
Dibamus booliati, Das & Yaakob, 2003
|
publication ID |
https://doi.org/10.5281/zenodo.4618718 |
|
persistent identifier |
https://treatment.plazi.org/id/DA57879D-7869-FF8D-A118-FCEE5D6D3908 |
|
treatment provided by |
Carolina |
|
scientific name |
Dibamus booliati |
| status |
sp. nov. |
Dibamus booliati , new species
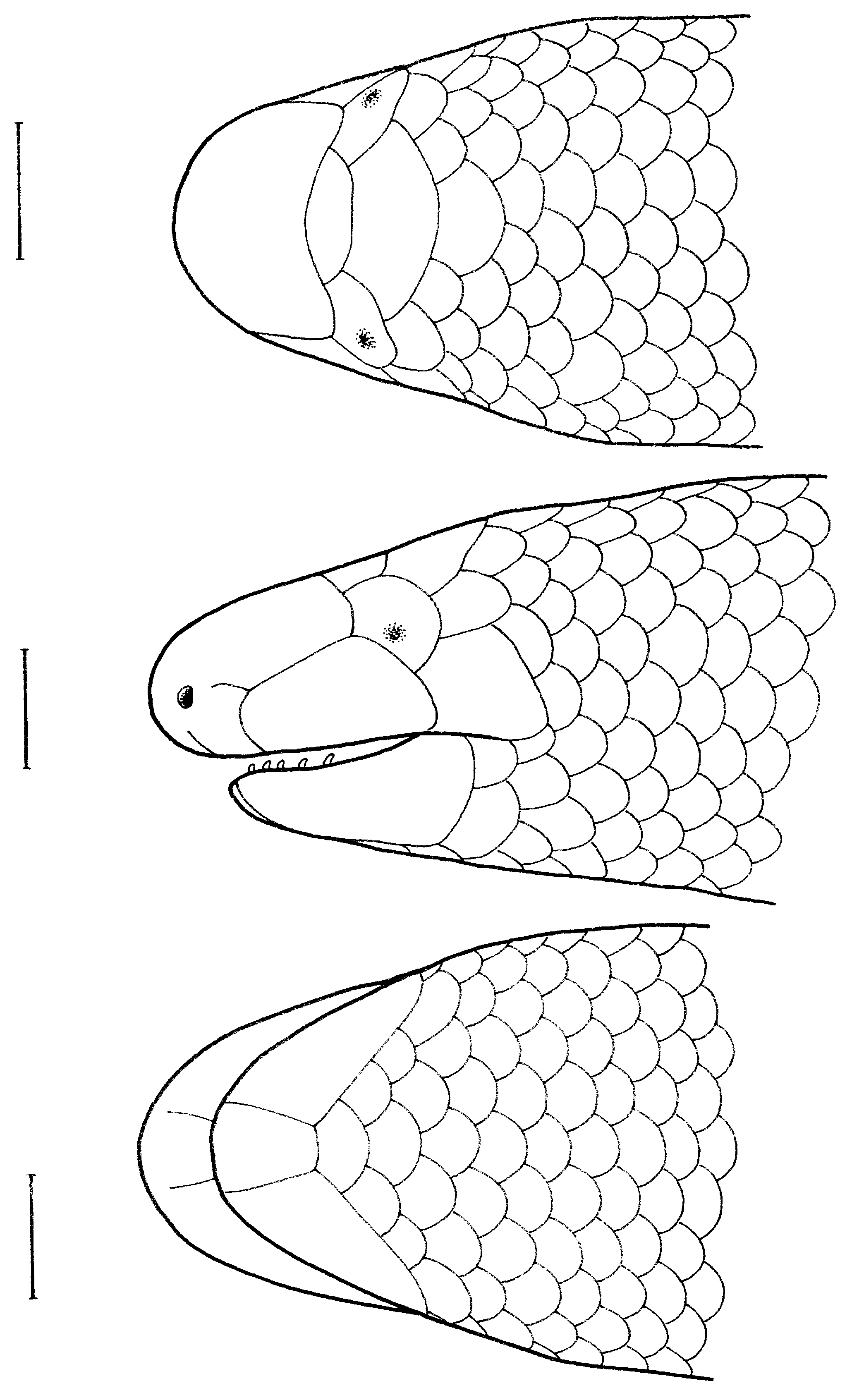
( Figs. 1-2 View Fig View Fig )
Material examined. – Holotype – ZRC 2.5368 View Materials ., female, Batu Gua Madu ( 04º 50’ 14.3” N; 101º 56’ 58.7”E), alt. 121 m ASL, near the town of Gua Musang, Kelantan State, Peninsular Malaysia, coll. N. S. Yaakob, I. Das & B. L. Lim, 19 Oct.2001. GoogleMaps
Paratype – ZRC 2.1944 View Materials , adult female, same locality as holotype, coll. M. W. F. Tweedie, Aug.1939 GoogleMaps .
slightly distinct from neck and from body; tail short (TL/ SVL ratio 0.13), tip rounded, slightly bulbous, tail base thick (TW 3.5 mm; TW/TL ratio 0.28), wider than rest of tail; body scales smooth, subhexagonal, except near preanal region, where they are subcycloid; transverse scale rows immediately posterior to head 24; at midbody 20; and just Diagnosis. – SVL to 102.7 mm; TL to 9.7 mm; TL/ SVL % 9.4-13.0; postocular single; rostral suture absent; interparietal posteriorly bordered by four slightly smaller nuchal scales; supralabial one, bordering ocular ventrally; scales bordering posterior edge of infralabial four; ventrals 180-209; subcaudals 24-39; presacral vertebrae 113-120; postsacral vertebrae 11-25; and dorsum and venter brown with a pale neck band.
Description of holotype. – SVL 93.5 mm, TL 12.3 mm; snout bluntly rounded (IN/IO ratio 0.47), projecting beyond jaws; nostril laterally oriented, oval, situated closer to snout-tip than to orbit (E-N/E-S ratio 0.80); head short, shorter than wide, HL 2.1 mm, HW 3.3 mm (HL/HW ratio 0.64), slightly flattened, HD 2.5 mm (HL/HD ratio 0.84); rostral pad with a large number of evenly distributed sensory papillae; rostral suture absent, nasal suture incomplete, extending from ocular to slightly beyond half the distance to nostril; labial suture joining nasal suture behind nostril; posterior border of rostral curved; both frontonasal and frontal wider than long, width of former 1.1 mm, of the latter 1.5 mm; interparietal single, longer than frontonasal, posteriorly bordered by four slightly smaller nuchal scales; postocular single; supralabial single, elongate, bordering ocular ventrally; infralabial lanceolate, 2.3 mm in length (infralabial length/HW ratio 0.70), infralabials separated by a smaller, trapezoid mental; scales bordering posterior edge of infralabia after postmental three bilaterally; ear opening absent; eyes dimly visible through ocular; tongue short, undivided anteriorly, pointed; teeth small, acute.
Body vermiform, BW 3.4 mm (BW/SVL ratio 0.04); head
anterior to vent 20; ventrals 180; subcaudals 39; presacral vertebrae 113; postsacral vertebrae 25; limbs absent; a single enlarged median scale in preanal region, overlapped by scales on sides; preanal pores absent; postanal scales not reduced.
Colouration. – In life, dorsum brownish-red, scales darkedged; snout-tip paler; nape collar cream. In preservative, dorsum pale brown, each scale edged with dark brown; venter slightly paler; snout-tip and throat pinkish-white; anal region cream; nuchal band cream.
Variation. – The paratype ( ZRC 2.1944 View Materials ) , a female (lacking hind limb flaps that characterize males within the genus), measures 102.7 mm in SVL, 9.7 mm in TL; TL/ SVL % 9.4%. It shows the following details of squamation and osteology: transverse scale rows at midbody 23; ventrals 209; subcaudals 24; presacral vertebrae 120; and postsacral vertebrae 11. Other characteristics including scale counts as in the holotype .
Although the subcaudal counts of the paratype (24) appear relatively lower (61.5%) than that for the holotype (39), similar, altbeit less dramatic differences are known within the Anelytropsis and Dibamus . For instance, Greer’s (1985) Table 4 provided subcaudal counts for all species then known, the lower count 77-96.7% of the upper.
Etymology. – The species name honours Dr. Lim Boo Liat, pioneering Malaysian zoologist, colleague and friend.
Natural history. – Gua Madu is a large rock shelter, adjacent to the town of Gua Musang and close to the ancient Chinese settlement of Pulai, an early Neolithic site representing the Hoabinhian culture, similar to that of Hoa Binh of Indo-China ( Chasen, 1940).
The holotype was taken ca. 3 cm from under the soil surface, ca. 15 cm from a limestone cliff. The soil at the site of collection was moist, reddish-brown in colour, and covered with fresh and dried leaves. The area is a recreational site for its impressive limestone formation, and is hemmed in by plantations of banana and cocoa. Tweedie’s (1954) specimen which is here designated the paratype of the new species was dug up while excavating a rock shelter in August 1939.
When picked up or otherwise molested, it exhibited death feigning for up to four minutes, the body curved into a circle, with the belly up and the head slightly raised. The scales of the body additionally were raised nearly perpendicular to the body, presumably through the movement of underlying muscles, producing a wrinkled appearance, somewhat similar to that commonly seen in earthworms. These wrinkles disappeared after a few minutes, presumably when the perceived threat vanished. This latter habit, thought to be a mimicry of potentially noxious earthworms that may occur in syntopy, has been observed in D. greeri from Vietnam ( Darevsky, 1992) and also in a hitherto undescribed species of Dibamus from Pulau Tioman (ID, unpubl. observ.).
Remarks. – Dibamus booliati is compared with all nominal
and one undescribed species. In showing a single postocular, Dibamus booliati , new species, can be separated from D. alfredi Taylor, 1962 (southern Thailand), D. celebensis Schlegel, 1858 ( Sulawesi, Indonesia), D. ingeri Das & Lim, 2003 (Mendolong, in Sabah, Borneo), D. kondaoensis Honda et al., 2001 (Kondao Island, formerly Pulau Condore, Vietnam), D. novaeguineae Duméril & Bibron, 1839 (New Guinea, Sulawesi, Philippines archipelago, and also apparently, Simuelue, in western Indonesia), D. smithi Greer, 1985 (Langbian Plateau, Vietnam), D. seramensis Greer, 1985 (Seram, Indonesia), D. taylori Greer, 1985 (Sumba, Lesser Sundas, Indonesia), D. vorisi Das & Lim, 2003 (Danum Valley, in Sabah, Borneo), and an undescribed species from Pulau Tioman, Pahang State, West Malaysia (illustrated by Lim & Lim, 1999; and Manthey & Grossmann, 1997), all of which have paired postoculars. The absence of rostral suture separates the new species from D. bourreti Angel, 1935 (Tam Dao, Vietnam), D. deharvengi Ineich, 1999 (Binh Châu, Vietnam), D. nicobaricus Fitzinger in: Steindachner, 1867 ( Nicobar Islands, India; see below) and D. somsaki Honda et al., 1997 ( Chanthaburi Province, Thailand). The subcaudal scale counts of 24-39 and presence of a pale nuchal band differentiate it from two remaining congeners, D. leucurus (Bleeker, 1860) (Sumatra, Borneo and the Philippines Archipelago) and D. montanus Smith, 1921 (Langbian Plateau, Vietnam).
Honda et al. (2001) did not include Dibamus nicobaricus in their key to the genus, arguing that Das’ (1996) redescription and revalidation of the species was based on material possibly not conspecific with the name-bearing type. To support their argument, they provided the diagnosis for D. leucurus (Bleeker, 1860) , apparently derived from Greer (1985), as relevant for the type of D. nicobaricus , that gives a range for the transverse rows of midbody scale counts as 20-23 and subcaudal counts of 41-52. Yet, Fitzinger’s (in Steindachner, 1867) Rhinophidion nicobaricum was based on the holotype NMW 23461 and the midbody scale count of the original description was specified as “…circa 23 Längsreihen (in Mitte der Köperlänge..)” and the subcaudal count is 36. In fact, in Greer’s (1985) review of the genus, there was some hesitation in synonymising the Nicobarese population with leucurus (presumably for the fact that only a single specimen- the NMW holotype- was examined). Based on the diagnosable characters separating the Nicobarese populations from leucurus (including complete vs incomplete rostral sutures; subcaudals 31-39 vs 41-52; snout acute vs rounded; lanceolate vs triangular infralabials; and infralabial length over 75% head width vs ca. 50% head width), and the fact that only a single (but locally widespread) species has been collected from these islands, despite recent intensive surveys (see Das, 1999), D. nicobaricus warrants retention as a valid species.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.