
Myriochele heeri Malmgren, 1867
|
publication ID |
https://doi.org/10.1080/00222930600711758 |
|
persistent identifier |
https://treatment.plazi.org/id/DD1C87D7-9E2D-FFB4-FE19-74BFFC46F9FF |
|
treatment provided by |
Felipe |
|
scientific name |
Myriochele heeri Malmgren, 1867 |
| status |
|
Myriochele heeri Malmgren, 1867 View in CoL , emended
( Figures 1 View Figure 1 , 2 View Figure 2 , 3B View Figure 3 , 4A, B View Figure 4 , 5 View Figure 5 , 6 View Figure 6 , 7A View Figure 7 , 12 View Figure 12 , 13 View Figure 13 ; Tables I, II)
Myriochele heeri Malmgren 1867, p 101 View in CoL –102, Table 7, Figure 37; Blake and Dean 1973, p 37, Figure 2 View Figure 2 ; Nilsen and Holthe 1985, p 21 –22, Figures 3 View Figure 3 , 4 View Figure 4 , 11c–e View Figure 11 ; Parapar 2003a, p 283 –284 [pro parte].
? Myriochele heeri: Imajima and Morita 1987, p 90 View in CoL –91, Figures 3a–k View Figure 3 , 4e–h View Figure 4 .
Type material
Swedish Museum of Natural History. The revised part of the collection consists of two jars. The first jar, labelled ‘‘ SMNH —Type—5847 syntype (s). Myriochele heeri Malmgren, 1867 . Svalbard, Vest Spitsbergen, Safe Haven. Leg. A. J. Malmgren 1864’’ has a vial inside labelled ‘‘ Myriochele heeri # 905 (6518), S. C. Byers 4 Oct. 2002 ’’ with three specimens, one of them ( Figure 3B View Figure 3 ) selected as lectotype (see below). The second jar, labelled ‘‘ SMNH —Type—5848 syntype (s). Myriochele heeri Malmgren 1867 . NW Greenland, Umanak, depth 455 m. Leg. C. T. Amondsen’’ also has a vial inside, labelled ‘‘ Myriochele heeri # 906, 2/ 2 specimen + tube (6517)’’ with two specimens supposedly also revised by S. C. Byers.
Additional material
Icelandic Museum of Natural History. BIOICE sample 2401 ( 13 specimens (spms); IMNH 4100 ) ; sample 2423 (six spms; IMNH 4101 ) ; sample 2424 (three spms; IMNH 4102 ) ; sample 2474 (one spm; IMNH 4103 ) ; sample 2691 (four spms; IMNH 4104 ) .
Museo Nacional de Ciencias Naturales. BIOICE sample 2080 (one spm; MNCN 16.01/ 10532); sample 2257 (two spms; MNCN 16.01/10533); sample 2303 (seven spms; MNCN 16.02/10534); sample 2400 (six spms; MNCN 16.01/10535).
malmgreni n. sp. (relative length of fourth and fifth chaetigers in M. islandica were taken from paratypes).
Swedish Museum of Natural History. BIOICE sample 2700 (nine spms; SMNH 83573).
National Museum of Natural History. East Angiak Island , Baffin Island region , Canadian Arctic Archipelago , st. 20A-B, 14 August 1968, 132– 245 m depth (more than five spms; USNM 51164 About USNM ). Southern Baffin Bay between W. Greenland and East Baffin Island: 67 ° 499N, 60 ° 469W, st. 26A, 16 August 1968, 1920– 1745 m depth (one spm; USNM 51165 About USNM ) .
