
Celsinotum candango, Sinev, Artem Y. & Elmoor-Loureiro, Lourdes M. A., 2010
|
publication ID |
https://doi.org/ 10.5281/zenodo.193847 |
|
DOI |
https://doi.org/10.5281/zenodo.5618337 |
|
persistent identifier |
https://treatment.plazi.org/id/CF2D87C4-FF89-FFD1-09AC-CF90FD04F9A0 |
|
treatment provided by |
Plazi |
|
scientific name |
Celsinotum candango |
| status |
sp. nov. |
Celsinotum candango View in CoL sp. n.
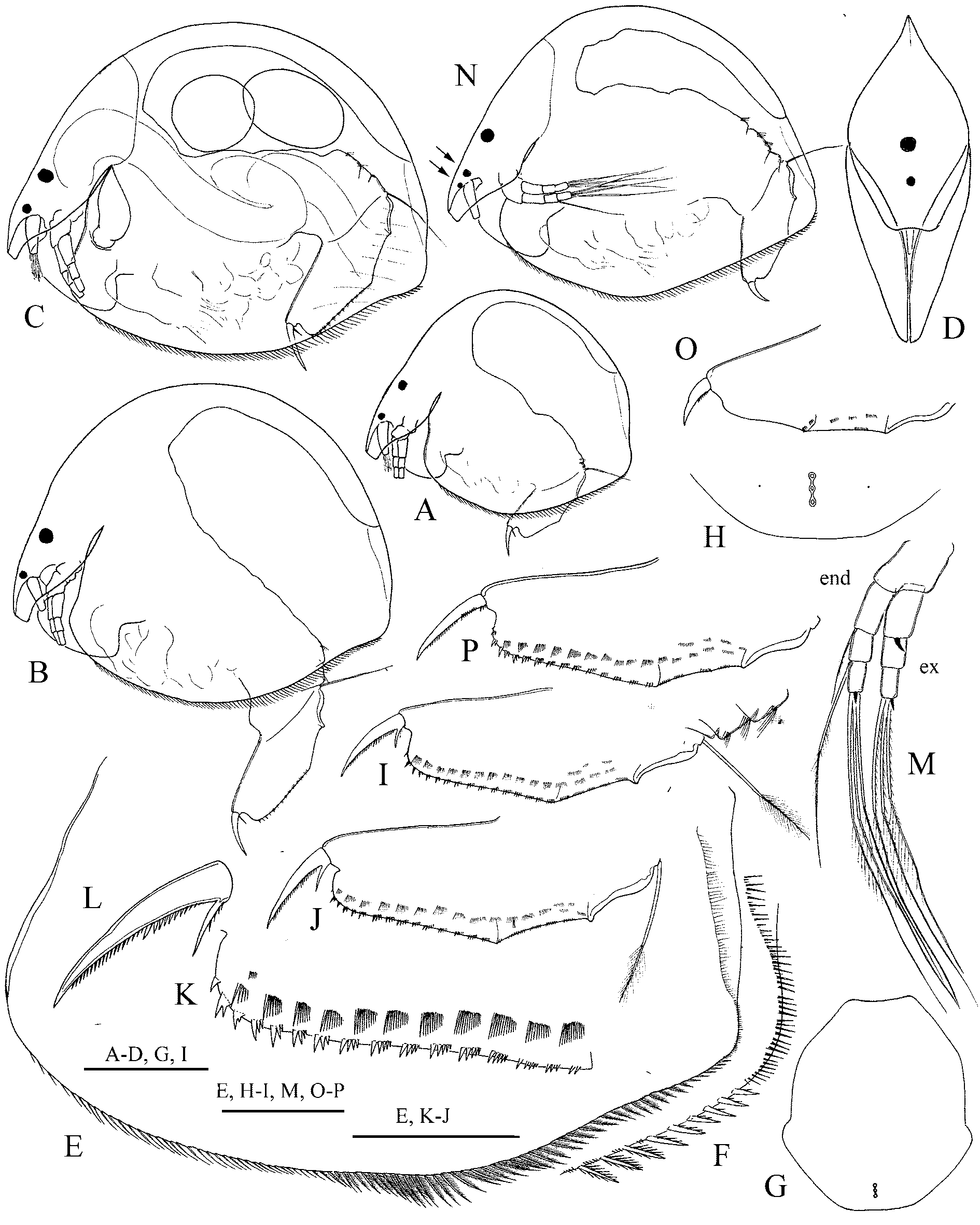
( Figs. 8–9 View FIGURE 8 View FIGURE 9 )
Etymology. From the name applied to people that built the city of Brasilia (1956–1960), and nowadays the name is used in reference to its inhabitants.
