
Acanthochondria cyclopsetta Pearse, 1952
|
publication ID |
https://doi.org/ 10.5281/zenodo.198947 |
|
DOI |
https://doi.org/10.5281/zenodo.6198871 |
|
persistent identifier |
https://treatment.plazi.org/id/E91687DD-3873-FFF4-5990-8C06FE5C53CA |
|
treatment provided by |
Plazi |
|
scientific name |
Acanthochondria cyclopsetta Pearse, 1952 |
| status |
|
Acanthochondria cyclopsetta Pearse, 1952
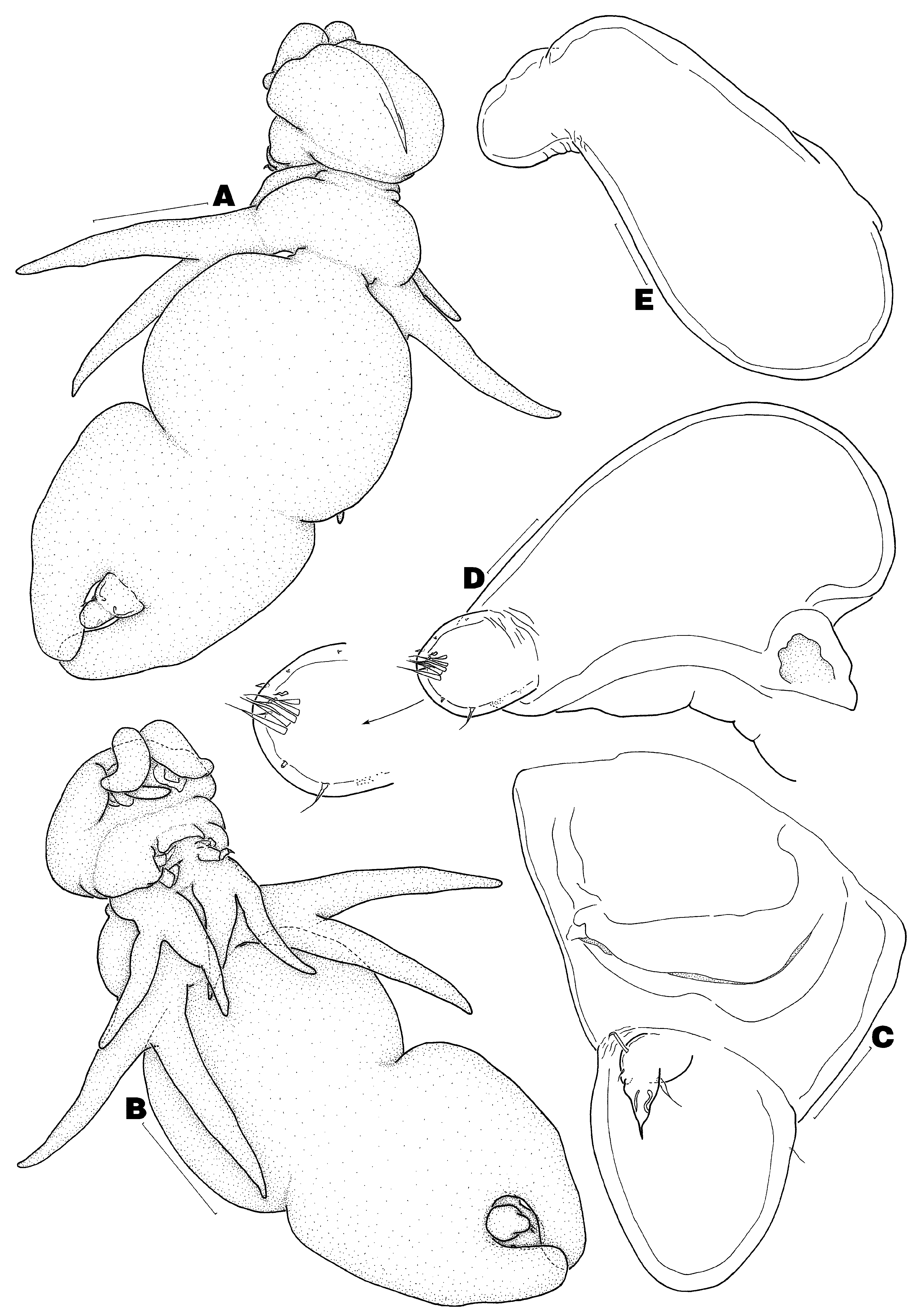
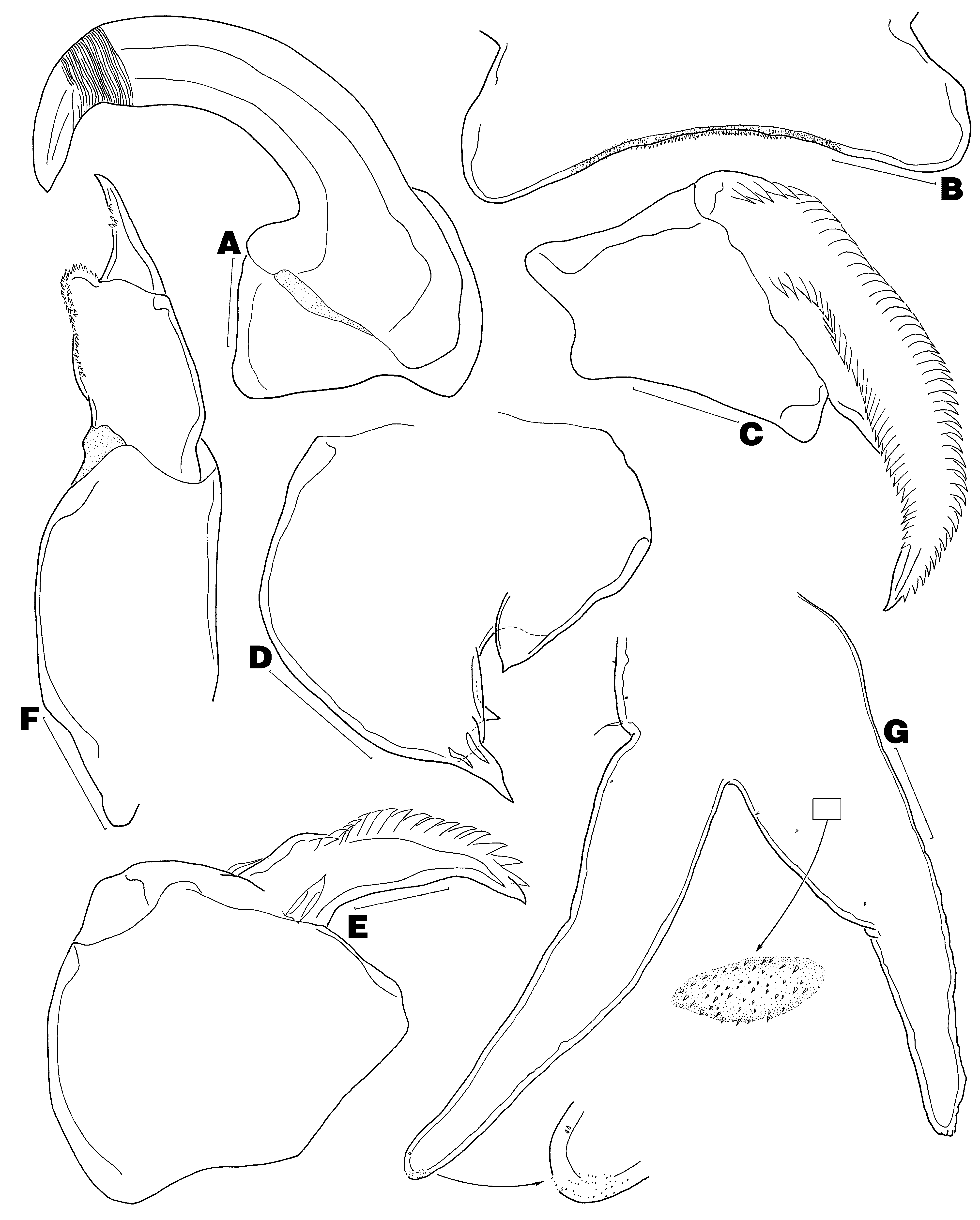
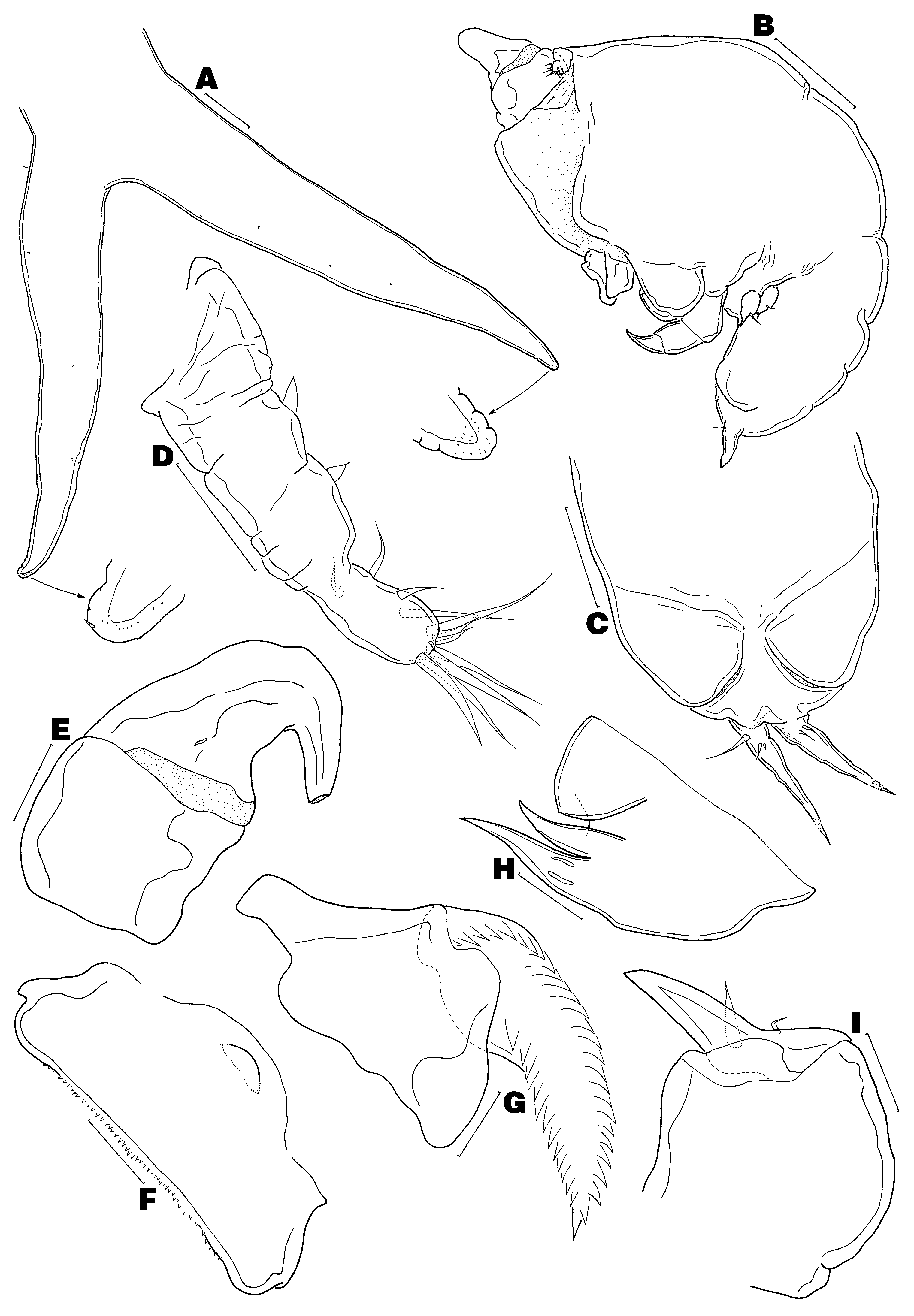