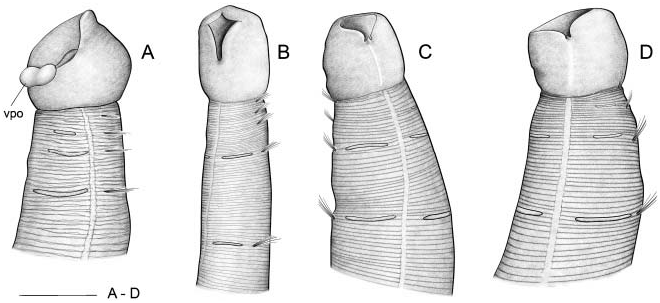
Description (based on lectotype of M. heeri ) ( Figure 3B View Figure 3 )
Body short, 19 mm long and 1.08 mm wide, very much shrivelled longitudinally and posteriorly tapered. Head region not rounded probably because of fixation; distinctly set off from thorax by a constriction. Mouth forming a longitudinal slit in the antero-ventral part of the head region. Eye-spots not visible. First three chaetigers uniramous ( TF 51:0-1:0- 1:0), with RLTC 51-1-2, followed by 25 biramous chaetigers. Length of first four chaetigers: 0.18, 0.23, 0.38, and 1.13 mm, respectively. Fourth to sixth or seventh chaetigers increasing in length; fourth and fifth chaetigers about 1.5 and 2.5 times longer than thorax; sixth and seventh chaetigers the longest, about three times longer than thorax. Following chaetigers progressively decreasing in length, becoming crowded in prepygidial segments. Notopodia with a dorsal bundle of capillary chaetae and a ventral bundle of short and smooth acicular chaetae. Neuropodial tori narrow, with uncini arranged in irregular rows. Hooks with two teeth constantly arranged one above the other. Posterior end distally tapered, with segments highly compressed and pygidium contracted, appearing as two low lateral lobes.
Description of Icelandic specimens
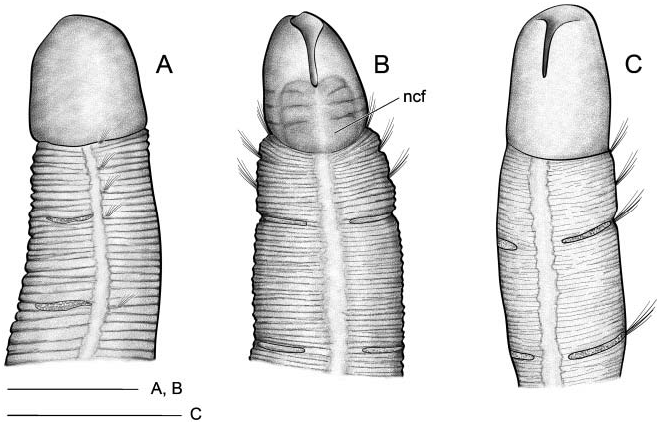
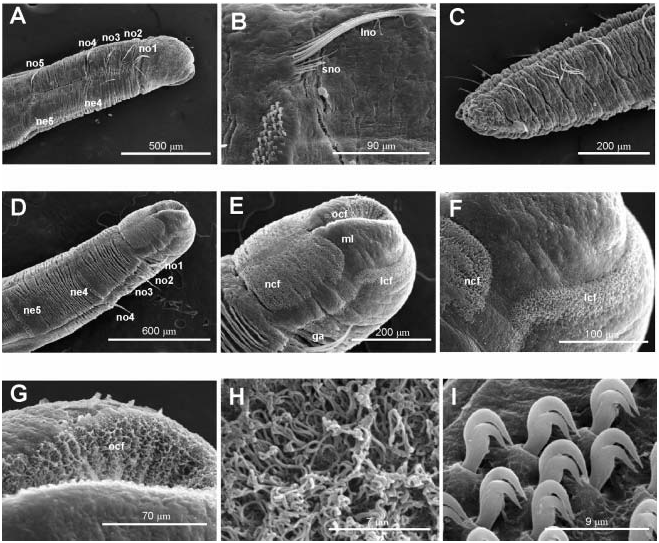
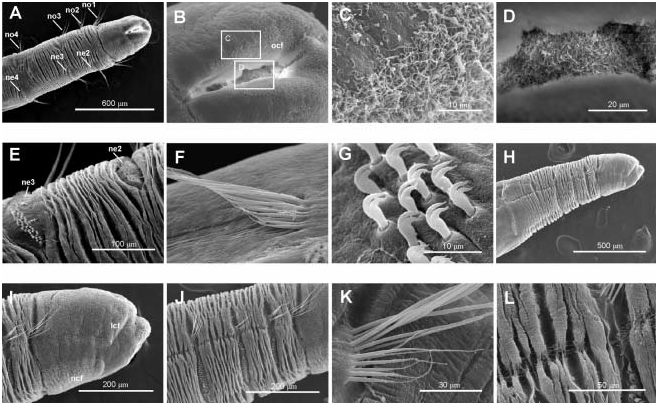
Body short, 10–12 mm long and 0.4–0.5 mm wide, 22–25 chaetigers. First three body chaetigers short and uniramous ( TF 51:0-1:0-1:0), shorter than wide, with relative length 1-1-1(1.5) ( Figures 4A, B View Figure 4 , 5A View Figure 5 , 12 View Figure 12 ); fourth to sixth or seventh chaetigers increasing in length. Fourth chaetiger, first biramous, 1.5–2 times thoracic length. From chaetiger eight, body segments decreasing in length. Notopodia with long capillary chaetae and short acicular chaetae ( Figure 5B View Figure 5 ); neuropodial hooks with two apical teeth located one above the other ( Figure 5I View Figure 5 ); pygidium with two low lateral lobes ( Figure 5C View Figure 5 ).
Two ciliated fields distinct using SEM at head level in BIOICE specimens: a front lateral field dorsal to the mouth lips and a neck field ventral to the mouth lips ( Figures 4B View Figure 4 , 5D–F View Figure 5 ). Oral cilia present on the inner side of the mouth lips ( Figure 5E, G, H View Figure 5 ).
Tube with cylindrical middle part and long tapering ends, covered by sponge spicules or sand grains depending on bottom nature ( Figure 13 View Figure 13 ).
Remarks
As Malmgren did not define holotype or paratypes and S. C. Byers, who revised the collection in 2002, created no lectotype or paralectotypes, the syntype proposed herein as lectotype for Myriochele heeri ( Figure 3B View Figure 3 ) was selected for three reasons:
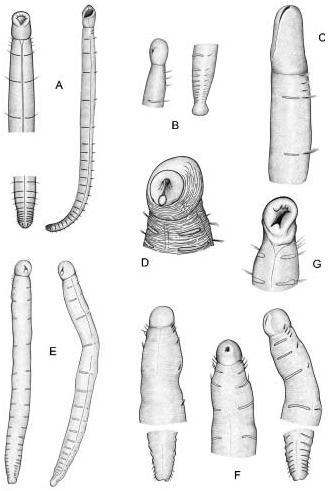
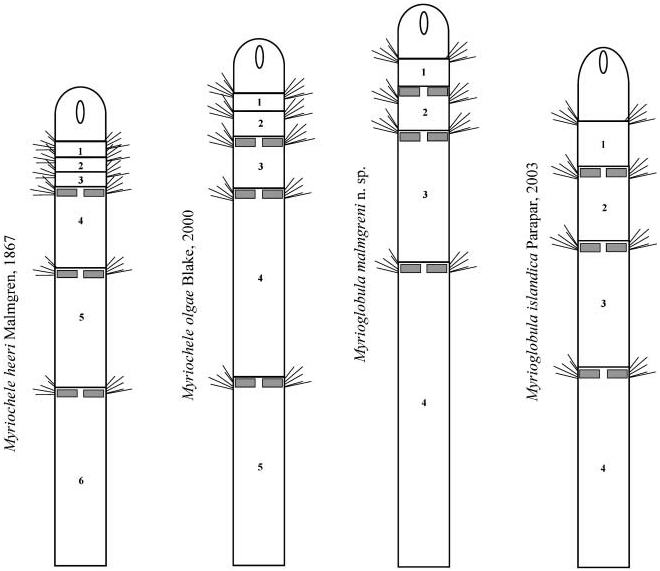
1. Both TF and RLTC are in accordance with Malmgren’s original description (see Introduction) and the general body appearance traditionally assigned to this taxon in classical literature (see Figure 1 View Figure 1 ).
2. The vial # 5847 with specimens from Spitzbergen, according to Byers’ notes, probably belongs to the original Malmgren types.
3. Even though it is wrinkled, it is in better condition than the other two specimens from Malmgren’s collection also assigned by the author to this species.
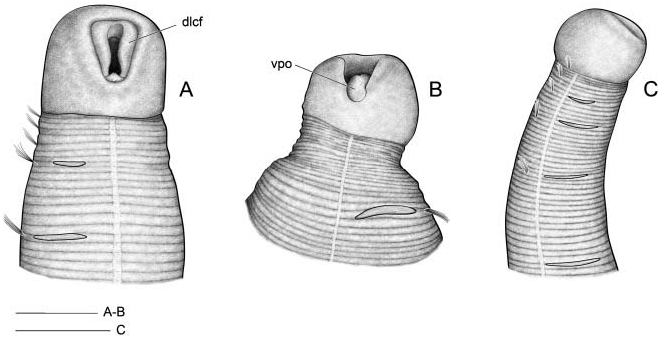
The three specimens in vial # 5847 of the M. heeri type series correspond to two species. One of them ( Figure 3A View Figure 3 ) has an asymmetric second anterior chaetiger giving rise to an anterior region with two uniramous chaetigers on the right side and only one on the left side ( TF 51:0-1:0(1:1)-1:1). Relative length of thoracic chaetigers, RLTC 51-1-3, with chaetigers 4–6 of similar length, the fourth being about 1.5 times longer than the sum of the preceding. The mouth is widely open with the ventral pharyngeal organ everted. This specimen corresponds to those identified as Myriochele aff. olgae from the BIOICE material (see below).
The other two specimens correspond to M. heeri: TF 51:0-1:0-1:0; RLTC51-1 - 1.5 (2) with chaetigers 4–6 increasing in length, the fourth being about 1.75 times the sum of the preceding. One of them is damaged (not illustrated) and the other, which was selected as lectotype ( Figure 3B View Figure 3 ), is much wrinkled longitudinally .
In vial # 5848 of the M. heeri type series, one specimen is damaged (not illustrated, corresponding to M. heeri ), but the other is in good condition, again with asymmetric anterior chaetigers and RLTC 51-1.5-3.5; this specimen is assigned again to M. aff. olgae ( Figure 3C, D View Figure 3 ).
Malmgren’s assumption that the four anterior chaetigers, instead of three, are of similar length (see Introduction) was probably due to a different concept in chaetiger delineation. Considering the relative proportions of anterior chaetigers to be of high taxonomic relevance, a clear delimitation of them is a matter of great importance. Following Blake (2000), who stated that in oweniids parapodia are located at the anterior border of segments (even though this is difficult to discern in Myriochele , a genus without clear segment delineation—but easily observed in Myriowenia ; see Blake 2000, Figure 5.7 View Figure 5 ), a large fourth chaetiger is revealed in this species. Nevertheless, the assumption of a middle position of the parapodia in the chaetiger, which is the generalized case in polychaetes and the idea probably assumed by Malmgren, led to the view of a short fourth chaetiger.
As was previously noted, the number and relative length of anterior body chaetigers are traditionally considered to have crucial taxonomic value for species discrimination in the genus Myriochele , with M. heeri characterized by the presence of three short uniramous chaetigers of similar length, while specimens with two uniramous chaetigers represent a different species (see below).
Myriochele gracilis Blake, 2000 View in CoL from off California and Myriochele riojai Parapar, 2003 View in CoL from Bransfield Strait ( Antarctica) share with Myriochele heeri View in CoL two of the most relevant diagnostic characters in Myriochele View in CoL systematics and qualifies them as very close species. These characters are the thoracic formula, TF51:0-1:0-1:0, and the relative length of thoracic chaetigers, RLTC51-1-1(1.5). Nevertheless, Myriochele gracilis View in CoL differs from M. heeri View in CoL in the prostomial shape—domed dorsally and with numerous small papillae in M. gracilis View in CoL and without these characters in M. heeri View in CoL — and the length of the first abdominal chaetigers—increasing in length from fourth to seventh or eighth in M. heeri View in CoL and all of consistent length in M. gracilis View in CoL . Myriochele riojai View in CoL differs from M. heeri View in CoL both in known geographic distribution (Antarctic versus Arctic and Boreo-Atlantic waters), by its body size ( 10–12 mm versus 23 mm), and by the position and number of notopodial acicular chaetae (much more numerous and present from the first chaetiger in M. riojai View in CoL ).
Although no specimens were examined, the descriptions and illustrations provided by Imajima and Morita (1987) of Japanese specimens of M. heeri View in CoL ( Figure 1F View Figure 1 ) leave little doubt about their correct identification. No specimens with two thoracic chaetigers were referred to by the authors.
The material collected by Blake and Dean (1973) consists of two vials. Vial # 51165 contains one damaged indeterminable specimen and vial # 51164 contains nine specimens out of their tubes and about 16 still inside. Among the former, five have the characteristics here accepted for M. heeri ( Figure 7A View Figure 7 ), one is regenerating the anterior end ( Myriochele indet., Figure 7B View Figure 7 ), and three, although in general appearance similar to M. heeri , belong to M. aff. olgae ( Figure 7C View Figure 7 ), with TF51:0-1:0-1:1 and RLTC51-1-2. One specimen identified as M. heeri shows the dorsolateral ciliary folds semi-everted ( Figure 7A View Figure 7 ) and the specimen identified as Myriochele indet. also shows the pharyngeal organ everted ( Figure 7B View Figure 7 ), as in Myriochele aff. olgae from SMNH 6518 ( Figure 3A View Figure 3 ).
Distribution
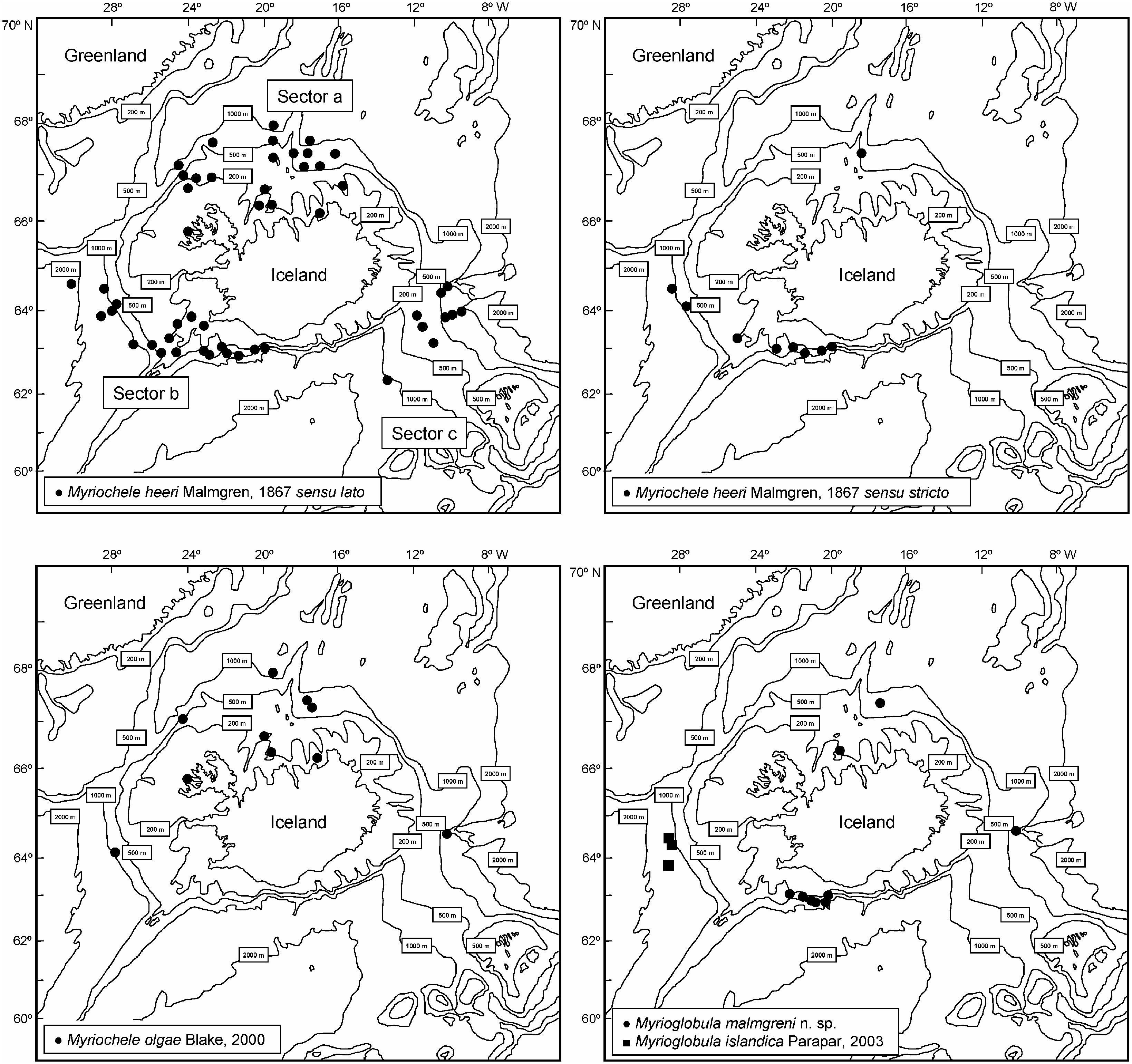
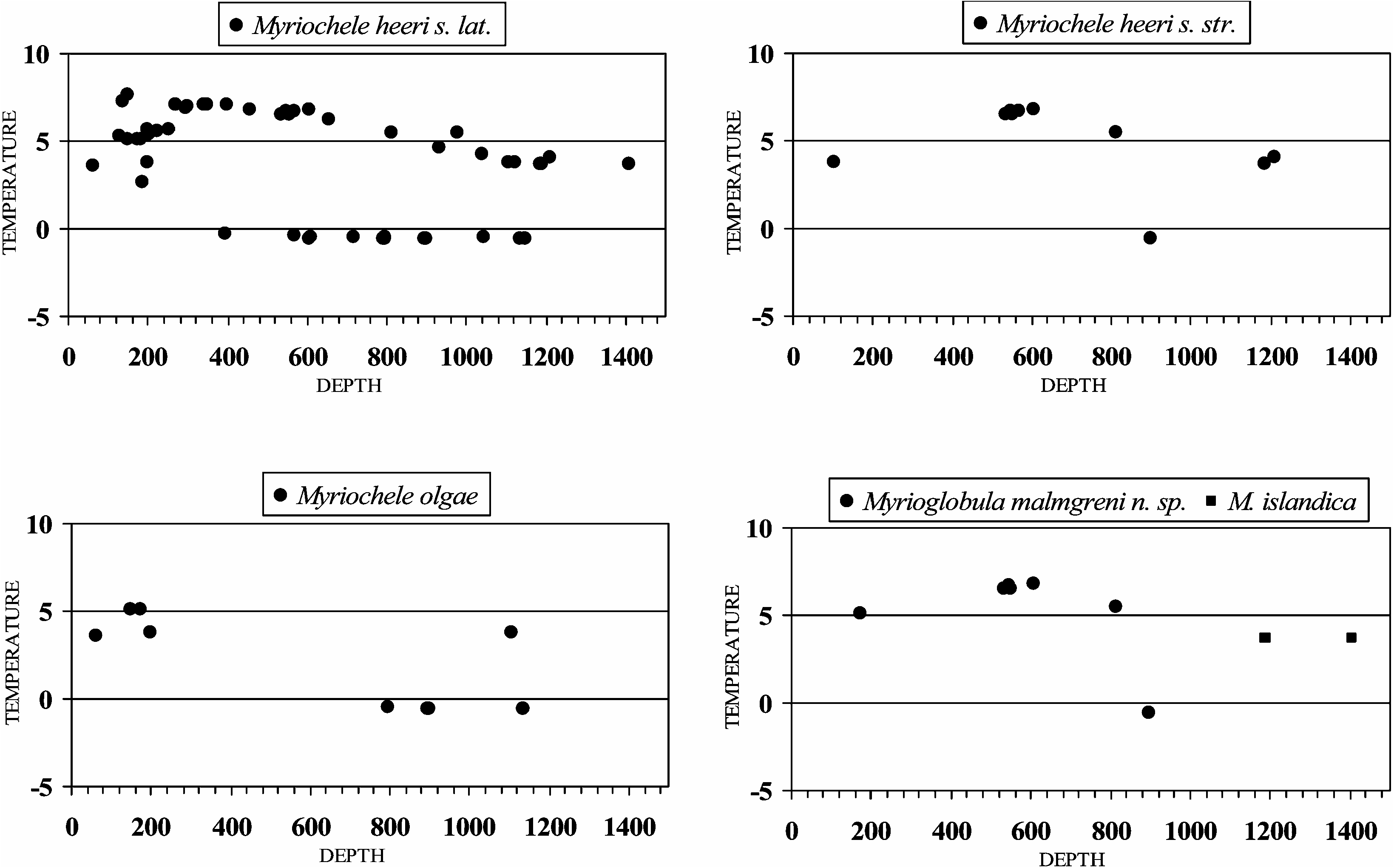
The revised material of M. heeri reveals an Arctic and Boreo-Atlantic distribution. With the only exception of a specimen collected on the northern coast (sector a) from sample 2080 ( 898.5 m depth and 20.6 ° C), the specimens from the BIOICE project were collected in sector b (southwest coast–Reykjanes Peninsula) in warm waters (3.8–6.8 ° C) and at depths ranging from 535 to 1210.5 m ( Figures 2 View Figure 2 , 6 View Figure 6 as M. heeri sensu stricto).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Myriochele heeri Malmgren, 1867
| Parapar, Julio 2006 |
Myriochele heeri :
| Imajima M & Morita Y 1987: 90 |
Myriochele heeri
| Parapar J 2003: 283 |
| Nilsen R & Holthe T 1985: 21 |
| Blake JA & Dean D 1973: 37 |
| Malmgren AJ 1867: 101 |