Type location. Lagoa do Henrique, a small freshwater pond situated at Brasilia National Park, Distrito Federal, Brazil (15° 41' 16.5" S, 47° 56' 22.2" W). The type series was collected on 27.v.2002 by L.M.A. Elmoor-Loureiro.
Holotype. An adult parthenogenetic female, MZUSP 19927. The label of holotype is: " Celsinotum candango sp. nov., 1 parth. Ƥ from Brasilia, DF, Brazil, HOLOTYPE ".
Paratypes. five parthenogenetic females ( MZUSP 19928), five parthenogenetic females ( MNRJ 21591), five parthenogenetic females (EL00647), six parthenogenetic females (AYS-2008-08) from the type locality.
Diagnosis. In lateral view, body high and rounded, with low dorsal keel, height/length ratio 0.8–0.75. Dorsal margin evenly arched, depression in the region of head shield weakly or not expressed. Last 5–8 setae ventral setae thick, spiniform, with only 1–3 setules on posterior margin. Head shield and head pores as for genus, lateral pores located at 1.5 IP distance from midline. Labrum as for genus.
Postabdomen of moderate length and width, narrowing distally, length about 2.5 height. Dorsal margin straight in both postanal and anal portions, distal part about 3.5 times longer than preanal one, postanal portion 2.–2.3 times longer than anal one. Preanal angle protruding, postanal angle well-defined. Postanal margin with 14–17 clusters of 2–5 small, sharp, narrow denticles in each. Length of longest denticles about 2/3 width of base of postabdominal claw. Postanal portion with 9–12 moderately broad lateral fascicles of setules, posteriormost setae of each fascicle longest, longer than nearby marginal denticles. Postabdominal claw of moderate length, 1.1–1.3 times longer than preanal portion of postabdomen. Basal spine short, about 0.2 of the claw length.
Antennule and antenna as for genus. Spine on basal segment of antenna exopodite about 1/3 length of middle segment. Thoracic limbs as for genus. Endite 1 of limb I with only two setae. Setae 4–6 of exopodite III of similar length. Epipodites IV–V with extremely large projections.
Differential diagnosis. Celsinotum candango sp. n. clearly differs from all other species of the genus in proportions of postabdomen, as postanal portion is only 1.2–1.3 times longer than anal one in all other species. It differs from Australian species ( C. hypsophilum Frey, 1991 , C parooensis Frey, 1991 , and C. platamoides Frey, 1991 ) in less developed keel, lateral head pores located close to midline (only 1.5 IP distance), longer spine on basal segment of antenna exopodite (in these species it is very small, less than 1/4 length of middle segments), and in the presence of extremely big projections on exopodites IV–V. Brazilian C. laticaudatum Smirnov & Santos-Silva, 1995 differs from C. candango in longer spine on basal segment of antenna exopodite (about 2/3 length of middle segment of antenna), in the shape of postabdomen (it is only weakly narrowing distally, distal part of postanal portion is almost rectangular) and in postabdominal denticles, in this species distalmost denticles are long and single.
Description. Parthenogenetic female. In lateral view, body high and rounded, strongly compressed laterally ( Fig. 8 View FIGURE 8 A–D), with expressed, but low, dorsal keel. Maximum height at the middle of the body, height/ length ratio 0.75–0.8 in adults. Dorsal margin evenly arched, depression in the region of head shield weakly or not expressed, posterodorsal angle not expressed, posterior margin weakly convex. Posterodorsal angle broadly rounded, with about 25 long setules of similar size, forming weakly separated groups. A row of about 50–70 long, very thin setules along posterior margin on inner side of carapace, with smaller setules between. Ventral margin convex, in some species distinctively concave in posteriormost part, straight, with 70–85 setae ( Fig. 8 View FIGURE 8 E). First 30–35 setae very short, with no recognisable setules, next 35–40 setae longer, densely setulated bilaterally, last 5–8 setae thick, spiniform, with only 1–3 setules on posterior margin ( Fig. 8 View FIGURE 8 F). Anteroventral angle rounded. Carapace ornamentation as weakly developed longitudinal lines.
Head elongated and narrow in lateral view, rostrum obliquely truncated, protruding forward and downwards. Ocellus small, eye two times larger than ocellus. Distance from tip of rostrum to ocellus larger than that between ocellus and eye.
Head shield with ridge going along the midline, maximum width behind mandibular articulation, very wide if flattened ( Fig. 8 View FIGURE 8 G). Rostrum broad, truncated. Posterior margin straight in middle portion. Three major head pores of similar size with narrow connection between them ( Fig. 8 View FIGURE 8 H). Middle pore smaller than two others. PP about 0.7–08. IP in adults. Lateral head pores at about 1.5 IP distance from midline, at level of central major head pore.
Labrum of moderate size ( Fig. 9 View FIGURE 9 A). Labral keel subtriangular, with rounded apex, height/width ratio about 2.5. Anterior margin of keel from convex to almost straight, posterior margin convex, without clusters of setae.
Thorax three times longer than abdomen. Dorsal surface of abdominal segments not saddle-shaped. No abdominal projections.
Postabdomen ( Fig. 8 View FIGURE 8 I–J) of moderate length and width, narrowing distally, length about 2.5 heights. Ventral margin straight. Basis of claws bordered from distal margin by clear incision. Distal margin from almost straight to weakly convex, distal angle broadly rounded. Dorsal margin straight in both postanal and anal portions, with distal part about 3.5 times longer than preanal one, with postanal portion 2.–2.3 times longer than anal one. Preanal angle protruding, postanal angle well-defined. Preanal margin weakly sinuous. Setae natatoriae about 2.1–2.3 length of preanal portion of postabdomen.
Postanal margin ( Fig. 8 View FIGURE 8 K) with 14–17 clusters of 2–5 small, sharp, narrow denticles in each, size of denticle increasing distally, number of denticles per cluster decreasing distally, in some specimens several distalmost denticles are single. Length of longest denticles about 2/3 width of base of postabdominal claw. Anal margin with about 5 groups of marginal setules. Postanal portion with 9–12 moderately broad lateral fascicles, posteriormost setae of each fascicle longest, longer than nearby marginal denticles. Anal portion with several smaller fascicles, spaced irregularly, in two rows. Postabdominal claw of moderate length, 1.1– 1.3 times longer than preanal portion of postabdomen ( Fig. 8 View FIGURE 8 L). Basal spine short and slender, about 0.2- of the claw length.
Antennule ( Fig. 9 View FIGURE 9 B) long and narrow, almost reaching the tip of rostrum. Antennular seta thin, of about half length of antennule, arising at 2/3 distance from the base. Nine terminal aesthetascs, two longest about 2/ 3 length of antennule itself. All aesthetascs projecting beyond anterior margin of the head shield.
Antenna relatively short ( Fig. 8 View FIGURE 8 M). Basal segment robust, branches relatively short, with basal segments two times longer than middle and apical segments. Seta arising from basal segment of endopodite thin, shorter than endopodite itself. Seta arising from middle segment o f endopodite of similar morphology with apical setae, but 1.5 times shorter. Spine on basal segment of exopodite significantly shorter than middle segment, about 1/3 of its length. Spines on apical segments short, about 1/4 length of the segment.
Thoracic limbs: five pairs.
Limb I of moderate size ( Fig. 9 View FIGURE 9 C–D). Epipodite small, rounded, without projection. Accessory seta short, about 1/4 length of ODL seta. ODL seta with thin setules in distal portion. IDL with three setae and two large clusters of stiff setules. IDL seta 1 very small, rudimentary, setae 2 and 3 setae with an annulus and thin setules in distal part, seta 3 slightly shorter than ODL seta, seta 2 about 2/3 length of seta 3. Endite 3 with four setae subequal in length. Endite 2 with three setae, longest of them longer than ODL seta. Endite 1 with two 2- segmented setae, both setulated in distal part. Endites 1 and 2 both without naked seta or sensillum on anterior face. Six-seven rows of thin setules on ventral face of limb. Two ejector hooks, one little shorter than other. Maxillar process elongated, with short setulated in distal part seta.
Limb II subtriangular ( Fig. 9 View FIGURE 9 E–G). Exopodite elongated, setulated distally, with very short seta. Eight scraping spines, three basalmost spines subequal in length, others increasing in length distally. Size of denticles on basal part of spines evenly decreasing from basal to distal spines, with the exception of spine 4, which have much thinner denticles than its neighbour. Distal armature of gnathobase ( Fig. 9 View FIGURE 9 G) with four elements, two lateral are minute, pointed, one of the middle elements geniculated, setulated in distal part, other broad, with two rows of denticles. Filter plate of seven setae, two posteriormost members 3–4 times shorter than others.
Limb III ( Fig. 9 View FIGURE 9 H–J). Epipodite oval. Exopodite elongated, subrectangular, with six setae. Seta 3 longest, setae 4–6 subequal in length, about 1/2 length of seta 3, setae 1–2 short. Setae 1–4 plumose, setae 5–6 with short thick setules in distal part. Distal endite with 3 setae, two distalmost members scraping, slender, sharp, with spinules in distal part, short bottle-shaped sensillum located between their bases; basalmost seta two times shorter, with long setules on basal side. Basal endite with 4 plumose setae increasing in size basally.
Four pointed soft setae increasing in size basally, a small bottle-shaped sensillum near the distalmost seta. Distal armature of gnathobase with four elements. First one an elongated, narrowing distally sensillum, second strongly geniculated seta, third and fourth-spines. Filter plate of seven setae of equal length.
Limb IV ( Fig. 9 View FIGURE 9 K–L). Pre-epipodite setulated; epipodite small, but with very large, curved, finger-like projection 4 times longer than epipodite itself. Exopodite rounded, with six setae. Seta 3 being longest, setae 1, 2, and 5 about 1/3 length of seta 3, setae 4 and 6 slightly shorter than seta 5. Inner portion of limb IV with four setae and small cylindrical sensillum. Scraping seta slender, with thin spinules in distal part, three slender flaming-torch seta decreasing in size basally, each with about 10 long setules. Small sensillum between the bases of middle and basal flaming-torch setae. Three soft setae slightly increasing in size basally. Gnathobase with short two-segmented seta and small hillock distally. Filter plate of five setae of equal length.
Limb V ( Fig. 9 View FIGURE 9 M). Pre-epipodite setulated; epipodite small, but with very large, curved, finger-like projection 5 times longer than epipodite itself. Exopodite subrectangular, not divided into two lobes, with four plumose setae, with length gradually decreasing basally. Inner limb portion as broad rounded lobe, with setulated inner margin. At inner face, two setae densely setulated in distal part, of same length as exopodite seta 4. Filter plate absent.
Ephippial female and male unknown.
Size. Length of single studied juvenile female 0.43 mm, height 0.36 mm. In adult female, length 0.59– 0.68 mm, height 0.47–0.54 mm.
Morphological abnormalities. Two of studied specimens have abnormalities in morphology of postabdomen ( Fig. 8 View FIGURE 8 O–P). In one specimen, postabdomen claw has no basal spine, in the other postabdomen is shortened and has deformed, shortened claw. The abnormalities in postabdomen morphology are regularly recorded for Chydoridae ( Smirnov 1971) . The latter specimen ( Fig. 8 View FIGURE 8 N) also had additional pigment spot in the region of ocellus, but the study of appendages revealed no other differences from the normal specimens of C. candango . Such abnormality was recorded only once, for Pleuroxus —the specimen with similar pigment spot near ocellus was described as a species, Pleuroxus triocellatus Smirnov, 1989 . Our data strongly questions the validity of P. triocellatus .
Distribution and ecology. At present, C. candango was only reported from the type locality: Lagoa do Henrique ( Brasília National Park), which is a small (diameter about 200m) shallow freshwater pond, with macrophytes almost all over its area, surrounded by typical "Cerrado" vegetation (tropical savanna). The sample was taken at the beginning of the dry season being the depth at sampling site of about 20–40 cm. Waters were oligotrophic low mineralized, acidic (pH: 4,93–5,50) and relatively low oxygenated (OD: 3,63– 6,32 mg/L).
Taxonomic notes. The genus Celsinotum was described by Frey (1991) for three endemic species of West Australia, inhabiting temporary saline water bodies. The fourth species of the genus, C. laticaudatum , was described by Smirnov & Santos-Silva (1995) from Central Amazonia. The description was far from complete, and no information about the trunk limbs, save for the IDL morphology, was provided. Due to the gross ecological differences from Australian species, and different morphology of postabdomen, position of C. laticaudatum within the genus seemed doubtful. Finding of Celsinotum candango leave no places to such doubts. The limb morphology of the species is quite similar to that reported for Celsinotum parooensis (see Frey 1991). Australian and American Celsinotum species significantly differs in one very important ecological feature—while Australian species inhabit temporary brackish and saline waters, American species are freshwater.
Our data confirms connections between the South American and Australasian faunas of Aloninae, already reported by several authors ( Santos-Wisniewski et al. 2001; Sanoamuang 1998; Sinev & Sanoamuang 2007). Such connections support the ancient origin and relict nature of aforementioned genera and of modern cladoceran fauna in general (Korovchinsky 2006).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Genus |