
Megalomma Johansson, 1925
|
publication ID |
https://doi.org/ 10.1080/00222930802140186 |
|
persistent identifier |
https://treatment.plazi.org/id/CE59AB25-EB09-4504-FE75-1ADB0D08586A |
|
treatment provided by |
Carolina |
|
scientific name |
Megalomma Johansson, 1925 |
| status |
|
Genus Megalomma Johansson, 1925 View in CoL emended
Branchiomma Claparède 1869: p.162 View in CoL , p.163, junior homonym (non Kölliker 1858). Megalomma Johansson 1925: p.9 View in CoL –10. Johansson 1927: p.130. Perkins 1984: p.351– 352. Fitzhugh 1989: p.76. Knight-Jones 1997: p.314. Fitzhugh 2003: p.107. Tovar- Herna«ndez and Salazar-Vallejo 2006: p.39–40.
Type species
Branchiomma köllikeri Claparède, 1869 , a junior synonym of Amphitrite vesiculosum Montagu, 1815 .
Diagnosis
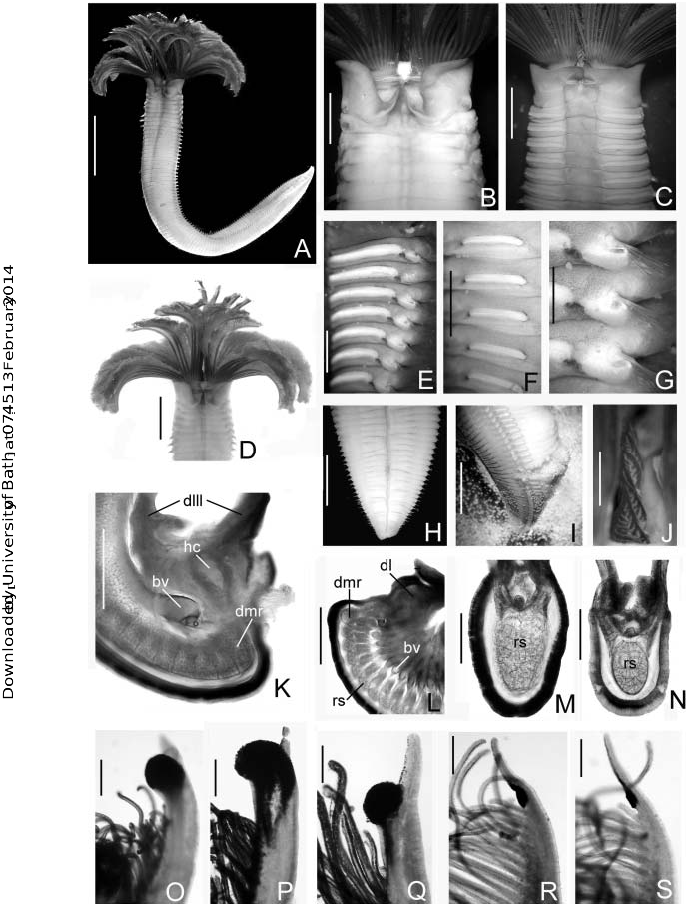
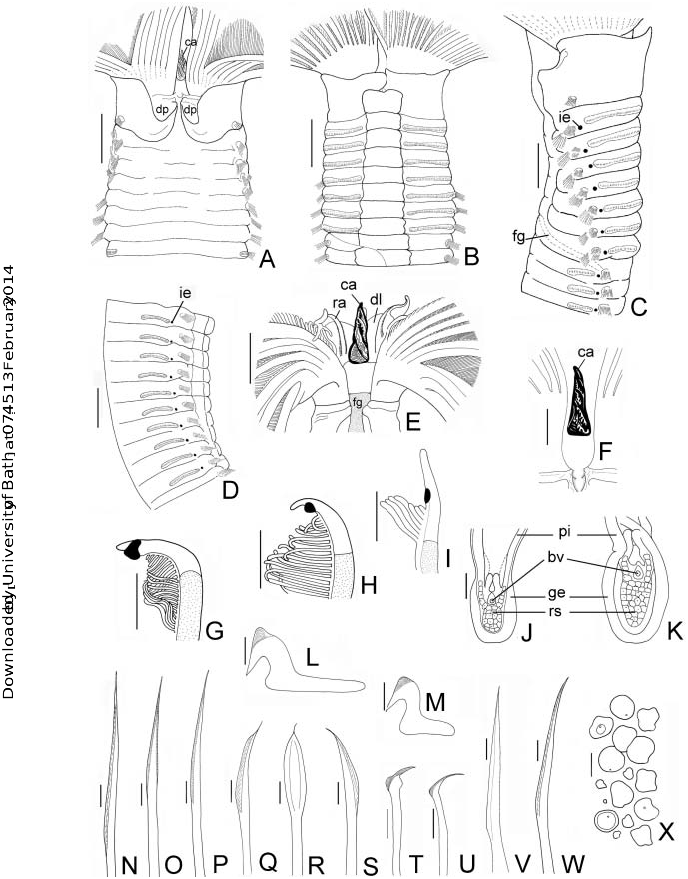
Medium- to large-sized species with numerous radioles; radiolar skeleton with up to 16 rows of cells; dorsalmost pair of radioles notably longer than the remaining radioles. Palmate membrane and radiolar flanges absent. Outer surface of radioles rounded. One or more pairs of radioles each with a single, sessile, compound eye situated along the inner radiole margin near the distal end. Dorsal lips triangular, radiolar appendages present, indicated by longitudinal ridge or ‘‘mid-rib’’ along entire length; distinct blood vessel and hyaline cartilage extending through length of appendages, without a branchial skeleton extension; dorsal pinnular appendages present or absent. Ventral lips and parallel lamella present. Caruncle can be present. Posterior peristomial ring collar present. Interramal eyespots can be present. Inferior thoracic notochaetae arranged in two or more transverse rows; they can be short, broadly hooded chaetae, near spatulate or long faintly hooded. Abdominal neurochaetal fascicles with two transverse rows of elongate, narrowly hooded chaetae. Thoracic uncini avicular; numerous, very small equal-sized teeth above the main fang; breast well developed; handles of medium length. Companion chaetae with very thin, tear-drop-shaped distal ends situated perpendicular to shaft. Abdominal uncini similar to thoracic ones, but with shorter handles.
Remarks
Johansson (1925) recognized that Branchiomma köllikeri Claparède, 1869 , did not correspond to Branchiomma Kölliker, 1858 , and proposed Megalomma as a replacement name (nomen novum) for a junior homonym. Thus, he stated ( Johansson 1925: p.9–10): ‘‘Als Kölliker sie aufs neue 1853 erwähnte, benannte er sie Branchiomma dalyelli . Der Gattungsname Branchiomma muss somit für diese Art festgestellt werden. Als Claparède 1869 für seine Art Köllikeri die Gattung Branchiomma bildete, war der Name schon präokkupiert. Die Gattung Branchiomma Claparède muss also einen neuen Namen erhalten. Ich schlage Megalomma vor, welcher Name wie Branchiomma auf die grossen zusammengesetzten Augen dieser Gattung hindeutet, doch ohne ihren Charakter als Branchialaugen hervorzuheben – die ‘‘Kiemen’’ sind ja doch keine Kiemen…’’. This can be translated as: Kölliker newly proposed Branchiomma dalyelli in 1853. Consequently, the generic name Branchiomma must apply to this species. When Claparède used Branchiomma for his species Köllikeri in 1869, the generic name was already in use. Therefore, the genus Branchiomma Claparède must receive a replacement name. I propose Megalomma , which, like the name Branchiomma , indicates the big compound eyes of this genus but without their character as branchial eyes, – the ‘‘gills’’ are not gills…
Later, Johansson in his magnificent monograph ( Johansson 1927) gave a diagnosis for Megalomma that he had named previously. Some authors have attributed the author/year as Johansson 1927 ( Hartman 1959; Perkins 1984; Fitzhugh 1989; Knight-Jones 1997; Tovar-Herna«ndez and Salazar-Vallejo 2006); however, the new name first published in 1925, as an indication, must be regarded as valid (ICZN 1999, Article 12.2.3).
Hartman (1959: p.549) regarded Branchiomma köllikeri Claparède, 1869 as a junior synonym of Amphitrite vesiculosum Montagu, 1815 , and she incorrectly included the latter as type species for Megalomma . However, the name of a type species remains unchanged even when it is a junior synonym or homonym, or a suppressed name, and the name of a type species should be cited by its original binomen (ICZN 1999, Article 67.1.2, Recommendation 67B).
The present definition primarily follows Fitzhugh (2003) except for the following: the type species has inferior thoracic chaetae long and slender, with hooded area, including tip, about 10 times longer than broad and only faintly hooded, whereas on most other species they are shorter and broadly hooded, nearly spatulate, such as described by Perkins (1984). In the modification provided here, we include these variations on inferior thoracic chaetae and also point out that the dorsalmost pair of radioles is notably longer than the remaining radioles; radioles are supported by up to 16 skeletal cells; a caruncle and interramal eyespost can be present.
All species of Megalomma have subdistal eyes, at least in the dorsalmost radiolar pair; usually this pair of eyes is prominent and whenever other eyes are present, this pair is the largest; one of the features provided here in the emendation points out that the dorsalmost pair of radioles is notably longer than the remaining radioles. Most original descriptions in Megalomma refer to the size and shape of the subdistal eyes (spiralled, flattened, globular) combined with the length of the radiolar tips; but only a few have detailed the length of the dorsalmost radiolar pair: M. acrophthalmos ( Grube 1878) ; M. claparedii ( Gravier 1908a) ; M. lobiferum ( Ehlers 1887) ; M. pacifica Johansson 1927 ; M. perkinsi Tovar-Herna «ndez and Salazar-Vallejo 2006; M. quadrioculatum ( Willey 1905) and M. suspiciens ( Ehlers 1904) . For others, the enlarged radiolar pair has been illustrated though not described: M. bioculatum ( Ehlers 1887, Pl. 53, Figure 1 View Figure 1 ); M. miyukiae Nishi 1998 ( Figure 1 View Figure 1 ); M. quadrioculatum ( Willey 1905: Pl. 7, Figure 168) and M. vigilans ( Claparède 1870: Pl. 14, Figure 3 View Figure 3 ). Furthermore, the epithelium surrounding the compound eye of the dorsalmost radiolar pair in M. coloratum ( Chamberlin 1919) and M. circumspectum ( Moore 1923) from the Mexican Pacific is very thick; this character should be examined in the other species of Megalomma .
All radioles, including the dorsalmost radiolar pair, are supported by 12–16 skeletal cells in cross-section ( Figures 3 View Figure 3 A–C, see also Brunotte 1888, Pl. 1, Figure 24) and their internal construction is identical to that in other sabellin genera: a blood vessel, a coelomic chamber, a pair of nerves, radiolar and pinnular skeleton, muscle, cartilage matrix, ciliated epithelium and cuticle ( Evenkamp 1931; Orrhage 1980; Bick and Randel 2005; Tovar-Herna«ndez and Sosa-Rodrı«guez 2006).
Personal observations on M. lobiferum and M. heterops Perkins, 1984 , in aquaria and in natural coral reef habitats from several localities in the Mexican Caribbean Sea, suggest that the dorsalmost radiolar pair and their eyes have a ‘‘periscope’’ role and are always erect. When these worms are disturbed they have a rapid withdrawal reaction; later the tip of the dorsalmost radiolar pair lean out the mouth of the tube, then this pair becomes completely extended, and finally the whole branchial crown is exposed again. Both the enlarged size and the thick epithelium of the dorsalmost pair can probably be explained by the fact that this pair in particular needs to provide support for the large eyes, and rigidity and flexibility to facilitate a rapid withdrawal reaction. These functions are possibly combined with hydrostatic mechanisms, and the giant axons in coordination with muscular contractions as has been shown in other sabellins ( Kryvi 1971; Mellon et al. 1980; Koechlin 1981).
The third character included in the generic emendation corresponds to the caruncle. This structure was recorded or illustrated for seven species: M. lobiferum ( Ehlers 1887) from Florida, M. suspiciens ( Ehlers 1904, Figure 2 View Figure 2 ) from New Zealand, M. quadrioculatum ( Willey 1905, Pl. 7, Figure 168) from Sri Lanka ( Ceylon), and a species of Megalomma (as Branchiomma and possibly M. vesiculosum ) from L’E« tang de Thau, southern France ( Brunotte 1888, Pl. 1, Figure 31, Pl. 2, Figure 35). Recently, Tovar-Herna« ndez and Salazar-Vallejo (2006) recorded the presence of a caruncle in topotypes of M. pigmentum Reish, 1963 (described from San Quintı«n Bay, Baja, CA); and a species referred as Megalomma sp. 1 (from Venezuela). The new species described below from the Eastern Tropical Pacific also has a caruncle; it is covered with four ciliated bands.
Interramal eyespots have only been recorded for Megalomma nechamae Knight- Jones, 1997, M. suspiciens , and the new species described below. These segmental ocelli have only otherwise been recorded for species in the genera Branchiomma , Pseudobranchiomma and Bispira ( Fitzhugh 1989) .
The presence of a caruncle, the long dorsalmost radiolar pair, the high number of radiolar skeletal cells and the interramal eyespots has prompted this modification of the definition of the genus. These new features should be taken into account in new taxonomic studies because they could provide additional information for analysis of phylogenetic relationships.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Megalomma Johansson, 1925
| Marı, Tovar-Herna, a Ana, ndez & Salazar-Vallejo, Sergio I. 2008 |
Megalomma
| Johansson 1925: 9 |
Branchiomma Claparède 1869 : p.162
| Claparede 1869: 162 |
Branchiomma köllikeri Claparède, 1869
| Claparede 1869 |
Amphitrite vesiculosum
| Montagu 1815 |