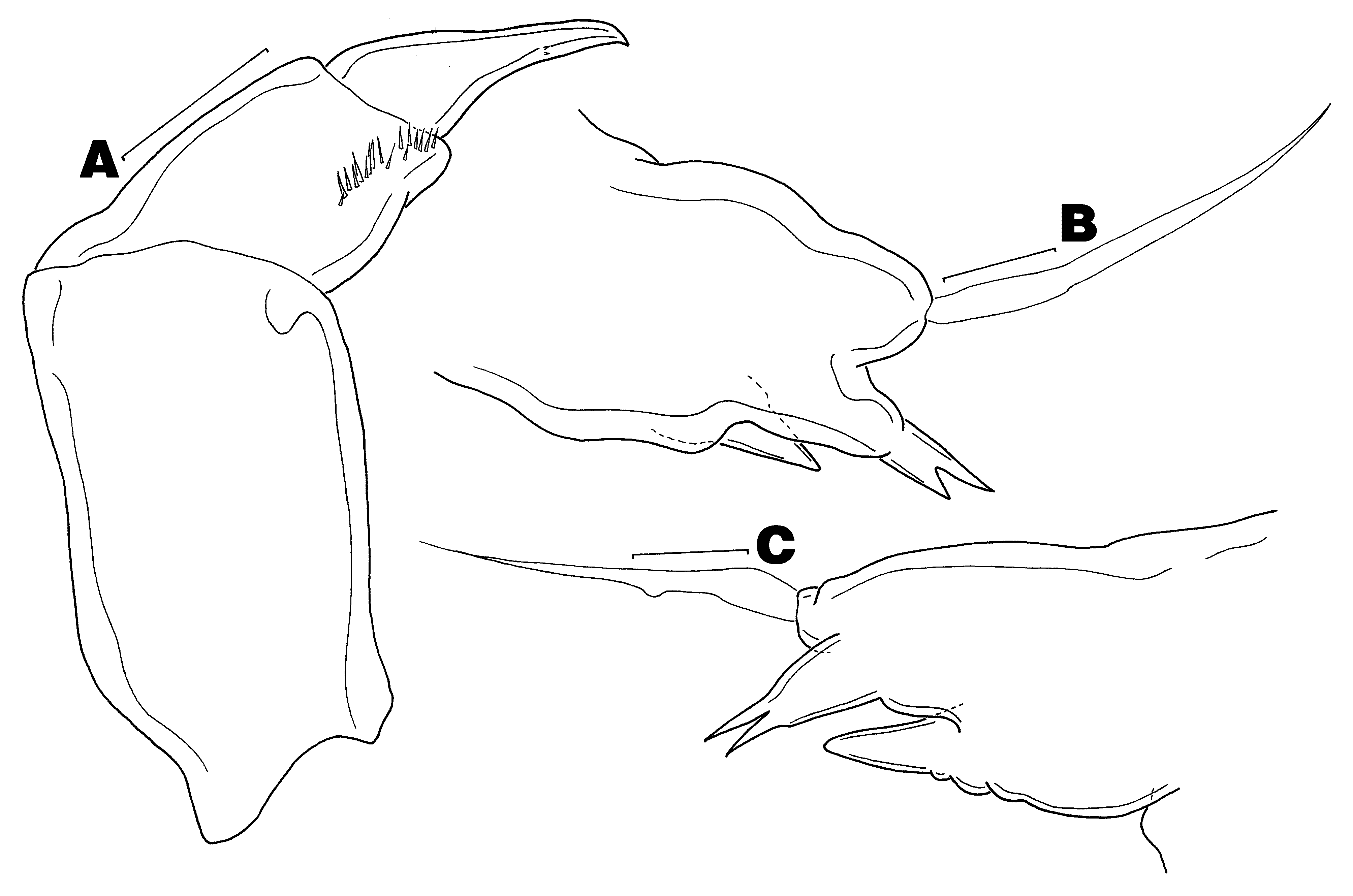
( Figs 1–4 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 )
Material examined. 3 Ψ and 3 ɗ [1 Ψ with attached ɗ, 1 Ψ with 2 attached ɗ, and 1 Ψ without attached ɗ] ( CASIZ 180282), ex branchial cavity wall of 1 Cyclopsetta fimbriata (Goode & Bean) (170.2 mm SL; CAS Fish Catalogue No. SU 49886), from the northwest Atlantic Ocean (32°26΄N, 79°13΄W), off the coast of South Carolina, U.S.A., 4 February, 1940.
Redescription of adult female. Body ( Figs 1 View FIGURE 1 A–B) divided into head, short neck, and large trunk. Total body length (from anterior margin of head to distal end of posterior processes on trunk) 5.67 mm; trunk width 2.33 mm (n = 1). Head composed of cephalosome only, about as long as wide. Neck region composed of first and second pedigers, former proportionately smaller than latter. Trunk composed of pedigers 3 and 4, delimited anteriorly from neck by transverse constriction, with pronounced mid-lateral constriction and pair of posterolateral processes; latter about 1.70 times as long as genito-abdomen. Genito-abdomen ( Figs 1 View FIGURE 1 A–C) divisible as 2 tagmata by transverse constriction; anterior tagma (genital somite) about as long as posterior tagma (abdominal somite), former with ventral pair of genital apertures (usual mid-ventral pair of setae not observed); posterior tagma suboval, slightly narrower than genital somite, with dorsal pair of sensilla and anteroventral pair of caudal rami. Caudal ramus ( Fig. 1 View FIGURE 1 C) spiniform, armed with 2 ventral setae, 1 dorsal seta, and 1 inner knob.
Antennule ( Figs 1 View FIGURE 1 D–E) unsegmented, distinctly separated into large fleshy base and narrower tip (Type B- V); latter ornamented with 2 patches of minute spinules along ventral margin and armed with 13 elements in total. Antenna ( Fig. 2 View FIGURE 2 A) 2-segmented, composed of coxobasis and 1-segmented endopod; coxobasis short, unarmed; endopod forming powerful uncinate claw bearing transverse surface striations near tip. Labrum ( Fig. 2 View FIGURE 2 B) with row of spinules along posterior margin. Mandible ( Fig. 2 View FIGURE 2 C) 1-segmented, with apical falcate blade armed with 46 teeth along convex margin and 33 teeth along concave margin (n = 1). Paragnath not observed. Maxillule ( Fig. 2 View FIGURE 2 D) unilobate, with 2 unequal apical elements and inner knob bearing acuate tip. Maxilla ( Fig. 2 View FIGURE 2 E) 2-segmented, composed of syncoxa and basis; syncoxa robust, unarmed; basis forming claw-like process, armed with 2 unequal basal setae and 17 marginal teeth (n = 1). Maxilliped ( Fig. 2 View FIGURE 2 F) 3- segmented, composed of elongate syncoxa, robust basis and short terminal claw (endopod); syncoxa naked; basis with 2 patches of denticles along inner margin; claw with multiple accessory teeth. Leg 1 ( Fig. 2 View FIGURE 2 G) rami relatively slender and elongate; surface of protopod and rami ornamented with several spiniform setules and numerous bullate patches each furnished with minute spinules (only 1 bullate structure drawn in Fig. 2 View FIGURE 2 G); exopod with 2 vestigial setal elements and patch of minute spinules apically. Leg 2 ( Fig. 3 View FIGURE 3 A) similar to leg 1, except slightly larger in size, exopod with only 1 reduced apical seta, and endopod with additional distal spinule patch.
Description of adult male. Body ( Fig. 3 View FIGURE 3 B) 0.40–0.42 mm long (n = 2), divided into globose cephalothorax and ventrally flexed genito-abdomen; latter with pair of unarmed opercula, each covering genital aperture ( Fig. 3 View FIGURE 3 C). Caudal ramus ( Fig. 3 View FIGURE 3 C) spiniform, bears 3–4 basal setae and minute spinules distally (usual small inner knob not observed).
Antennule ( Fig. 3 View FIGURE 3 D) unmodified, wrinkled, with armature of 1-1-2-2-8. Antenna ( Fig. 3 View FIGURE 3 E) short and stout, with both segments unarmed (tip broken off in illustration). Labrum ( Fig. 3 View FIGURE 3 F) with anteromedian knob. Mandibular blade ( Fig. 3 View FIGURE 3 G) with 26 teeth on convex side and 13 teeth on concave side (n = 1). Maxillule ( Fig. View FIGURE 3
3H) similar to that of female. Maxilla ( Fig. 3 View FIGURE 3 I) as in female, except without teeth on basis. Maxilliped ( Fig. 4 View FIGURE 4 A) as in female, except with fewer spinules on basis and fewer accessory teeth on claw. Legs 1 and 2 ( Figs 4 View FIGURE 4 B–C) rudimentary, each with long outer subapical seta, distal lobe bearing apical bifurcate element, and inner conical process.
Remarks. We attribute our adult female specimens of the Chondracanthidae Milne Edwards, 1840 from Cyclopsetta fimbriata (Goode & Bean) as Acanthochondria cyclopsetta Pearse, 1952 based on similarities in the relative size of the posterolateral processes of the trunk (each process is more than 1.50 times as long as the genitoabdomen), shape and size of the abdomen (this somite is ovate and nearly as long as the genital somite), insertion of the caudal rami (each ramus is inserted at the proximal end of the abdomen), and shape of legs 1 and 2 (the rami of both legs are relatively slim and highly elongate). The shape of the head, trunk, and antennule of our specimens could not be accurately compared with those of the holotype since these features were slightly distorted by coverslip compression in the latter material. Similarly, the fine details of the head appendages and ornamentation of both leg pairs could not be precisely compared as the holotype is poorly preserved. Despite these shortcomings, we are confident our specimens are conspecific with A. cyclopsetta considering the former were also collected from the western Atlantic Ocean and from the same host genus, i.e. Cyclopsetta Gill, as the latter; both host species have overlapping geographic distributions (see Khidir et al. 2004); and our specimens share salient morphological features with A. cyclopsetta as noted above. Collection of new A. cyclopsetta specimens from the type host is, however, still warranted (we must add that we did not recover any specimens of A. cyclopsetta from four Cyclopsetta chittendeni Bean specimens housed in the Ichthyology Department at LACM). For distinguishing features of A. cyclopsetta , see the Remarks section of the following taxon.
| CAS |
California Academy of Sciences |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |