
Alpheus formosus Gibbes, 1850
|
publication ID |
https://doi.org/10.5281/zenodo.180922 |
|
DOI |
https://doi.org/10.5281/zenodo.3501034 |
|
persistent identifier |
https://treatment.plazi.org/id/D16A1836-FFAF-FFA1-8386-FD4AFE85C0AC |
|
treatment provided by |
Plazi |
|
scientific name |
Alpheus formosus Gibbes, 1850 |
| status |
|
Alpheus formosus Gibbes, 1850 View in CoL
Figs. 1–4 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 , 7 View FIGURE 7 c–e
Alpheus formosus Gibbes 1850: 196 View in CoL ; Kingsley 1878a: 190; Heilprin 1888: 322; Heilprin 1889: 151; Coutière 1899 (for page numbers see Chace & Forest 1970); Coutière 1910: 485; Verrill 1900: 579; Rathbun 1900: 152; Rathbun 1901: 106; Verrill 1922: 84; Balss 1924: 177; Boone 1930: 166; Chace 1956: 146; Hazlett 1962: 82; Bullis & Thompson 1965: 8; Wiliams 1965: 64; Fausto-Filho 1970: 56; Hendrix 1971: 83; Chace 1972: 67; Corrêa 1972: 3; Coelho & Ramos 1972: 148; Ray 1974: 104; Abele 1976: 273; Camp et al. 1977: 25; Gore et al. 1978: 225; Christoffersen 1979: 314 (part.); Corredor et al. 1979: 32; Christoffersen 1980a: 65; Christoffersen 1980b: 39; Rodríguez 1980: 145; Fausto-Filho 1980: 113; Morgado 1980: 33; Reed et al. 1982: 768; Carvacho 1982: 18; Cubit & Williams 1983: 24; Williams 1984: 94; Lemaitre 1984: 426; Young 1984: 29; Freitas-Lins 1985: 39; Young 1986: 106; Sterrer 1986: 325; Rodríguez 1986: 143; Abele & Kim 1986: 197, 206; Kim & Abele 1988: 39, 40; Márquez 1988: 34; Markham et al. 1990: 421; Martínez-Iglesias et al. 1996: 33; Hernández Aguilera et al. 1996: 32; Martínez-Iglesias et al. 1997: 423; Hobbs 1998: 906; Santos & Coelho 1998: 73; Posey et al. 1998: 154; Christoffersen 1998: 358; Vargas & Cortés 1999: 901; Morgado & Tanaka 2001: 177; Coelho et al. 2006: 51; Manning & Reed 2006: 289.*
Alpheus formosus View in CoL -a—Knowlton & Mills 1992: 2.
Alpheus formosus sp. a—Knowlton et al. 1993: 1630.
Alpheus formosus A—Williams et al. 2001: 377.
Alpheus formosis (lap. cal.)— McClure 2005: 142, fig. 14, pl. 4, fig. A.*
Crangon formosus — Hay & Shore 1918: 384; Schmitt 1924a: 65; Schmitt 1924b: 73; Pearse 1932: 107; Schmitt 1935: 144; Schmitt 1936: 368; Schmitt 1939: 28; Pearse 1950: 150; Pearse & Williams 1951: 143.*
Crangon formosum — Manter 1933: 177.*
Alpheus Poeyi Guérin-Méneville 1857: 19 View in CoL .
Alpheus panamensis View in CoL (not sensu Kingsley 1878)— Pocock 1890: 519; Zimmer 1913: 391; Fausto-Filho 1974: 5.*
Not Alpheus formosus View in CoL — Christoffersen 1979: 315 (part., specimens from E Pacific) (= A. panamensis Kingsley, 1878 View in CoL ); Frische 2003: 38 (color photograph, = Alpheus View in CoL sp.).
*Remark: Two species were previously confused under the name “ Alpheus formosus View in CoL ”; therefore, some of the abovelisted records may actually refer to the new species and not to A. formosus View in CoL .
Type material.— USA, Florida. Neotype, ovig. female (CL 9.7), USNM 1100697, USA, Florida, Big Pine Key, Bahia Honda State Park, shallow subtidal (less than 1.5 m), under rocks, coll. A. Anker, 18 Apr 2004 [fcn 04-010].
Additional material examined.— Panama (Caribbean coast). 1 male (CL 5.6), 1 female (CL 6.0), USNM 1100699, Bocas del Toro, Isla Bastimentos, shallow subtidal ( 0.5–1 m), from coral rocks, coll. A. Anker, 21 Oct 2005 [fcn 05-103]; 1 male (CL 7.8) USNM 1100700, Isla Grande, southern shore, shallow subtidal ( 0.5– 1.5 m), from coral rubble, coll. A. Anker, 21 Apr 2006 [fcn 06-425]; 1 juvenile (CL not measured), USNM 1100701, Isla Grande, western point, from coral rocks, shallow subtidal ( 0.5–1m), coll. A. Anker and C. Hurt, 6 Oct 2005 [fcn 05-101]; 1 male (CL 6.9), USNM 1100702, same collection data as previous specimen [fcn 06-392]. Costa Rica (Caribbean coast). 3 males (CL 5.8, 6.6, 6.5), 1 ovig. female (CL 6.7), USNM 1100698, Cahuita, shallow subtidal ( 0.5–1.5 m), from rock crevices and Halimeda clumps, coll. A. Anker, I.S. Wehrtmann and L. Harris, 26–27 Nov 2005 [fcn 05-100]; 1 male (CL 10.3), MNHN-Na 16385, same collection data as previous specimens [fcn 05-102]. Aruba. 2 males (CL 5.3, 6.3), 1 female (CL 6.2), 1 ovig. female (CL 7.1), MNHN-Na 16360, Baby Beach, shallow subtidal ( 1–1.5 m), from crevices in coral rubble, coll. A. Anker, 6 Dec 2003 [fcn 03-011]. Guadeloupe. 1 female (CL not measured), MNHN-Na 13715, Le Moule, depth 2 m, coll. F. Fasquel, 29 Oct 1999; 1 male (CL not measured), MNHN-Na 16386, same collection data as for previous specimen. Brazil. 2 males (CL 6.7, 7.8), 1 ovig. female (CL 7.9), MNRJ 20755, LT 702, Atol das Rocas, between Piscina das Tartarugas and Piscina das Rocas, in pools, among calcareous algae, coll. P.S. Young, P.C. Paiva and A.A. Aguiar, 28 Oct 2000 [ 1 male dissected]; 1 male (CL 5.7), MNRJ 20756, Atol das Rocas, Piscina do Cemitério, in calcareous algae, coll. P.S. Young et al., 1 Jan 2001; 1 male (CL 9.5), 1 ovig. female (CL 9.4), MNRJ 20757, Atol das Rocas, near Pedra do Garapira, in tide pool, coll. P.S. Young, P.C. Paiva and A.A. Aguiar, 14 Oct 2000; 1 male (CL 11.0), MNRJ 20758, Atol das Rocas, Piscina das Rocas, in calcareous alga, coll. P.S. Young et al., 26 Oct 2000; 1 male (CL not measured), MNHN-Na 16359, Pernambuco, Rio Formozo, coll. A.C. Branner Hartt, Dec 1875 [specimen lacking all pereiopods]. USA, Florida. 1 male (CL 11.5), OUMNH-ZC 2007-13 -034, USA, Florida, near Marineland, 5 km south of Sebastian Inlet, shallow subtidal, coll. J.A. Baeza, 15 Jul 2007 [fcn 07-240; specimen somewhat dessicated during transport, rehydrated in EtOH]. British Virgin Islands. 1 juvenile (CL not measured), Guana Island, coll. Guana Marine Invertebrate Project, 2000 [fcn Vc 0600] [specimen missing minor cheliped, identification tentative based on length of adrostral furrows; specimen will be deposited in LACM].
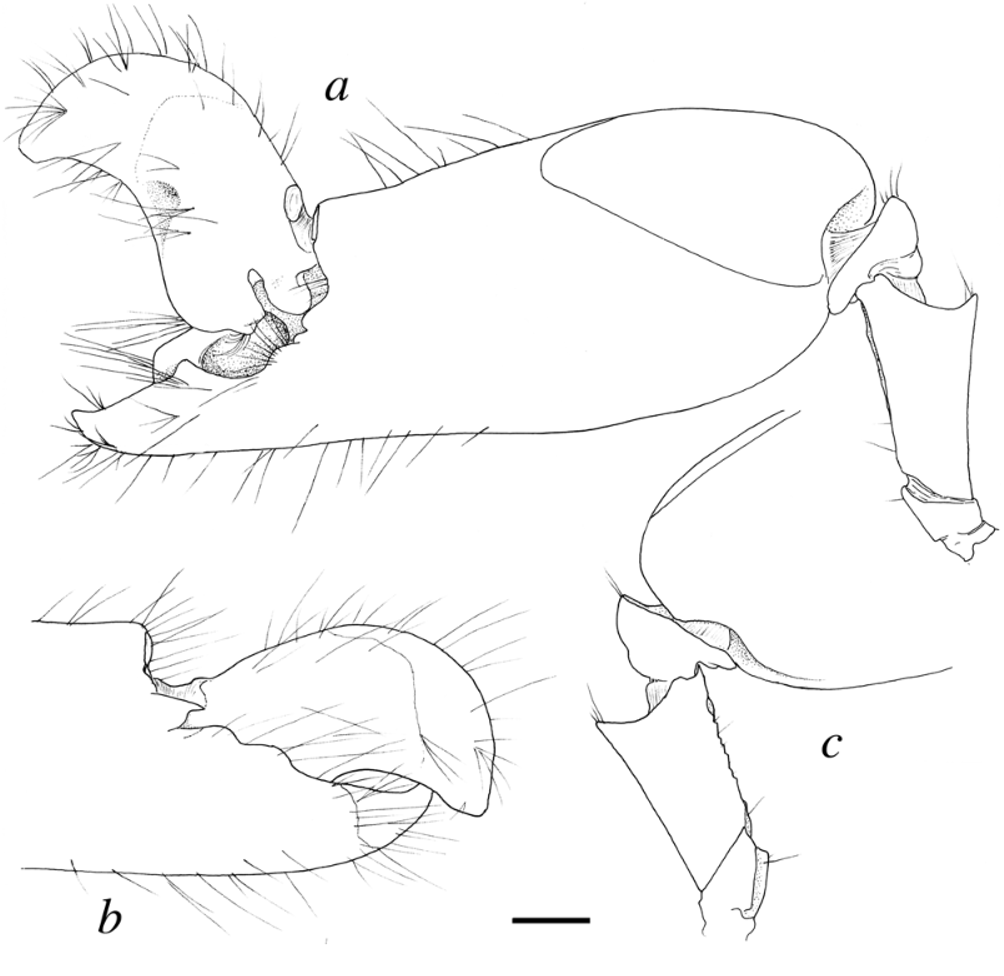
Redescription.—Carapace glabrous, non-setose, somewhat compressed laterally. Rostrum well developed, dorsally flattened; margins fringed with setae directed up- or sidewards ( Fig. 1 View FIGURE 1 a, b); tip acute, reaching to or slightly beyond distal margin of first segment of antennular peduncle ( Fig. 1 View FIGURE 1 a); rostral carina absent; orbito-rostral process broad, round W-shaped. Orbital hoods with acute teeth inserted at some distance from anterior margin ( Fig. 1 View FIGURE 1 a). Adrostral furrows deep, narrow, abruptly delimited from rostrum, posteriorly extending almost to level of cornea base ( Fig. 1 View FIGURE 1 a). Pterygostomial angle rounded, somewhat protruding anteriorly ( Fig. 1 View FIGURE 1 b, c); cardiac notch well developed. Eyes completely concealed in dorsal, lateral and partly in frontal view. Ocellar beak vertically protruding towards rostrum, more or less visible in lateral view ( Fig. 1 View FIGURE 1 c).
Antennular peduncles relatively stout, second segment longer than dorsally visible portion of first, about 1.5 times as long as wide ( Fig. 1 View FIGURE 1 a); stylocerite with acute tip, slightly overreaching distal margin of first segment ( Fig. 1 View FIGURE 1 a, b); mesioventral carina of first segment with large, bluntly protruding tooth ( Fig. 1 View FIGURE 1 c); lateral flagellum with numerous tufts of aesthetascs, accessory ramus rudimentary ( Fig. 1 View FIGURE 1 b). Antenna with stout basicerite bearing strong acute ventrolateral tooth, its tip not reaching level of stylocerite tip ( Fig. 1 View FIGURE 1 b); carpocerite stout, reaching slightly beyond distolateral spine of scaphocerite; scaphocerite with very strong distolateral tooth separated from narrow blade by deep cleft ( Fig. 1 View FIGURE 1 a), tip of distolateral tooth reaching far beyond blade and exceeding end of antennular peduncle ( Fig. 1 View FIGURE 1 a); lateral margin of scaphocerite very slightly concave at mid-length ( Fig. 1 View FIGURE 1 a).
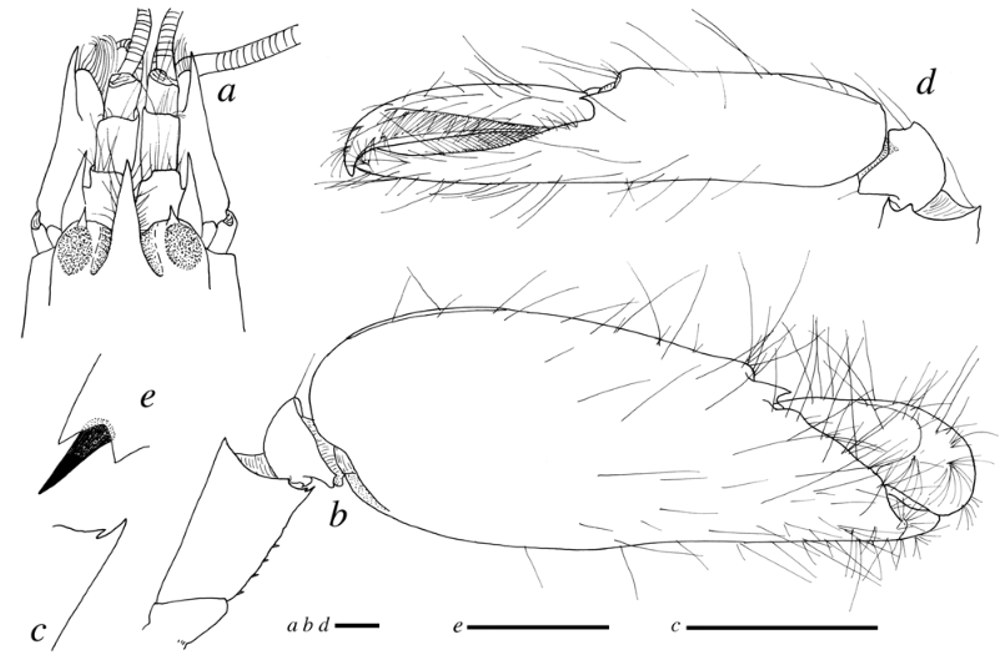
Mouthparts (mandible, maxillule, maxilla, first and second maxillipeds) typical for Alpheus . Third maxilliped moderately stout; coxa with distally subacute lateral plate; exopod reaching penultimate segment; antepenultimate segment slightly flattened, ventral margin straight; penultimate segment about 2.5 times as long as wide; arthrobranch well developed (third maxilliped generally very similar to that of A. panamensis , cf. Kim & Abele, 1988: fig. 15d, and to that of below-described new species, cf. Fig. 5 View FIGURE 5 d).
Male major cheliped ( Fig. 2 View FIGURE 2 ) with short, stout ischium; merus stout, about 2.5 times as long as wide proximally, dorsal margin distally protruding ( Fig. 2 View FIGURE 2 a, c); ventrolateral margin straight, ventromesial margin rugose, with spinules, distally with acute tooth ( Fig. 2 View FIGURE 2 c); carpus very short, cup-shaped; chela large, subcylindrical, broadly oval in cross-section; palm smooth, moderately setose distally; distomesial margin with strong acute tooth ( Fig. 2 View FIGURE 2 b); linea impressa conspicuous ( Fig. 2 View FIGURE 2 a); adhesive discs relatively large ( Fig. 2 View FIGURE 2 a); fingers more than half-length of palm, dactylus usually reaching beyond pollex, with large plunger, latter distally with stamen-shaped sensillae ( Fig. 2 View FIGURE 2 a). Female major cheliped similar to that of male, except for somewhat smaller size and different proportions of fingers to palm ( Fig. 4 View FIGURE 4 b). Mesiodistal tooth on merus sometimes ( Fig. 4 View FIGURE 4 b, c), independently of sex, with two acute teeth: one larger and stronger tooth, and one much smaller tooth.
Male minor cheliped ( Fig. 3 View FIGURE 3 ) with short, stout ischium; merus about three times as long as wide proximally; ventrolateral margin straight; ventromesial margin somewhat rugose, usually with spinules; distally with acute tooth ( Fig. 3 View FIGURE 3 d); carpus cup-shaped; chela smooth, ovate in cross-section, somewhat setose mesially ( Fig. 3 View FIGURE 3 b); distomesial margin of palm with strong acute tooth ( Fig. 3 View FIGURE 3 a, b); linea impressa conspicuous ( Fig. 3 View FIGURE 3 a); adhesive discs small ( Fig. 3 View FIGURE 3 a); fingers about as long as palm, with well developed rows of balaeniceps setae ( Fig. 3 View FIGURE 3 a–c), tips strongly curved, crossing when chela closed. Female minor cheliped similar to that of male; chela with well developed rows of balaeniceps setae ( Fig. 4 View FIGURE 4 d).
Second pereiopod with ischium subequal to merus ( Fig. 1 View FIGURE 1 d); carpus five-segmented, ratio of carpal segments (from proximal to distal) approximately: 6/3/1.5/1.5/3 ( Fig. 1 View FIGURE 1 d); chela simple, with fingers slightly longer than palm. Third pereiopod with ischium bearing small ventrolateral spine ( Fig. 1 View FIGURE 1 e); merus unarmed, about six times as long as wide; carpus unarmed; propodus with stout spines or pairs of spines along ventral margin and two spines distoventrally near articulation with dactylus ( Fig. 1 View FIGURE 1 e); dactylus about 1/3 propodus length, simple, conical, gradually curved towards acute tip, with some setae ( Fig. 1 View FIGURE 1 e). Fourth pereiopod similar to third pereiopod. Fifth pereiopod smaller and slenderer than third and fourth pereiopods; ischium unarmed; propodus with row of spines and at least eight rows of grooming setae distolaterally; dactylus conical, as in third pereiopod.
Abdominal pleura with posteroventral margins broadly rounded; sixth somite without articulated flap, dorsolateral projections on each side of telson bluntly rounded. Male second pleopod with appendix masculina reaching somewhat beyond appendix interna, with numerous slender apical and subapical setae. Uropod with protopod bearing elongate acute distolateral tooth ( Fig. 1 View FIGURE 1 f); exopod with strong acute distolateral tooth adjacent to distolateral spine; diaeresis bearing strong lateral tooth, mesial to distolateral spine ( Fig. 1 View FIGURE 1 f, g); distolateral spine stout, usually black to tan-brown ( Figs. 4 View FIGURE 4 e, 7e), sometimes pale in alcohol ( Fig. 1 View FIGURE 1 f, g); distal margins of endopod with row of spinules ( Fig. 1 View FIGURE 1 g). Telson moderately broad, subrectangular, tapering towards posterior margin ( Fig. 1 View FIGURE 1 f); dorsal surface without median groove, with two pairs of strong dorsal spines, situated at some distance from lateral margins, both pairs situated in posterior 2/3 length of telson ( Fig. 1 View FIGURE 1 f); posterior margin convex centrally, with two pairs of posterolateral spines, mesial being much longer than lateral ( Fig. 1 View FIGURE 1 f); anal tubercles well developed. Gill formula typical for Alpheus .
Size.—The largest Caribbean specimen is a male from Cahuita, Costa Rica, with CL 10.3 mm and TL 33.5 mm; the CL of remaining adult specimens range from 5.3 mm to 7.8 mm. The CL of the neotype (ovigerous female) from Florida Keys is 9.7 mm; the maximum CL of Florida specimens was 14.7 mm ( Hendrix 1971). The largest specimen from Brazil is a male from Atol das Rocas with 11.0 mm CL and 35.7 mm TL.
Color pattern.—Body red-brown to purple-brown, with more or less intense yellow mediodorsal band extending from rostrum tip to posterior margin of sixth abdominal somite; carapace with white lateral band extending from anterolateral margin to cardiac notch, continued by zigzag white band ending at sixth abdominal somite; rostrum entirely yellow, usually not bordered by reddish brown on margins; carapace flanks below white band pale; telson and uropods red-brown with large yellow patches and spots; walking legs and second pereiopods bluish, sometimes intense magenta blue; antennular peduncles yellow-brown; antennal scaphocerite brown with white areas, flagella pale bluish; chelipeds pale brown to intense orange brown, major chela with pale patch near articulation with dactylus; dactylus and pollex distally pinkish; minor chela mostly pale brown to orange brown ( Fig. 7 View FIGURE 7 c–e); distolateral spine on uropdal exopod usually brown to almost black ( Fig. 7 View FIGURE 7 e); eggs orange-reddish ( Fig. 7 View FIGURE 7 d).
Ecology.—Intertidal to about 40 m ( Sterrer 1986), commonly associated with reef-fringing habitats, in crevices of rocks, dead and living corals and coral rubble ( Verrill 1922; present study); mixed sand-rubble bottoms among turtle grass ( Thalassia testudinum ), under rocks and coral rubble; in crevices on reefs of the sabellariid polychaete Phragmatopoma sp. ( Camp et al. 1977; Gore et al. 1978); in empty conch shells ( Strombus ) on the outer fringing reefs near Thalassia beds ( Hendrix 1971); occasionally in sponges, e.g., Callyspongia , Ircinia ( Pearse 1950; Chace 1956), among bryozoan colonies, Schizoporella sp. ( Christoffersen 1979; Morgado & Tanaka 2001), and in living corals, e.g., Oculina and Porites ( Chace 1972; Reed et al. 1982); occasionally entering sea-connected cave systems ( Hobbs 1998).
Type locality.—Key West, Florida ( Gibbes 1850).
Distribution.—Western Atlantic: North Carolina ( Hay & Shore 1918; Williams 1965, 1984; Christoffersen 1979); reefs of South Carolina ( Pearse & Williams 1951); Florida: southern and central-eastern Florida (e.g., Fort Pierce, Sainte-Lucie, Hutchinson Island, Sebastian Inlet), Florida Keys (e.g., Key West, Big Pine Key, Cedar Key), Dry Tortugas ( Gibbes 1850; Coutière 1910; Zimmer 1913; Balss 1924; Boone 1930; Pearse 1932; Hendrix 1971; Camp et al. 1977; Gore et al. 1978; Reed et al. 1982; Posey et al. 1998; present study); Bermuda ( Heilprin 1888; Verrill 1922; Sterrer 1986); US Virgin Islands ( Hendrix 1971); St. John ( Zimmer 1913); British Virgin Islands: Guana (present study); Bahamas: Bimini, Cay Sal Bank ( Pearse 1950; Lemaitre 1984), Blue Holes Cave System ( Hobbs 1998); Puerto Rico; Culebra ( Rathbun 1901; Schmitt 1935; Ray 1974); French Antilles: Martinique, Guadeloupe ( Carvacho 1982; present study); Tobago ( Chace 1972); Barbados ( Zimmer 1913; Schmitt 1924b); Netherlands Antilles: Aruba, Curaçao, Bonaire ( Schmitt 1924a, 1936); present study); Venezuela: Los Roques, Tucacas, Isla Margarita, Arrecifes ( Chace 1956; Rodríguez 1980; Rodríguez 1986); Colombia: Providencia ( Schmitt 1939); Panama: Isla Grande, Colón, Bocas del Toro (Abele 1976; Cubit & Williams 1983; present study); Costa Rica: Limón, Cahuita ( Vargas & Cortés 1999; present study); Honduras: Utila (A. Anker, pers. obs.); Mexico: Quintana Roo ( Chace 1972), Veracruz, Campeche (Hernández-Aguilera et al. 1996), Puerto Morelos, Punta Estrella ( Markham et al. 1990); Jamaica: Kingston ( Zimmer 1913); Cuba: Gulf of Batabano ( Martínez-Iglesias et al. 1996, 1997); Brazil: Natal, Rio Grande do Norte, Pernambuco, Alagoas, Bahia (including Abrolhos Islands), Ceara, Espirito Santo, Rio de Janeiro, São Paulo ( Rathbun 1900; Fausto-Filho 1970; Corrêa 1972; Christoffersen 1979, 1998; Coelho et al. 2006), Fernando do Noronha ( Pocock 1890); Atol das Rocas (present study). Some of the above-listed literature records may actually refer to the below-described new species, and not to A. formosus , as the two species were not distinguished in the past.
Remarks.— Alpheus formosus is closely related to A. panamensis but can be distinguished from the latter by the orbitorostral grooves extending to the level of eye base posteriorly (vs. to the base of the orbital hoods in A. panamensis ); the rostral plate being slightly narrower at base; and the usually much darker distolateral spine on the uropod (black in A. formosus vs. pale, distally tan in A. panamensis ) (see also Kim & Abele 1988). The color of the uropodal spine may be lighter if the animal has recently molted. For instance, some alcohol-preserved specimens from Atol das Rocas, including the illustrated male specimen, have a pale spine, without trace of black pigment ( Fig. 1 View FIGURE 1 f). Most individuals, however, including the neotype, have a dark brown to blackish spine in life and in alcohol ( Fig. 4 View FIGURE 4 e). Alpheus formosus can be distinguished from the below-described new species by the presence of balaeniceps setae on the fingers of the minor chela in both sexes (more developed in males), the posteriorly more extending orbitorostral grooves and several features of the color pattern (see below).
GenBank number.— EF532605 View Materials (fcn 98-163), EF532606 View Materials (fcn 98-189), EF532607 View Materials (fcn 98-190), EF532608 View Materials (fcn 05-101, USNM 1100701), EF532609 View Materials (fcn 06-414), EF532610 View Materials (06-499), AF309903 View Materials , AF309904 View Materials .




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Alpheus formosus Gibbes, 1850
| Anker, Arthur, Hurt, Carla & Knowlton, Nancy 2008 |
Alpheus formosis
| McClure 2005: 142 |
Alpheus formosus
| Mills 1992: 2 |
Alpheus formosus
| Frische 2003: 38 |
| Christoffersen 1979: 315 |
Crangon formosum
| Manter 1933: 177 |
Crangon formosus
| Pearse 1951: 143 |
| Pearse 1950: 150 |
| Schmitt 1939: 28 |
| Schmitt 1936: 368 |
| Schmitt 1935: 144 |
| Pearse 1932: 107 |
| Schmitt 1924: 65 |
| Schmitt 1924: 73 |
| Hay 1918: 384 |
Alpheus panamensis
| Fausto-Filho 1974: 5 |
| Zimmer 1913: 391 |
| Pocock 1890: 519 |
Alpheus Poeyi Guérin-Méneville 1857 : 19
| Guerin-Meneville 1857: 19 |
Alpheus formosus
| Coelho 2006: 51 |
| Manning 2006: 289 |
| Morgado 2001: 177 |
| Vargas 1999: 901 |
| Hobbs 1998: 906 |
| Santos 1998: 73 |
| Posey 1998: 154 |
| Christoffersen 1998: 358 |
| Martinez-Iglesias 1997: 423 |
| Martinez-Iglesias 1996: 33 |
| Hernandez 1996: 32 |
| Markham 1990: 421 |
| Kim 1988: 39 |
| Marquez 1988: 34 |
| Young 1986: 106 |
| Sterrer 1986: 325 |
| Rodriguez 1986: 143 |
| Abele 1986: 197 |
| Freitas-Lins 1985: 39 |
| Williams 1984: 94 |
| Lemaitre 1984: 426 |
| Young 1984: 29 |
| Cubit 1983: 24 |
| Reed 1982: 768 |
| Carvacho 1982: 18 |
| Christoffersen 1980: 65 |
| Christoffersen 1980: 39 |
| Rodriguez 1980: 145 |
| Fausto-Filho 1980: 113 |
| Morgado 1980: 33 |
| Christoffersen 1979: 314 |
| Corredor 1979: 32 |
| Gore 1978: 225 |
| Camp 1977: 25 |
| Ray 1974: 104 |
| Chace 1972: 67 |
| Correa 1972: 3 |
| Coelho 1972: 148 |
| Hendrix 1971: 83 |
| Fausto-Filho 1970: 56 |
| Bullis 1965: 8 |
| Wiliams 1965: 64 |
| Hazlett 1962: 82 |
| Chace 1956: 146 |
| Boone 1930: 166 |
| Balss 1924: 177 |
| Verrill 1922: 84 |
| Coutiere 1910: 485 |
| Rathbun 1901: 106 |
| Verrill 1900: 579 |
| Rathbun 1900: 152 |
| Heilprin 1889: 151 |
| Heilprin 1888: 322 |
| Kingsley 1878: 190 |
| Gibbes 1850: 196 |