
Myriochele olgae Blake, 2000
|
publication ID |
https://doi.org/10.1080/00222930600711758 |
|
persistent identifier |
https://treatment.plazi.org/id/DD1C87D7-9E27-FFB7-FE41-7613FF26FA9C |
|
treatment provided by |
Felipe |
|
scientific name |
Myriochele olgae Blake, 2000 |
| status |
|
Myriochele olgae Blake, 2000 View in CoL
( Figures 2 View Figure 2 , 3A, C, D View Figure 3 , 4C View Figure 4 , 6 View Figure 6 , 7C View Figure 7 , 8 View Figure 8 , 9 View Figure 9 , 12 View Figure 12 , 13 View Figure 13 ; Tables I, II)
Myriochele olgae Blake 2000, p 111 View in CoL –113, Figure 5.5 View Figure 5 .
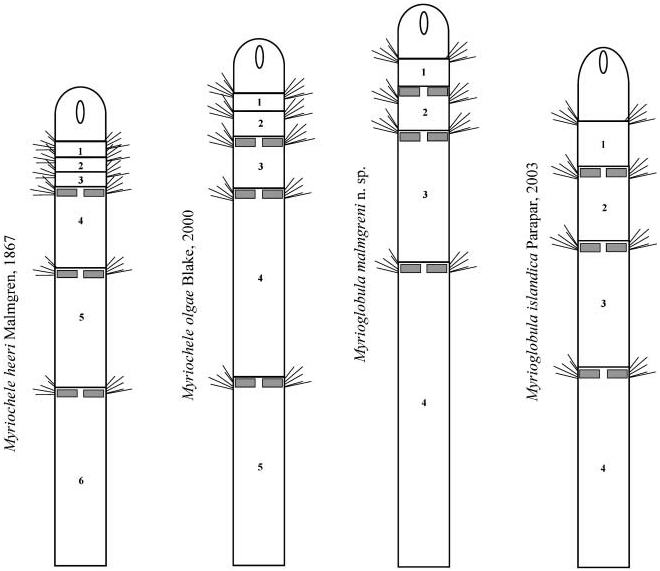
Myriochele heeri Malmgren 1867, p 101 View in CoL –102, Table 7, Figure 37; Blake and Dean 1973, p 37, Figure 2 View Figure 2 ; Parapar 2003a, p 283 –284 [pro parte].
Myriochele heeri: Nilsen and Holthe 1985, p 21 View in CoL –22, Figures 3 View Figure 3 , 4 View Figure 4 , 11c–e View Figure 11 [? pro parte].
Type material
National Museum of Natural History. North Pacific Ocean, USA, California, Santa Maria Basin, off Point Sal , 145 m ( holotype; USNM 186431 About USNM ) .
Additional material
Icelandic Museum of Natural History. BIOICE sample 2575 (five spms; IMNH 4105); sample 2700 (three spms; IMNH 4106).
Museo Nacional de Ciencias Naturales. BIOICE sample 2065 (five spms; MNCN 16.01 About MNCN / 10536); sample 2080 (four spms; MNCN 16.01 About MNCN /10537); sample 2081 (one spm; MNCN 16.01 About MNCN /10538); sample 2100 (nine spms; MNCN 16.01 About MNCN /10539); sample 2152 ( 13 specimens; MNCN 16.01 About MNCN /10540); sample 2167 (three spms; MNCN 16.01 About MNCN /10541) ;
sample 2364 (seven spms; MNCN 16.01/10542); sample 2480 (one spm; MNCN 16.01/ 10543).
Description based on Icelandic specimens
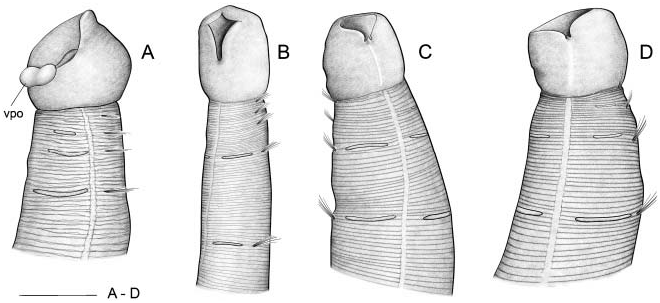
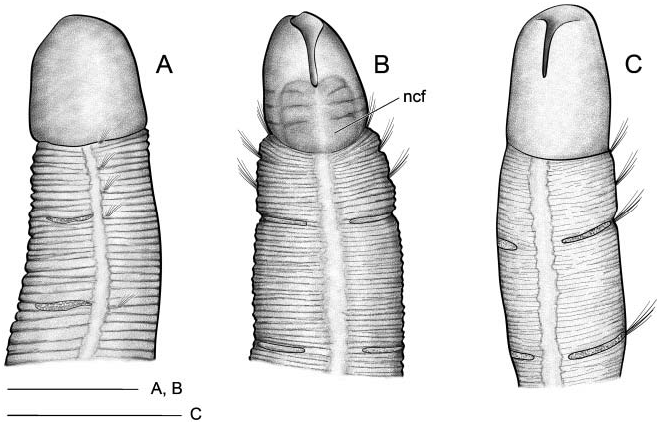
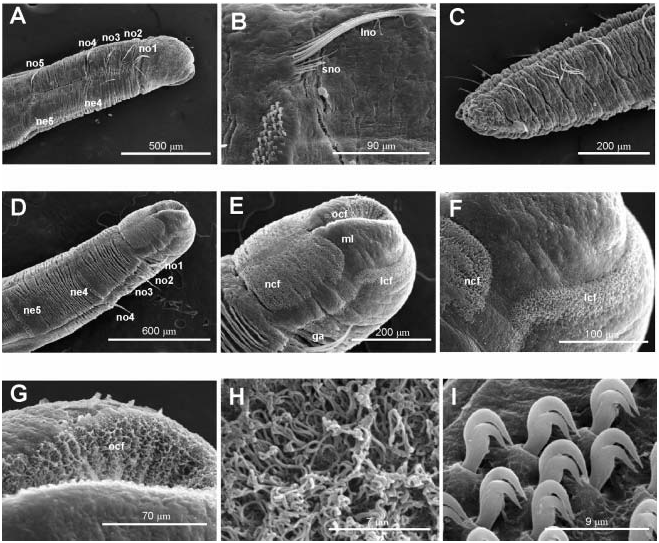
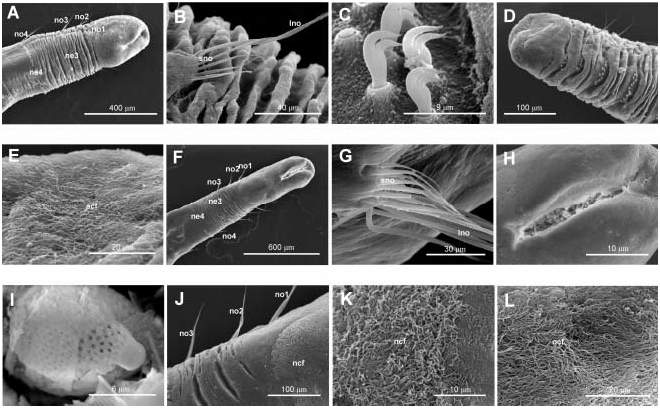
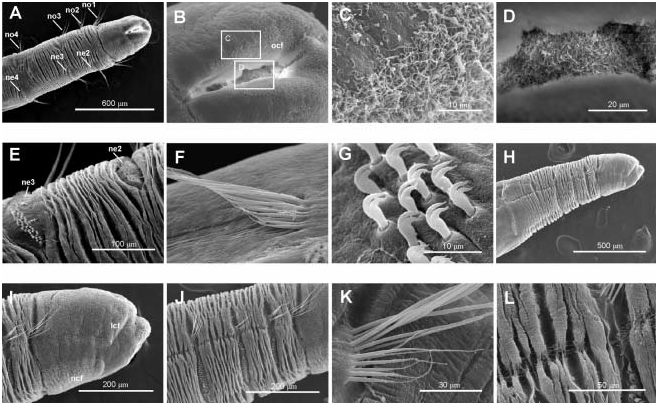
Body short, 5–10 mm length and 0.5 mm wide, 18–23 chaetigers. First two body chaetigers uniramous; third biramous ( TF 51:0-1:0-1:1), first and second shorter than wide and third longer than wide, with increasing length ( RLTC 51-1(1.5)-3) ( Figures 4C View Figure 4 , 8A, F View Figure 8 , 12 View Figure 12 ). Fourth to sixth chaetigers (second to fourth abdominal) elongated and of similar length; fourth about two (1.5) times as long as the sum of three previous chaetigers. Following chaetigers decreasing in length. Notopodia with long capillary chaetae and short acicular chaetae ( Figure 8B, G View Figure 8 ); neuropodial hooks with two apical teeth located one above the other ( Figure 8C View Figure 8 ); pygidium with two low lateral lobes ( Figure 8D View Figure 8 ).
As in M. heeri , ciliated fields associated with head and mouth distinct ( Figure 8F, J–L View Figure 8 ); a dense ciliated anal field observed at the pigidium ( Figure 8D, E View Figure 8 ); food remains, possible diatom frustules, present inside the mouth of one specimen ( Figure 8H, I View Figure 8 ).
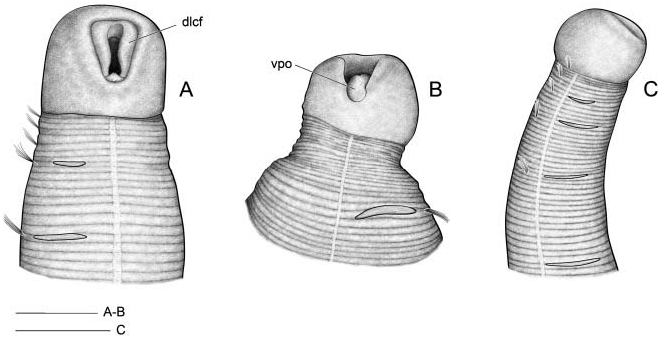
Tube with cylindrical middle part and long tapering ends, covered by sponge spicules or sand grains depending on bottom nature ( Figure 13 View Figure 13 ).
Distribution
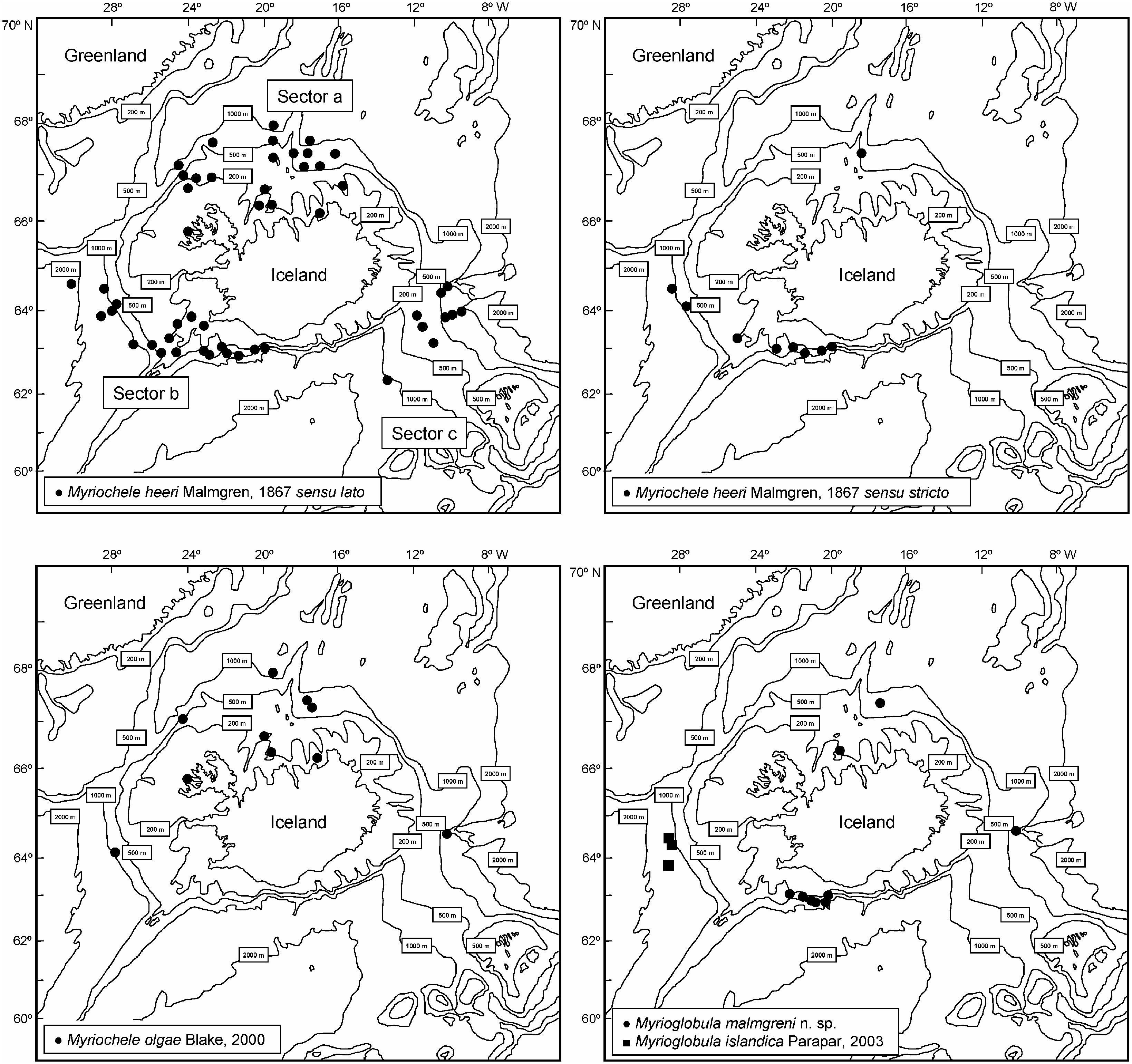
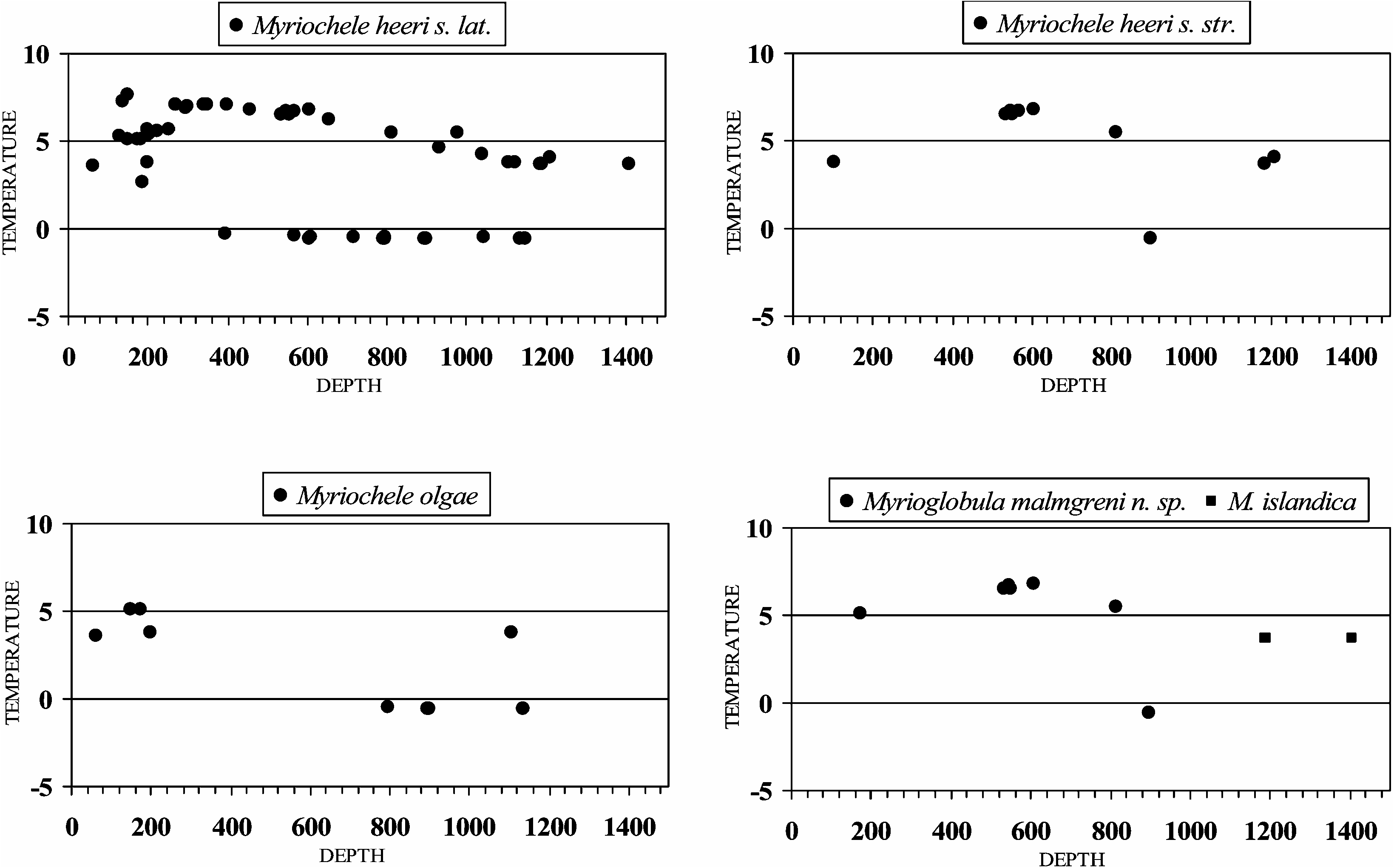
Myriochele olgae was originally described from the East Pacific coast of North America (California) and is now reported from Iceland. In Icelandic waters, the species seems to be restricted to sector a on the north coast ( Figure 2 View Figure 2 ) at a wide depth ( 198–1136.5 m) and temperature (20.6–5.1 ° C) range ( Figure 6 View Figure 6 ).
Remarks
The investigation of the holotype of Myriochele olgae ( USNM 186431) showed that the general body shape, size, number of chaetigers, and thoracic formula of the specimen are completely in agreement with the Icelandic specimens (Table II). The only difference was in the relative length of the anterior biramous chaetigers; in the holotype chaetiger 3 is about four times longer than the uniramous chaetigers (chaetigers 1 and 2) and chaetiger 4 is the longest of the body, the following being shorter. In Icelandic specimens chaetiger 3 is about three times longer than uniramous chaetigers and an increasing length is observed from the fourth to sixth chaetigers. In both cases chaetiger 4 is about two times longer than the first three chaetigers combined. In my opinion, those differences are not significant enough to describe a new species.
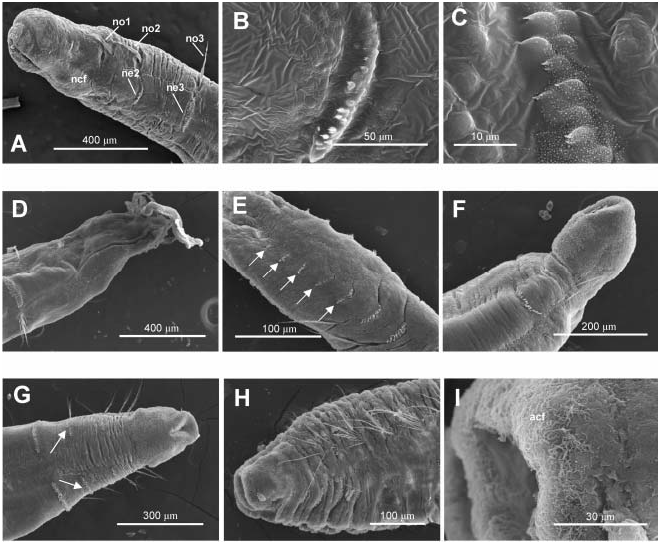
Specimens named Myriochele aff. olgae from BIOICE samples 2423 and 2424 ( Figure 9 View Figure 9 ) show some abnormal characters such as: (1) asymmetrical second biramous chaetiger with left neuropodial hooks partially piercing the epidermis ( Figure 9A–C View Figure 9 ); (2) anterior end broken or in process of regeneration ( Figure 9D, F View Figure 9 ); (3) a supernumerary second biramous chaetiger ( Figure 9G View Figure 9 ); and (4) last chaetigers without notopodia ( Figure 9E View Figure 9 ). Cilia associated with pygidial lobes were also seen ( Figure 9H, I View Figure 9 ).
Unlike M. heeri View in CoL , in which the principal taxonomic characters (number of uniramous/ biramous anterior chaetigers and their relative length) seem to be consistent, the Icelandic specimens of Myriochele olgae View in CoL show a high variability in the presence of asymmetric anterior chaetigers and differences in the relative chaetiger length (see above). This variability would correspond to decapitation due to predation or an asexual reproduction process by bipartition, a reproductive strategy previously reported by Parapar (2003a) in the BIOICE oweniids Galathowenia fragilis ( Nilsen and Holthe, 1985) View in CoL and Galathowenia oculata ( Zachs, 1923) View in CoL .

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Myriochele olgae Blake, 2000
| Parapar, Julio 2006 |
Myriochele olgae
| Blake JA 2000: 111 |
Myriochele heeri : Nilsen and Holthe 1985 , p 21
| Nilsen R & Holthe T 1985: 21 |
Myriochele heeri
| Parapar J 2003: 283 |
| Blake JA & Dean D 1973: 37 |
| Malmgren AJ 1867: 101 |