |
https://doi.org/ 10.5852/ejt.2018.456 |
|
publication LSID |
lsid:zoobank.org:pub:DE1B1DEE-9871-4803-9F67-025F2B439560 |
|
DOI |
https://doi.org/10.5281/zenodo.3818836 |
|
persistent identifier |
https://treatment.plazi.org/id/F64287A2-5017-FFDC-17FA-F8D3FBB409E3 |
|
treatment provided by |
Valdenar |
|
scientific name |
Fissuroderes higginsi Neuhaus & Blasche, 2006 |
| status |
|
Fissuroderes higginsi Neuhaus & Blasche, 2006
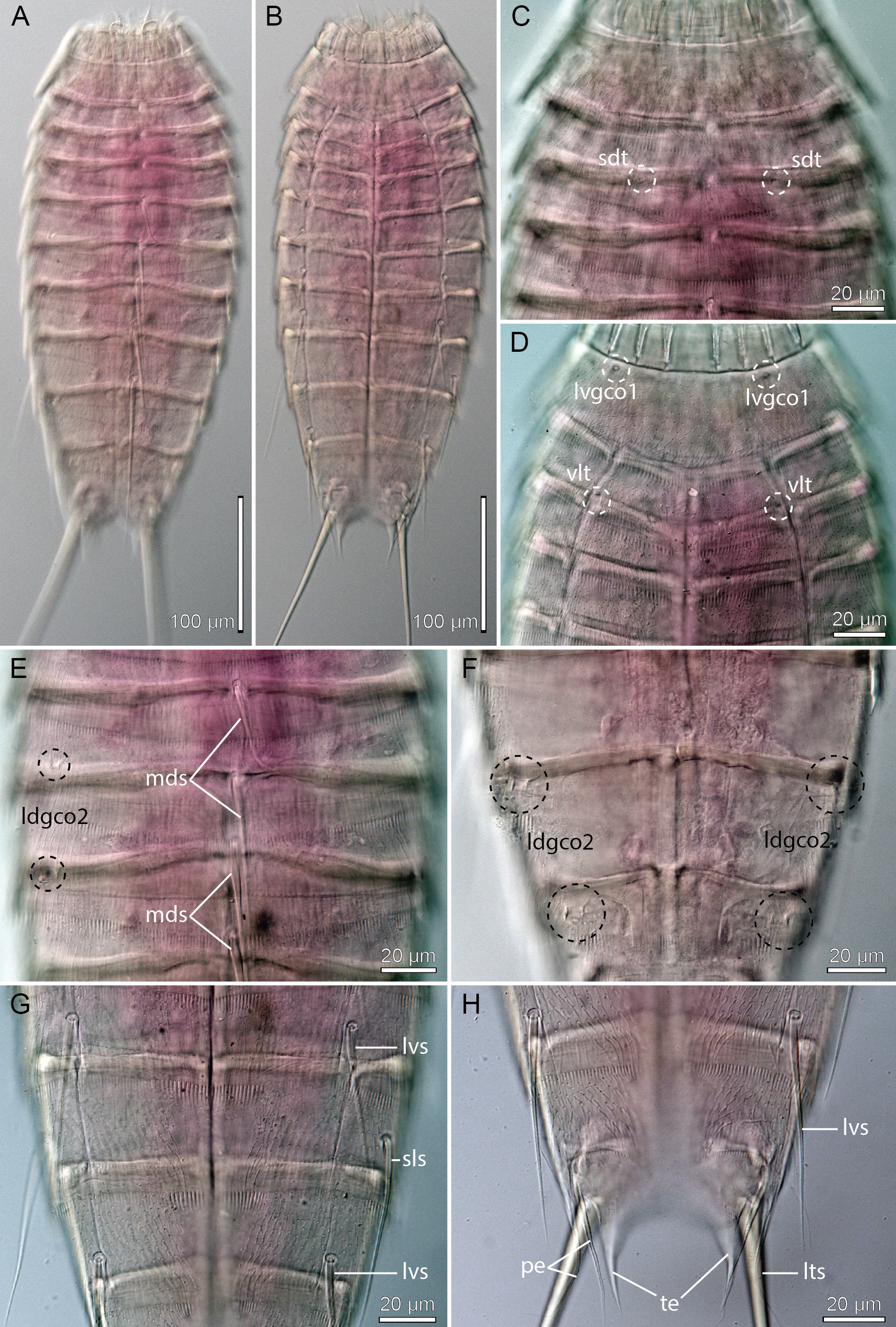
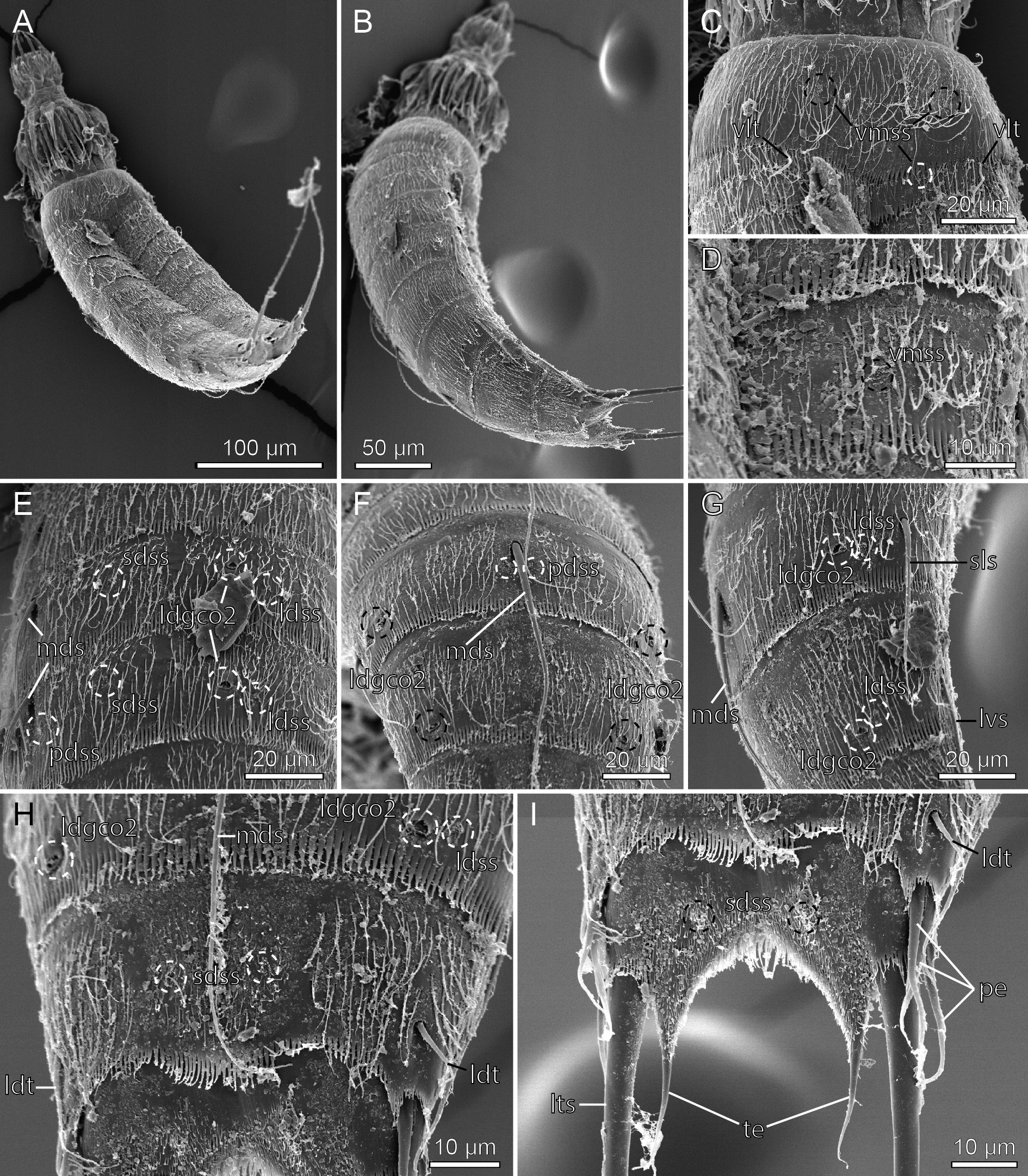
Figs 26–27 View Fig View Fig , Table 18 View Table 18
Material examined
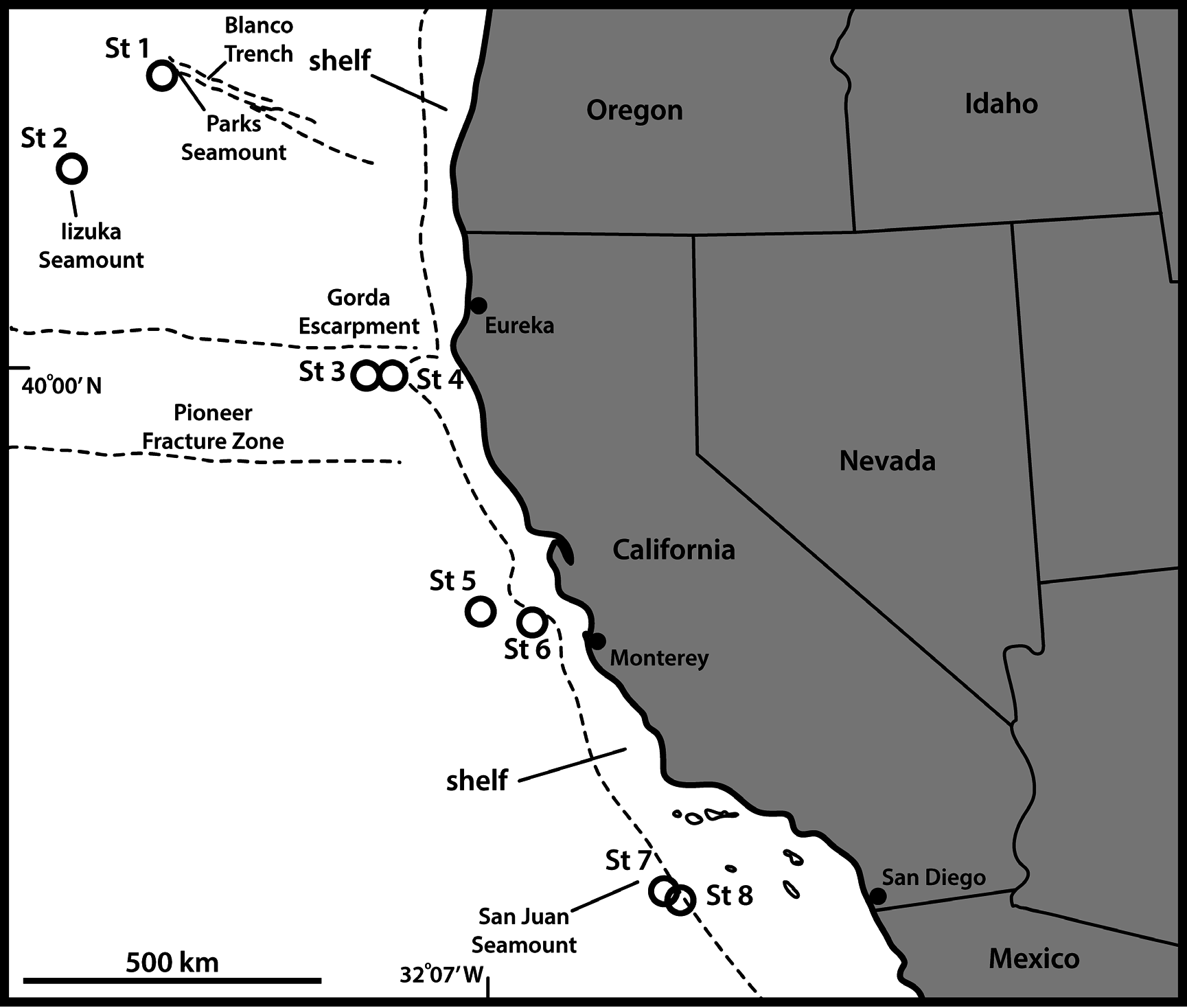
UNITED STATES OF AMERICA: 1 adult ♂, St. 3, mounted in Fluoromount G (NHMD-227025). See Fig. 1 View Fig for localities and Table 1 View Table 1 for detailed station data.
Other material
UNITED STATES OF AMERICA: 1 adult ♂, St. 7, mounted for SEM and stored in the first author’s personal reference collection.
Notes on morphology and comparison with type material
A summary of the distribution of cuticular structures, i.e., sensory spots, glandular cell outlets, spines and tubes is provided in Table 18 View Table 18 .
Fissuroderes higginsi was described from New Zealand, from depths ranging from 40 to 885 m ( Neuhaus & Blasche 2006). The species is highly characteristic by its displacement of the lateral spine on segment 8 to a sublateral position (note that Neuhaus & Blasche (2006) refer to the spines as lateral accessory, but line art and LM images show it in a sublateral position), and glandular cell outlets type 2 arranged in a laterodorsal series on segments 5, 6, 8 and 9.
The material examined for the present study yielded two specimens ( Figs 26–27 View Fig View Fig ) that are nearly identical to F. higginsi as described by Neuhaus & Blasche (2006). The meristic data (segment and spine lengths) of the single specimen mounted for LM were all about 10–20% larger than those of the type material, but segment to spine length dimensions were the same, including the conspicuously long middorsal spine on segment 8. Morphologically, the recorded specimens were nearly identical with the type specimens. The only noted differences were 1) a pair of subdorsal sensory spots on segment 3, reported from the type material, but not visible in the specimens from the present study, and 2) the dorsal tubes on segment 2 that are laterodorsal in the type specimens, but occur in a more subdorsal position in the Californian specimens ( Fig. 26C View Fig ). These minor differences can be neglected as either uncertainties (the sensory spots) or intraspecific variation (the tubes), and we therefore feel confident that the recorded specimens are F. higginsi .
There is obviously a considerable depth range between the New Zealand (40 to 885 m depth) and the Californian populations (3675–3853 m depth), but we see a similar trend in the depth preferences of E. hakaiensis , which again stresses that depth might play a minor role for species adapted to live on the shelf and below the sublittoral zone. Again, the species has a surprisingly large distribution. It might not be as extreme as was the case for E. unispinosus , but there is still a considerable distance between the type locality in New Zealand and the Californian deep-sea. This could support that deep-sea kinorhynchs actually show greater distributional ranges than species from shallower habitats.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |