
Milnesium irenae, Rocha & González-Reyes & Ostertag & Lisi, 2022
|
publication ID |
https://doi.org/10.5852/ejt.2022.822.1807 |
|
publication LSID |
lsid:zoobank.org:pub:522FD009-B4C9-4A80-871E-883E2EBE09C8 |
|
DOI |
https://doi.org/10.5281/zenodo.6620644 |
|
persistent identifier |
https://treatment.plazi.org/id/E52817A9-9080-427A-B4DF-AD4C58C52352 |
|
taxon LSID |
lsid:zoobank.org:act:E52817A9-9080-427A-B4DF-AD4C58C52352 |
|
treatment provided by |
Felipe |
|
scientific name |
Milnesium irenae |
| status |
sp. nov. |
Milnesium irenae View in CoL sp. nov.
urn:lsid:zoobank.org:act:
Figs 13–18 View Fig View Fig View Fig View Fig View Fig View Fig , Tables 3 View Table 3 , 6–7 View Table 6 View Table 7
Diagnosis
Peculiar, complex cuticular ornamentation including cuticular grooves, branched rugosity and a true, very fine, reticular design appearing different from that of the congeneric species with reticulated cuticle. Nine rows of pseudoplates present, formula: CP: I:0; II:4; III:4; IV:8; V:10; VI:8; VII:10; VIII:8; IX:12; X:4. Six peribuccal lamellae present, six peribuccal papillae with the medioventral reduced, two lateral cephalic papillae; buccal tube slightly funnel-shaped; stylets, their furcae and supports very developed. Claw configuration [2-3]-[2-2]; internal secondary branches of legs I–III with long basal spurs directed towards, almost touching the claw base. Ontogenetic change not observed in the available material, but hatchlings are probably lacking.
Etymology
We dedicate the new species to the researcher Irene Luisa Doma.
Material examined
Holotype ARGENTINA • ♀; La Pampa Province, Santa Rosa City ; 36º37′13″ S, 64º17′26″ W; about 177 m a.s.l.; 17 Sep. 2017; Rocha-Doma leg; moss and lichens from trees; UNLPam 1657(1). GoogleMaps
Paratypes ARGENTINA • 1 ♀; same collection data as for holotype; MCNS Tar.000023(2) GoogleMaps • 4 ♀♀; same collection data as for holotype; UNICT 5899(1) to 5899(4) GoogleMaps • 13 ♀♀; same collection data as for holotype; UNLPam 1367(1) , 1430(1) to 1430(3) , 1599(1) , 1599(3) , 1603(1) , 1603(3) , 1603(4) , 1656(1) to 1656(4) GoogleMaps .
Morphological description
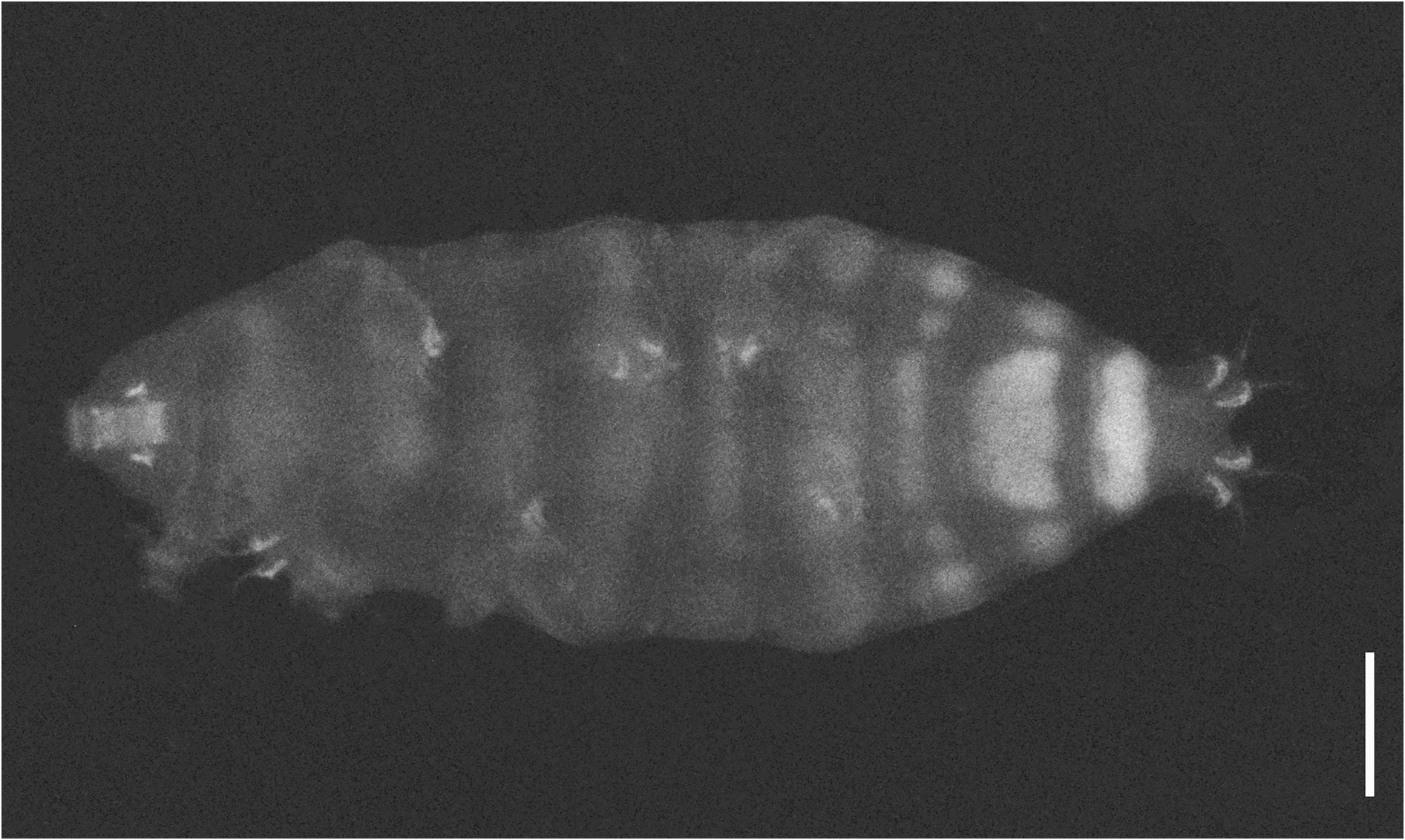
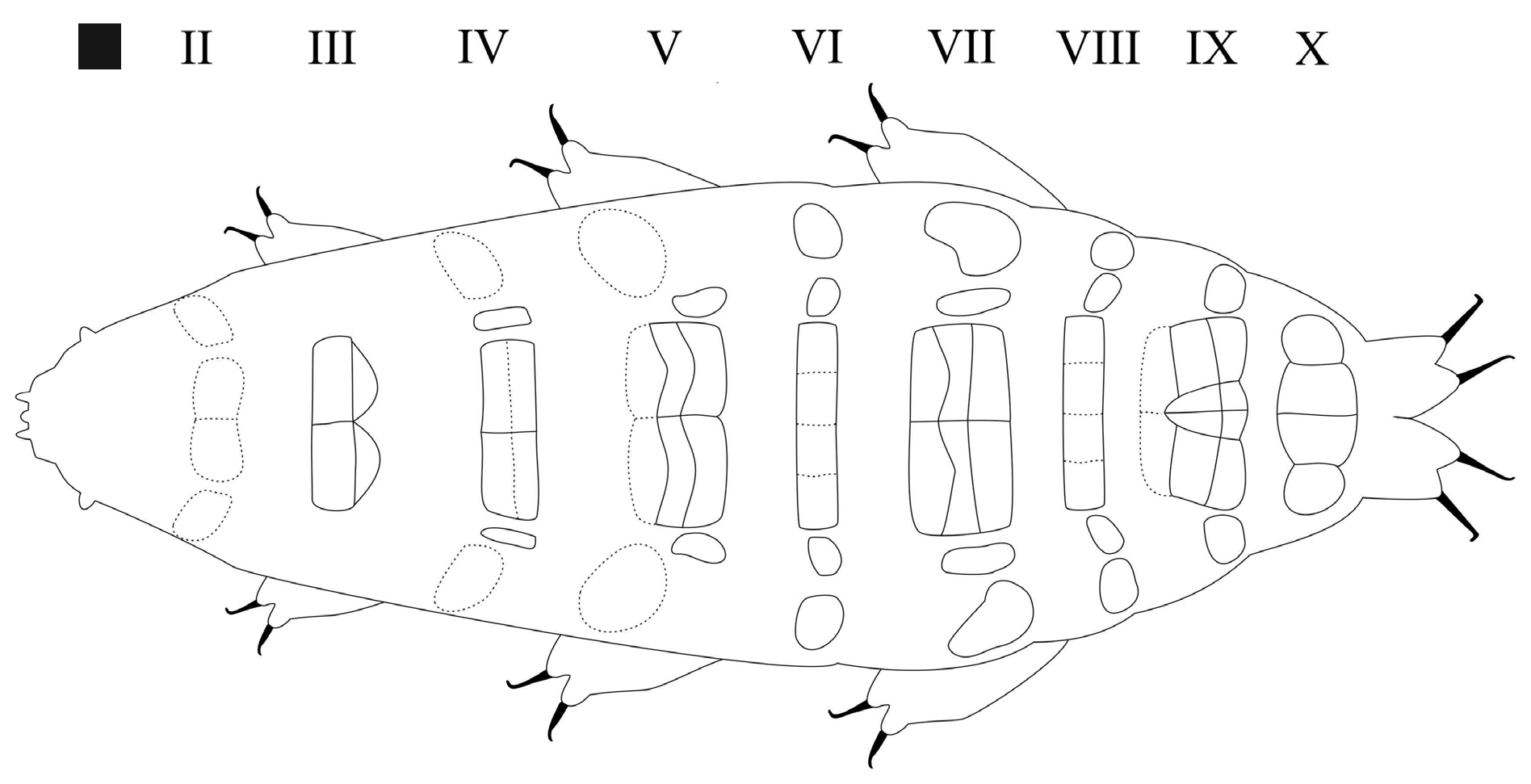
Body reddish up to 664 µm long (habitus in Fig. 13 View Fig ); large eyes present. Cuticle with complex sculpture ( Fig. 16 View Fig ): on the dorsal surface of trunk segments (starting with legs I in very few specimens, with legs II in some, and with legs III in the majority of them), clearly visible cuticular grooves ( Fig. 16A–B, D View Fig ) are present, some developing from cuticular invaginations for muscle attachments ( Fig. 16B View Fig arrows); the grooves form, more peripherally, a rugosity made of wrinkles that sometimes outline on some areas a network ( Fig. 16A–B View Fig arrowheads) visible as bright crossing lines delimiting dark elongated ‘dots’. The wrinkles gradually become interrupted and disappear, giving way to a reticular design ( Fig. 16 View Fig ) consisting of a fine-scale mesh, elongated in a few areas, isodiametric in the rest with a mesh diameter of about 0.5–1.0 µm. This reticule made of isodiametric mesh, though difficult to see, is quite spread on the cuticle, also on more cephalic segments where the other elements of the cuticular sculpture are absent: on all specimens it is visible at least starting with legs II, but in some it starts with legs I. This reticule, made of a delicate mesh, has a rather different appearance from the more common reticular pattern, resulting from close dimples, of many other species of Milnesium ; in M. irenae sp. nov. it looks instead more similar to the more irregular reticulations present in Parachela Schuster , Nelson, Grigarick & Christenberry, 1980, such as, for example, several species of Isohypsibiidae Sands, McInnes, Marley, Goodall-Copestake, Convey & Linse, 2008 (e.g., Dianea sattleri (Richters, 1902) and Ursulinius pappi (Iharos, 1966)) and Doryphoribiidae Gąsiorek, Stec, Morek & Michalczyk, 2019 (e.g., Doryphoribius bindae Lisi, 2011 and Grevenius kotovae (Tumanov, 2003)) . Pseudoplates present, arranged in 9 rows, formula: CP: I:0; II:4; III:4; IV:8; V:10; VI:8; VII:10; VIII:8; IX:12; X:4 ( Figs 14–15 View Fig View Fig ).
Row I is absent; row II, situated anteriorly to legs I, has four pseudoplates, all difficult to see: two medial, about quadrangular, touching in the central line, and two lateral, about trapezoidal, laying obliquely. Row III, situated in line with legs I, has four central pseudoplates arranged in two pairs, connected, with two anterior about rectangular, and two caudal, vaguely triangular pseudoplates. Row IV, situated between legs I and II, has eight pseudoplates: the four medial arranged in two pairs, transversally elongated, connected, forming a unique about rectangular structure; each of these four pseudoplates is also about rectangular but with the transverse line dividing the two anterior from the two posterior pseudoplates weakly outlined; two about rectangular pseudoplates, just lateral to the central complex, longitudinally elongated, plus two more lateral, bigger but difficult to see, about trapezoidal/ellyptical laying obliquely and slightly more anteriorly. Row V, situated in line with legs II, has ten pseudoplates: the six medial arranged in three pairs (the most cephalic difficult to see), transversally elongated, connected, forming a unique about rectangular structure; each these six pseudoplates vaguely rectangular but with the two transverse lines dividing the adjacent anterior/posterior pairs of pseudoplates not straight; two commashaped pseudoplates, just lateral to the central complex and aligned posteriorly, plus two more lateral pseudoplates, laying more anteriorly, bigger and about oval but difficult to see. Row VI, situated between legs II and III, has eight pseudoplates aligned in a single transverse row, with the four medial ones connected laterally and thus forming a unique wide rectangle (but with the longitudinal lines dividing the four pseudoplates weakly outlined); lateral to that central complex, two separate pseudoplates on each side, with the mid-lateral about oval, and the most lateral, bigger, more or less rounded/oval. Row VII, situated in line with legs III, has ten pseudoplates: the six medial arranged in three pairs, transversally elongated, connected, forming a unique about rectangular structure; each of these six pseudoplates vaguely rectangular but with the two transverse lines dividing the three pairs not straight; laterally, on each side, a longitudinally elongated pseudoplate, and a more lateral, bigger, about bean-shaped. Row VIII, situated just caudally to legs III, has eight pseudoplates aligned in a single transverse row, with the four medial connected laterally and thus forming a unique wide rectangle (but with the longitudinal lines dividing the four pseudoplates weakly outlined); lateral to that central complex, aligned with its posterior margin, two separate pseudoplates on each side, with the mid-lateral one about oval, and the most lateral one, bigger, more or less rounded/oval. Row IX, situated between legs III and IV, has twelve pseudoplates with a median aggregation of 10 pseudoplates (the more cephalic pair difficult to see) forming a complex pattern (see Fig. 15 View Fig ), and two lateral single pseudoplates about quadrangular/ rounded, aligned posteriorly. Row X, situated just anterior to legs IV, has four pseudoplates, sided and aligned in a single row transversally, with the medial ones about quadrangular, the lateral ones about rounded.
Six peribuccal lamellae present, and six peribuccal papillae plus two lateral papillae present; medio-ventral peribuccal papilla reduced, ( Fig. 17B View Fig arrow; pt of such papilla [ 7.3–10.4] vs [ 14.6–20.8] of the other peribuccal papillae). Buccal tube ( Fig. 17 View Fig ) not perfectly cylindrical, slightly wider anteriorly (posterior/ anterior width ratio 78–88%); stylets very robust, with very developed furcae, whale-tail shaped, and with supports which after their insertion on the buccal tube becoming gradually wider, assuming overall a triangular shape ( Fig. 17A, C View Fig ); pt of stylet support insertion point on the buccal tube length [ 68.9–74.9].
Claws of the Milnesium type with configuration [2-3]-[2-2] ( Fig. 18 View Fig ), rather robust, in particular the secondary branches; primary branches of legs IV more slender than on legs I–III (compare in Fig. 18A– B View Fig with C–D). Percentual ratio of secondary branches with respect to primary branches for each couple higher on legs I (79–96%), slightly lower on legs II–III (72–95%) and definitely lower on legs IV (63– 83%). Internal secondary branches of legs I–III with long basal spurs which form an acute angle inferiorly and have tip nearly reaching the claw base ( Fig. 18A View Fig ). Secondary branches with basal thickenings (‘lunulae’, Fig. 18A, C View Fig white arrowheads), primary branches with very thin, short, accessory points ( Fig. 18B View Fig black arrow); long and thick cuticular bars present under the claws I–III ( Fig. 18A View Fig black arrowhead). Tiny cuticular tubercles, often difficult to see, present on all legs, more visible on legs IV ( Fig. 18C–D View Fig white arrows).
Remarks
Milnesium irenae sp. nov. is the only described species of the genus with this form of complex cuticular sculpture, and with reticular pattern more similar to some Parachela than to other congeneric species with ‘reticulated’ cuticle. According to the literature, other congeneric species have a ‘reticule’ actually made of dimples close to one another, while in M. irenae the reticule mesh is not given by dimples. The medioventral peribuccal papilla is reduced (comments in Discussion). We found specimens with body length from 349 µm up to 664 µm (presumably, from second instar on) without remarkable differences between the smallest and the largest; therefore, possible ontogenetic change is unknown in the passage from hatchlings to second instar, while seems to be absent from second to third instar. Morphometric data are given in Table 6 View Table 6 and Supp. file 3; in Table 7 View Table 7 the statistically significant differences (through Student t -tests) of overlapping pt ranges of claw heights between the new species and the similar ones.
Differential diagnosis
Milnesium irenae sp. nov., due to its unusual cuticular sculpture, differs from all known species of the genus, though morphologically it has more affinity with some species of the old granulatum group. In addition, the characters of the stylets, their furcae and supports have peculiarities which differentiate it from many, if not all, congeneric species (the characters of those structures have not been described in detail in all past species descriptions). Here, we differentiate the new species from those having the same claw configuration [2-3]- [2-2] at least in some life stages, plus some kind of cuticular sculpture. These species are: M. almatyense Tumanov, 2006 (youngs and seniors), M. berladnicorum Ciobanu, Zawierucha, Moglan & Kaczmarek, 2014 , and adults of M. variefidum Morek, Gąsiorek, Stec, Blagden & Michalczyk, 2016 .
Milnesium irenae sp. nov. differs from them as follows:
1. Milnesium almatyense known from Kazakhstan ( type locality), Kyrgyz Republic ( Morek et al. 2020b), by different body colour: reddish in M.irenae sp. nov. vs white in M. almatyense ; eyes present in M. irenae vs absent of M. almatyense ; different and more complex cuticular sculpture in M. irenae (with tiny mesh, long, branched grooves, and tubercles) equal in specimens of all sizes found vs simpler (with delicate reticulation only in hatchlings, while only pseudopores in juveniles and adults) in M. almatyense ; nine rows of pseudoplates in M. irenae vs eight in M. almatyense with different number of pseudoplates in each correspondent row ( Table 3 View Table 3 ); higher pt of buccal tube standard width, [ 46.1–56.9] in M. irenae vs [ 26.2–33.1] in M. almatyense ; higher buccal tube standard width/length ratio, 46–57% in M. irenae vs 26–33% in M. almatyense .
2. Milnesium berladnicorum only known from the type locality in Romania, by different body colour: reddish in M. irenae sp. nov. vs brownish in M. berladnicorum ; different and more complex cuticular sculpture in M. irenae (with tiny mesh, long, branched grooves, and tubercles) vs simpler (with just bright spots, not ascertained whether either dimples or pseudopores) in M. berladnicorum ; different number of pseudoplates in each correspondent row ( Table 3 View Table 3 ); statistically significant higher buccal tube posterior/anterior width ratio, 78–88%, mean 82%, in M. irenae vs 69–79%, mean 73%, in adults of M. berladnicorum ; (t
25 = 8.09, p <0.001).
3. Milnesium variefidum only known from the type locality in Scotland, by different body colour: reddish in M. irenae sp. nov. vs white or transparent in M. variefidum ; eyes present in M. irenae vs absent in M. variefidum ; different cuticular ornamentation, with tiny mesh, long, branched grooves, and tubercles in M. irenae vs with scattered pseudopores, minute wrinkles and only some short grooves in M. variefidum ; presence of nine rows of pseudoplates in M. irenae vs seven rows in M. variefidum with different number of pseudoplates in each correspondent row ( Table 3 View Table 3 ); statistically significant differences about pt of almost all claw heights ( Table 7 View Table 7 ).




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |