
Orbiniellajamesi, Georgieva & Wiklund & Ramos & Neal & Glasby & Gunton, 2023
|
publication ID |
https://doi.org/ 10.3853/j.2201-4349.75.2023.1800 |
|
publication LSID |
lsid:zoobank.org:pub:32014E75-6253-41C0-BEDC-7A461321A0A1 |
|
DOI |
https://doi.org/10.5281/zenodo.10997908 |
|
persistent identifier |
https://treatment.plazi.org/id/BECCFCE3-8F48-4E0A-B9D4-A1547389E69E |
|
taxon LSID |
lsid:zoobank.org:act:BECCFCE3-8F48-4E0A-B9D4-A1547389E69E |
|
treatment provided by |
Felipe |
|
scientific name |
Orbiniellajamesi |
| status |
sp. nov. |
Orbiniellajamesi View in CoL sp. nov.
urn:lsid:zoobank.org:act:BECCFCE3-8F48-4E0A-B9D4-A1547389E69E
Fig. 18 View Figure 18
Holotype: AMW.53705, IN2017_ V03 _100; 9 June 2017; off Byron Bay , NSW, Australia, beam trawl, start: 28.05°S 154.08°E, 999 m, end: 28.10°S 154.08°E, 1013 m GoogleMaps . Paratype: AM W.52199, same locality as holotype GoogleMaps . Other specimens examined: NHMUKANEA 2023.1201 – 1202 About NHMUK , same locality as holotype. GoogleMaps DNA vouchers: AMW.53705 ( COI) GoogleMaps , AM W.52199 (16 S).
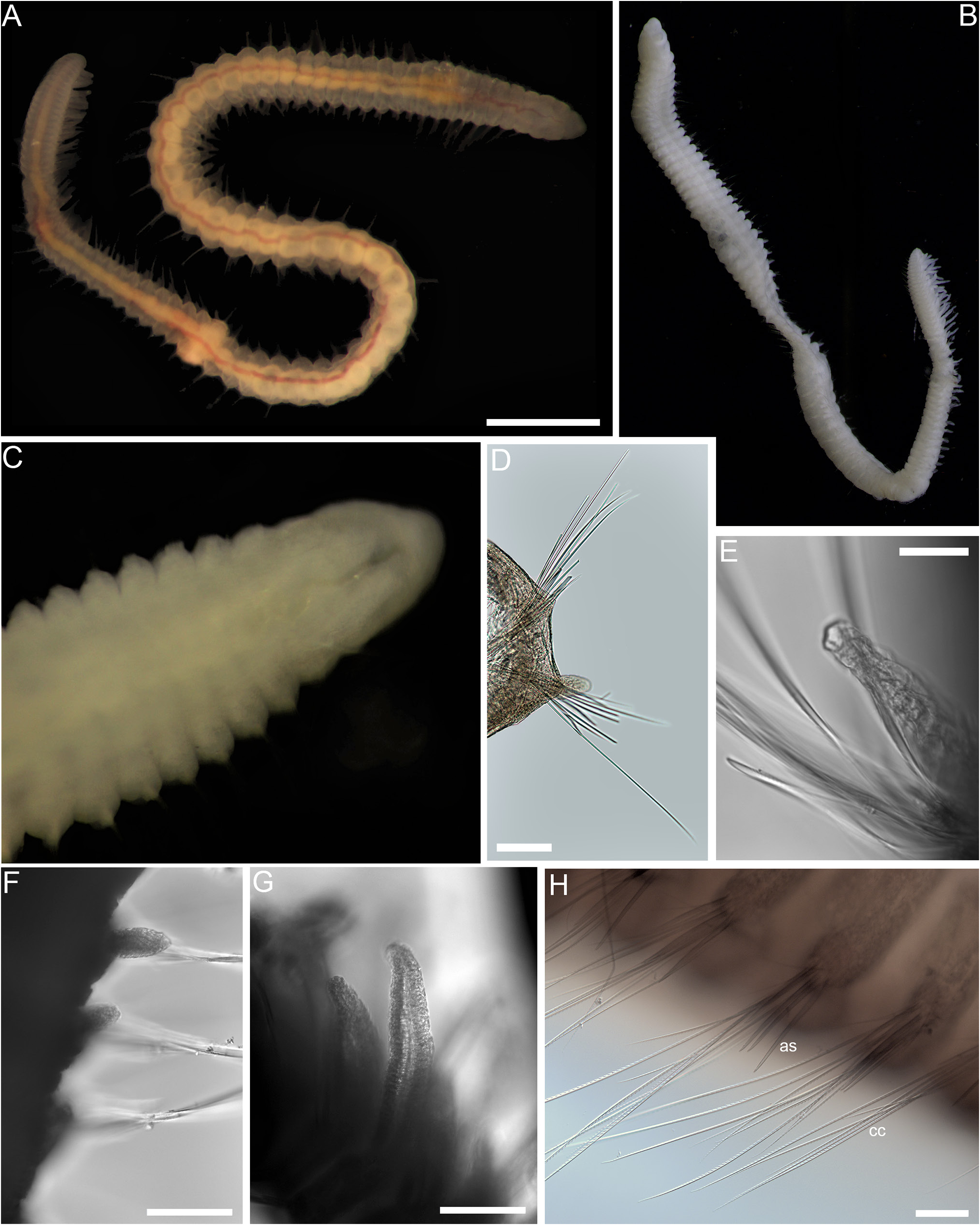
Description. Holotype AMW.53705 complete, ~ 8 mm long and 0.4 mm wide (widest, excluding chaetae) for ~84 chaetigers. Specimen NHMUKANEA 2023.1201 complete, ~ 7 mm long and 0.4 mm wide for ~80 chaetigers. Specimen NHMUKANEA 2023.1202 complete, ~ 7 mm long and 0.4 mmwide for ~80 chaetigers. SpecimenAM W.52199 anterior fragment with 21 chaetigers. Body somewhat dorsoventrally flattened throughout, not divided into distinct regions; longest chaetigers in mid-body, shortest in posterior body; posterior parapodia not dorsally elevated. Live specimen (holotype AM W.53705) reddish in colour ( Fig. 18A View Figure 18 ), ethanol-preservedspecimen off-white ( Fig. 18B View Figure 18 ).
Prostomium bluntly conical ( Fig. 18C View Figure 18 ), without appendages, eyes absent. Nuchal organs only detected as lateral pits on prostomium. Peristomium approximately twice as long as prostomium, weakly annulated dorsally, but distinct annulation observed ventrally, with two achaetous rings of similar size.
Notopodia low mounds from which chaetae emerge ( Fig. 18D View Figure 18 ); neuropodial postchaetal lobes from chaetiger 4 (5 in specimen AM W.52199), extending posteriorly to approximately start of branchiate region, whereafter they appear to be absent (or minute); best developed about mid-body where they slender, subconical-shaped, approximately 0.25× lengthof chaetae ( Fig. 18E View Figure 18 ). Branchiae present; absent in anterior-most segments; becoming apparent after segment 55. Branchiae initially small and ovoid ( Fig. 18F View Figure 18 ), increasing in length towards posterior to a maximum size of approximately ⅔ length of chaetae, strap-like ( Fig. 18G View Figure 18 ); reducing slightly in size over the last few chaetigers.
Chaetae include both crenulated capillaries ( Fig. 18H View Figure 18 ) and short acicular spines ( Fig. 18H View Figure 18 ) in both rami; furcate setae absent; no evidence of imbedded aciculae. Notochaetae bundles of crenulated capillaries of various lengths throughout; straight slightly serrated spines present from chaetiger 1 (up to 3 per ramus observed). Neurochaetae generally slendererthan notochaetaecomposed of crenulated capillaries and up to 3 spines; neuropodial spines slenderer and longer than those in notopodia, distally slightly curved into slender tip. Pygidium with two broad lobes, anal cirri not observed.
Distribution. IN2017_V03, Station 100. Pilot whale carcass, off Byron Bay, New South Wales, Australia in 999–1013 m.
Etymology. Named for James Hayhurst, for his support to one of the authors (M. Georgieva) during a multitude of scientific endeavours.
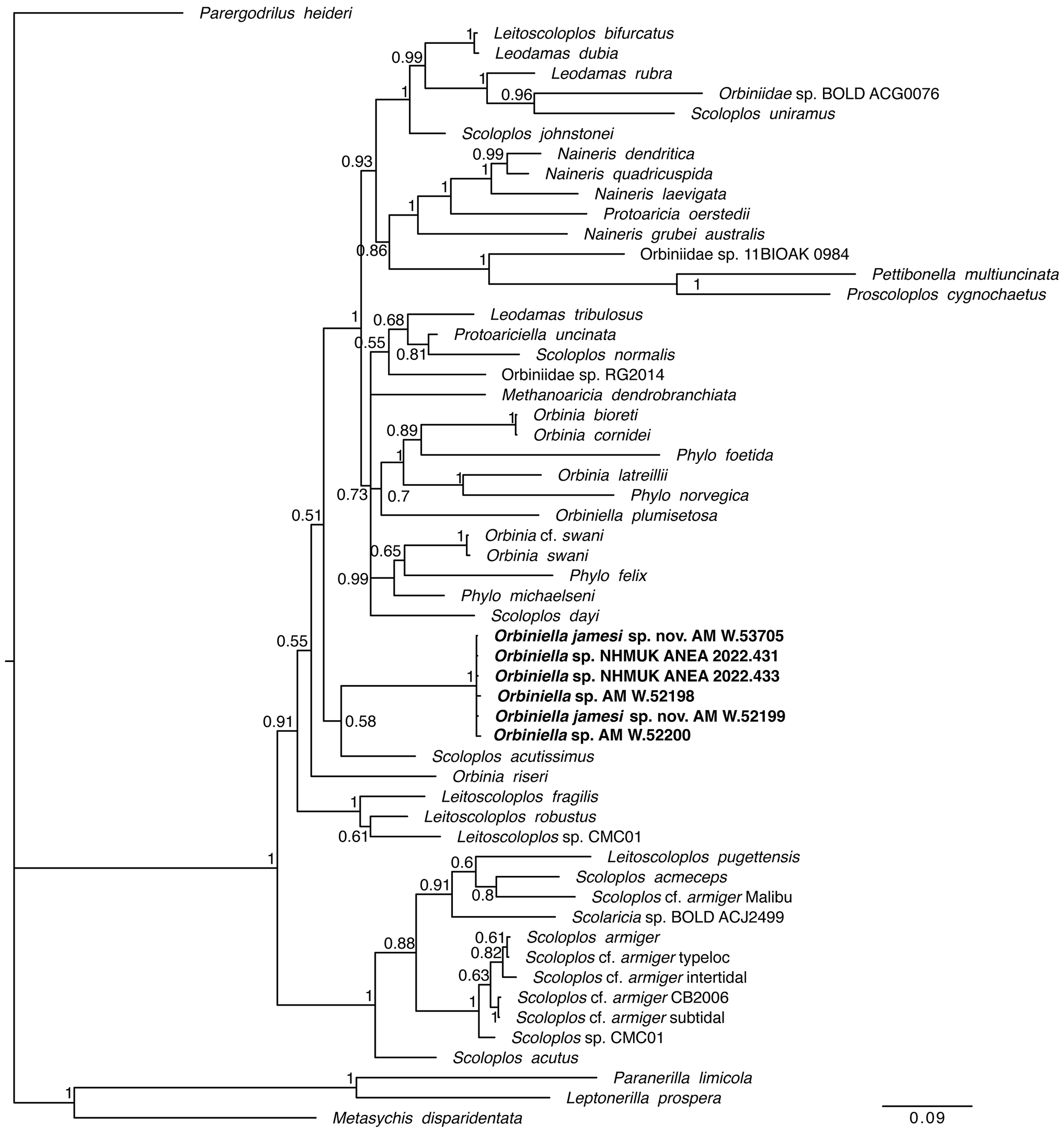
Remarks. Orbiniellajamesi sp. nov. specimens with neuropodial postchaetal lobes exhibit identical COI sequences to Orbiniella sp. without suchlobes ( Figs 19 View Figure 19 , 20 View Figure ). Support values in our phylogenetic analysis are generally low, with our specimens being resolved as most closely related toa Scoloplosacutissimus specimenwhereas theonly Orbiniella specimen for which genetic data was available falls outside of this group.
Specimens collected in this study belong to Orbiniidae that lack a distinct body division into thorax and abdomen regions due to a dorsal shift of chaetae. Such forms are currently included in genera Methanoaricia , Orbiniella (Parapar, Moreira, & Helgason, 2015) , Proscoloplos , Protoariciella , and Uncorbinia ( Beesley et al., 2000; Solis-Weiss & Fauchald, 1989; Blake, 2000; Parapar et al., 2015). Methanoaricia however differs from orbiniid specimens presented here in having a long and narrow prostomium, while Proscoloplos are generally small and along with Protoariciella and Uncorbinia have hooked chaetae. Uncorbinia has only asingledescribed speciesfrom northwestern Australia and is considered to be a probable synonym of Califia ( Blake, 2000) .
Assignment of the IN2017_V03 orbiniid specimens to the existing genus Orbiniella also reflects our molecular phylogenetic results for the family Orbiniidae ( Fig. 19 View Figure 19 ), which largely do not demonstrate clear genetic definitions. We therefore reserve the establishment of anew genus until current genetic relationships are better known, but we proceed with the formalization of new species Orbiniella jamesi sp. nov. Wetentativelyassign thenew speciesto genus Orbiniella duetopossession of abroadlyconical prostomium, bi-annulate peristomium, poorly developed parapodia, lack of furcate chaetae, no obvious division of body into thorax and abdomen, and no dorsal shift of parapodia.
To date, only one other orbiniid species is known from a chemosynthetic environment, Methanoaricia dendrobranchiata Blake, 2000 . This species has large branched branchiae which may be advantageous in the generally lower oxygen conditions prevalent in these environments. It is therefore possible that branchiae might be a character common to orbiniids that occur within chemosynthetic environments, however further discoveries are necessary to verify this.

| AMW |
Australian Museum |
| AM |
Australian Museum |
| NHMUKANEA |
Natural History Museum, London |
| COI |
University of Coimbra Botany Department |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |