
Megalomma carunculata, Marı & Tovar-Herna & ndez & Salazar-Vallejo, 2008
|
publication ID |
https://doi.org/ 10.1080/00222930802140186 |
|
persistent identifier |
https://treatment.plazi.org/id/CE59AB25-EB05-4502-FE29-1C5F0EA05DDF |
|
treatment provided by |
Carolina |
|
scientific name |
Megalomma carunculata |
| status |
sp. nov. |
Megalomma carunculata View in CoL sp. nov.
Material examined
Type material. ( LACM–AHF POLY 2196, holotype ♀) Mexican Pacific, Guerrero, Acapulco, Punta Manzanillo, Coll. S. I. Salazar, 4 August 1988, 1 m.
Non-type material. ( UMAR–POLY) Mexican Pacific, Oaxaca, Huatulco, El Maguey , Coll. S. Rodrı «guez, 4 July 2007 (one ♀); Puerto A « ngel, Aguete, Coll. G. Cancino, 25 August 2006, 1.5 m (one); Puerto A « ngel, La Boquilla, Coll. J–B, G–N, 23 April 2005 (one) .
Additional material. Megalomma pigmentum Reish, 1963 ( ECOSUR) Mexican Pacific, Bahı«a San Quintı«n, Coll. L. E. Caldero« n, 8 December 1981. Megalomma sp. 1 Tovar-Herna«ndez and Salazar-Vallejo, 2006 ( USNM 57945) Venezuela, Bahı«a de Mochima, Cumana«, Coll. Edwards, 22 June 1971, 6– 10 m, in calcareous sand (seven). Megalomma lobiferum ( Ehlers, 1887) (ECOSUR) Mexican Caribbean, I.
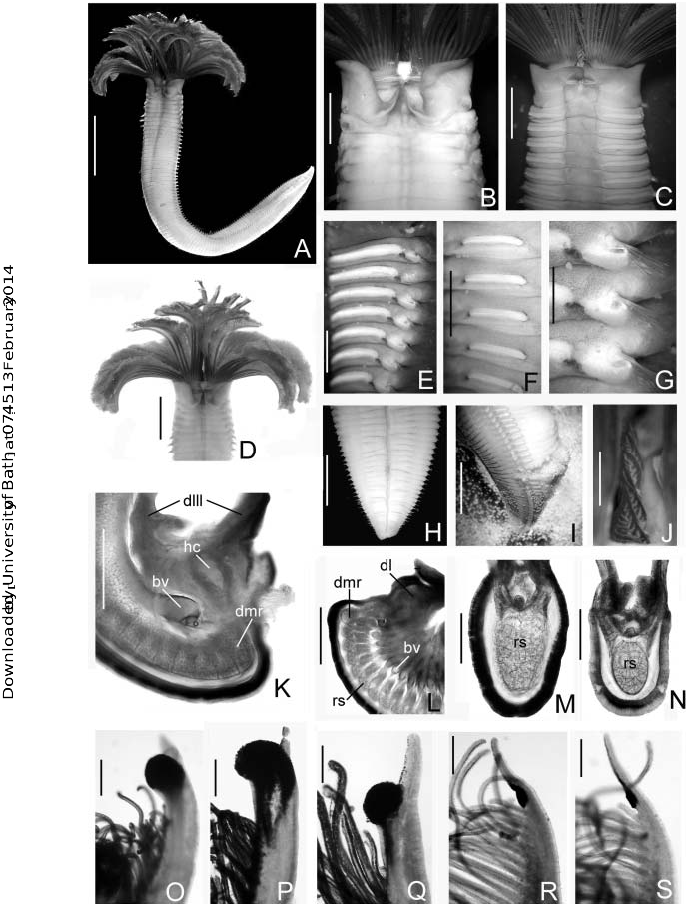
view, (F) anterior abdominal segments, lateral view, (G) thoracic chaetigers, (H) posterior abdomen, ventral view, (I) oocytes in posterior abdomen, (J) caruncle, dorsal view, (K) base of the dorsal lip, transverse section, (L) base of the right branchial lobe, transverse section, (M) base of the dorsalmost radiole, transverse section, (N) base of a lateral radiole, transverse section, (O,P) tips of the dorsalmost radioles, (Q) tip of a lateral radiole, ( R) tip of a ventral radiole, (S) tip of the ventralmost radiole. (A–D,H,J,M–O, R,S) Holotype LACM-AHF; (E– G,I,K,L,P,Q) UMAR-POLY, Huatulco. Scale bars: (A) 20 mm, (B,C,H,I) 5 mm, (D) 10 mm, (E,F) 2.5 mm, (G,J) 1.25 mm, (K,L) 1.3 mm, (M–S) 0.15 mm.
Contoy, Punta Sur, Coll. S. I. Salazar, 2 March 2001 (one). Veracruz, I. Verde, Coll. S. I. Salazar, 5 August 1985 (one). Panama, Fuerte Sherman, Colo« n, Coll. S. I. Salazar, 2 June 2002, 30 ° C, 30 ups (one).
Description
(Based on holotype; variation of non-type material in parentheses)
Colour, body shape and size. Branchial crown, dorsal lips and ventral lappets purple; caruncle with purple helicoidal bands; body cream-coloured. Radioles purple with two white bands distributed over outer and lateral radiole margins and adjacent pinnules, bands located about three-quarters of the way along crown, each band occupies a space of 20–22 pairs of pinnules. Radiolar tips white ( Figure 1A View Figure 1 ). Thoracic and abdominal segments with interramal purple eyespots (ie): large in thorax ( Figures 1E,G View Figure 1 , 2C View Figure 2 ) then decreasing towards posterior abdomen ( Figures 1F View Figure 1 , 2D View Figure 2 ). Total branchial crown length 22 mm (18). Body depressed, total thoraxabdomen length 93 mm (54–82 mm), maximum width 11 mm (8–10 mm) throughout most of thorax. Tube rigid, covered with fine sand grains, tube wall thickness 3 mm, diameter 13 mm.
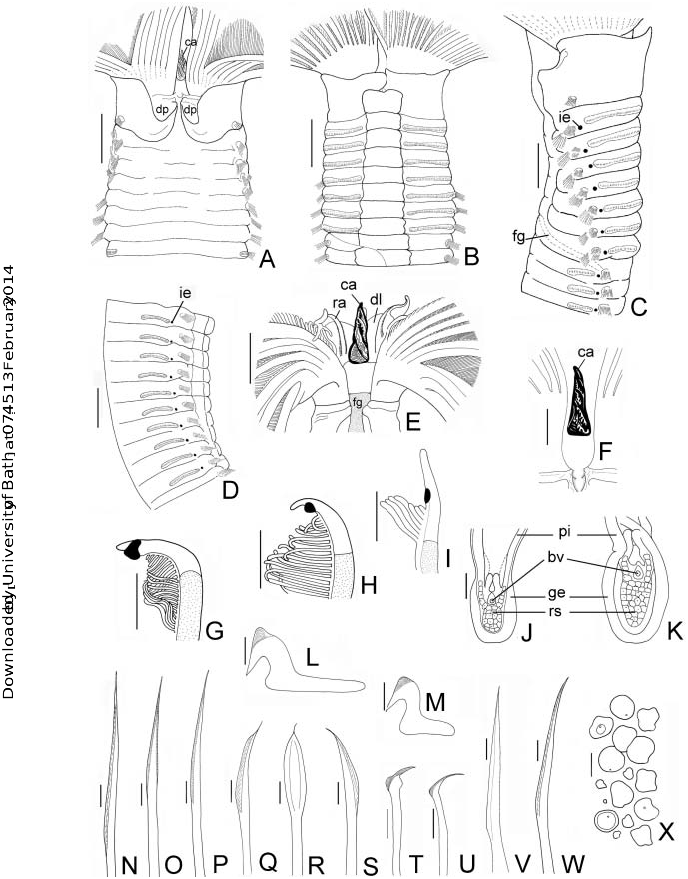
Branchial crown. Branchial lobes semi-circular. Forty-two pairs of radioles (32). Outer surfaces of radioles rounded ( Figures 1M,N View Figure 1 , 2H,I View Figure 2 ). Radiolar skeleton axis composed of 12–16 rows of cells ( Figures 1M,N View Figure 1 , 2J,K View Figure 2 ). Sub-distal compound eyes present in all radioles. Dorsalmost pair of radioles erect, rigid with rachis notably thick in lateral view ( Figures 1 View Figure 1 K–M, 2G); eyes distinctly larger, spirally arranged around radioles ( Figures 1O,P View Figure 1 , 2G View Figure 2 ); radiolar tips short filiform; tip of the dorsalmost radiolar pair with a thick epithelium near the compound eye. Second to fifth dorsalmost pairs of radioles erect, turgid with medium-sized filiform radiolar tips ( Figures 1D View Figure 1 , 2H View Figure 2 ), rounded eyes distinctly smaller than dorsalmost ones ( Figures 1Q View Figure 1 , 2H View Figure 2 ). Lateral and ventral radioles with subspherical small eyes, tips long, filiform ( Figures 1R,S View Figure 1 , 2I View Figure 2 ). Pinnules long along basal third-quarter of radiolar length, then decreasing gradually towards the compound eye. Dorsalmost pair of radioles with proximal pinnules twice as long as distal pinnules. Dorsal lips (dl) erect, triangular about one-quarter the length of the branchial crown, with radiolar appendages (ra) and two long lateral lamellae (dlll) ( Figure 2E View Figure 2 ) but without branchial skeleton. Instead, the dorsal lips supported with hyaline cartilage (hc) ( Figure 1K View Figure 1 ). Ventral lips about one-quarter as long as dorsal lips, broadly rounded. Ventral parallel lamellae short, completely concealed by ventral collar margin. Caruncle (ca) long, erect, triangular, placed above the mouth, between dorsal lips ( Figures 1J View Figure 1 , 2E,F View Figure 2 ).
radiole, (J) third ventral radiole, transverse section, (K) dorsalmost radiole, transverse section, (L) thoracic uncinus from second segment, (M) abdominal uncini from segment 10, (N) elongate narrowly hooded chaeta from collar, (O,P) superior narrowly hooded thoracic chaetae, (Q–S) inferior thoracic chaetae broadly hooded, ( T,U) companion chaetae, ( V,W) elongate, narrowly hooded abdominal chaetae, (X) oocytes. (A–X) Holotype LACM-AHF. Scale bars: (A–E) 5 mm, (C,D) 4 mm, (E) 2.5 mm, (F) 1.25 mm, (G–K) 0.15 mm, (L–N,P–W) 0.18 mm, (O) 0.7 mm.
Peristomium . Mid-dorsal collar margins not fused to faecal groove, forming a broad gap. Dorsal pocket (dp) well developed, ‘‘U-shaped’’ ( Figures 1B View Figure 1 , 2A View Figure 2 ). Ventral margin of collar incised forming two subtriangular, ventral lappets slightly overlapped ( Figures 1C View Figure 1 , 2E View Figure 2 ). Ventral shield of collar rectangular, swollen ( Figures 1C View Figure 1 , 2B View Figure 2 ). Lateral collar margins as high as dorsal margins ( Figure 2C View Figure 2 ). Notopodia in chaetiger 1 composed of superior and inferior rows of elongate, narrowly hooded chaetae ( Figure 2N View Figure 2 ); superior group distinctly longer than inferior one.
Thorax. Seven thoracic segments. Ventral thoracic shields well developed, swollen, rectangular ( Figures 1C View Figure 1 , 2E View Figure 2 ). Notopodial fascicles with a superior group of elongate, narrowly hooded chaetae ( Figures 1G View Figure 1 , 2O,P View Figure 2 ), and two inferior rows of short hooded chaetae ( Figures 1G View Figure 1 , 2 View Figure 2 Q–S). Pre- and post-chaetal lobes well developed. Neuropodial uncini with main fang surmounted by numerous rows of minute teeth, breast well developed, handles elongate ( Figure 2L View Figure 2 ), no longer than companion chaetal shaft. Companion chaetae with membranous, tear-drop shaped distal end, perpendicular to the slender shaft ( Figures 2T,U View Figure 2 ). Thoracic neuropodial tori longest on chaetiger 2, not contacting ventral shields ( Figures 1C,E View Figure 1 , 2B,C View Figure 2 ). Tori successively shorter on remaining thoracic chaetigers ( Figures 1E View Figure 1 , 2C View Figure 2 ).
Abdomen. Abdominal segments: 127 (86–122). Abdominal shields well developed, swollen, rectangular, divided by the faecal groove. Abdominal neuropodia with two transverse rows of elongate, narrowly hooded chaetae ( Figures 2V,W View Figure 2 ); chaetae in posterior row longer than those in anterior row. Abdominal notopodia with avicular uncini, main fang surmounted by numerous rows of minute teeth; breast well developed; handles reduced, less than half as long as the handles of thoracic uncini ( Figure 2M View Figure 2 ). Tori slightly shorter than those of segment 7 ( Figures 1F View Figure 1 , 2D View Figure 2 ). Pygidium short, broadly rounded, without eyespots ( Figure 1H View Figure 1 ).
Gametes. Holotype a mature female; no discrete ovaries were detected but early and fully developed oocytes were found free-floating in the coelom of the entire abdomen, apparently underwent vitellogenesis in the coelom (extraovarian oogenesis), as indicated by a change in shape (rounded and polygonal) ( Figure 2X View Figure 2 ), diameter (0.7–1.5 mm) and appearance of the cytoplasm.
Remarks
Megalomma carunculata View in CoL sp. nov. is a beautiful species, although rare in the Mexican Pacific, inhabiting tide pools. The holotype and one specimen from Oaxaca are females with abdomens full of oocytes ( Figure 1I View Figure 1 ), both collected in mid-summer. Information on the reproductive biology of species in Megalomma View in CoL is scarce. Hornell (1893) recorded the method of dispersion of gametes in M. vesiculosum ( Montagu, 1815) : towards mid-summer a female apparently ceased feeding and formed a cone with its radioles; the female gradually accumulated numerous, large ova; then, rather suddenly, the worm spread its radioles with considerable force, so that the ova shot upwards and outwards to a distance of several inches. A minute later, the worm again expanded its radioles, passing up ova again from below in rapid succession. Then when the radioles were once again clogged, a similar sudden jerk dispersed them widely. This process was repeated for three-quarters of an hour with more or less regularity. A short time after the beginning of the operation (almost simultaneously), a neighbouring individual began to manifest signs of a certain unusual excitement, and a few minutes later from the centre of its radioles, there arose a thin greyish cloud, like smoke wreaths composed of spermatozoa that were being ejected. Later, Wilson (1936) described development in M. vesiculosum , and confirmed that the species is gonochoric and a broadcast spawner, and he was also able to obtain larvae throughout the summer.
Megalomma carunculata View in CoL sp. nov., M. lobiferum ( Ehlers, 1887) View in CoL , M. pigmentum Reish, 1963 View in CoL , M. quadrioculatum ( Willey, 1905) View in CoL , M. suspiciens ( Ehlers, 1904) View in CoL , M. sp. 1 Tovar-Herna« ndez and Salazar-Vallejo, 2006, and M. ‘‘de l’E« tang de Thau’’ ( Brunotte, 1888), have been described as having a caruncle. According to the classification by Knight-Jones (1997), M. pigmentum View in CoL belongs to group 2B ( Table 1), it is characterized by the following features: anterior margin of collar not fused to faecal groove, pockets absent; compound eyes only in the dorsalmost pair of radioles; ventral lappets lanceolated, overlapped; ventral shield of collar divided into asymmetrical parts. Megalomma quadrioculatum View in CoL has the dorsal collar margins fused to the faecal groove, dorsal pockets well developed and eyes on dorsalmost, second and third pairs of radioles, it belongs to group 1B ( Table 1). Megalomma carunculata View in CoL sp. nov., M. lobiferum View in CoL , M. suspiciens View in CoL and M. ‘‘de l’E« tang de Thau’’ belong to group 1A ( Table 1); these have the anterior margin of the collar fused to the faecal groove. Megalomma carunculata View in CoL sp. nov., has subtriangular ventral lappets, overlapped (rounded, not overlapped in M. lobiferum View in CoL , M. suspiciens View in CoL and M. ‘‘de l’E« tang de Thau’’); dorsal pockets well developed, in ‘‘U-shaped’’ (less developed in M. lobiferum View in CoL , well developed and incised laterally in M. suspiciens View in CoL and M. ‘‘de l’E« tang de Thau’’); compound eyes in all radioles (on most of radioles in M. lobiferum View in CoL , M. suspiciens View in CoL and M. ‘‘de l’E« tang de Thau’’); interramal eyespots in thoracic and abdominal segments (absent in M. lobiferum View in CoL , M. suspiciens View in CoL and M. ‘‘de l’E« tang de Thau’’). The presence of a caruncle in other groups deserves some study to assess whether this character is an autoapomorphy or a plesiomorphy.
Etymology
The specific name is derived from the Latin word for caruncula, meaning a fleshy outgrowth in the body, and refers to the presence of the caruncle.
Description of the caruncle and dorsal lips
Material examined
Megalomma lobiferum ( Ehlers, 1887) (ECOSUR) Mexican Caribbean, Quintana Roo, Contoy, Punta Sur, Coll. S. I. Salazar, 2 March 2001 (one). Veracruz, I. Verde, Coll. S. I. Salazar, 5 August 1985 (one). Panama, Fuerte Sherman, Colo « n, Coll. S. I. Salazar, 2 June 2002, T530 , S530/00 (one). ( LACM–AHF, AF 00–59, Vc 0634) British Virgin Islands, Guana, Beef Island , Trellis Bay , Coll. K. Fitzhugh, 12 July 2000, tubes taken from coral rubble (two). ( LACM–AHF, AF 00–59, Vc 0565) Beef Island , Trellis Bay, directly off airport, Coll. K. Fitzhugh, 12 July 2000 (one). ( USNM 073019 About USNM ) Panama, Galeta reef, Coll. A. A. Reimer, April 1982 (six) .
Megalomma pigmentum Reish, 1963 (ECOSUR) Mexican Pacific, Bahı«a San Quintı«n, Coll. L. E. Caldero« n, 8 December 1981.
Megalomma carunculata sp. nov. ( LACM–AHF POLY 2196 , holotype) Punta Manzanillo, Acapulco, Guerrero, Mexican Pacific , Coll. S. I. Salazar, 4 August 1988, 1 m. ( UMAR–POLY) Mexican Pacific, Oaxaca, Huatulco, El Maguey, Coll. S. Rodrı «guez, 4 July 2007 (one); Puerto A « ngel, Aguete, Coll. G. Cancino, 25 August 2006, 1.5 m (one); Puerto A « ngel, La Boquilla, Coll. J–B, G–N, 23 April 2005 (one) .
Gross observations
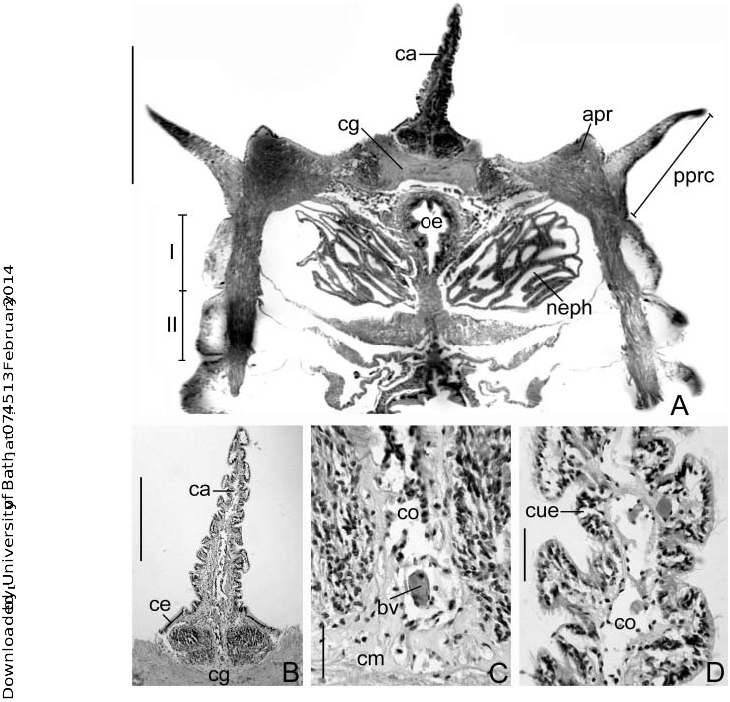
The caruncle (ca) is an erect, triangular lobe ( Figures 1J View Figure 1 , 2 View Figure 2 E–F), located dorsally above the mouth (mo), and between the dorsal lips (dl) ( Figure 2E View Figure 2 ). In fresh and old preserved material of M. lobiferum , M. carunculata sp. nov. and M. pigmentum the caruncle is purple, the length can be as long as the dorsal lips but never longer. The caruncle of M. lobiferum is covered by four ciliated bands that run longitudinally ( Figure 3A View Figure 3 ); there are one dorsal (db) ( Figure 3B View Figure 3 ), one ventral (vb) ( Figure 3C View Figure 3 ) and two lateral bands (lb): the right lateral band (rlb) ( Figure 3C View Figure 3 ) and the left lateral band (llb) ( Figure 3B View Figure 3 ). The cilia from the lateral bands are hypertrophied, wider distally, tongue-like, forming a ciliated curtain or membranella – possibly hypertrophied nuchal organs – ( Figures 3B,E,F View Figure 3 ). The dorsal band of cilia is an extension of those cilia present in the anterior peristomial ring (apr); and extends posteriorly towards the faecal groove (fg) ( Figure 3A View Figure 3 ). All bands are separated by non-ciliated narrow grooves (g) ( Figure 3C View Figure 3 ) and therefore they may not belong to the same type of sensory organs.
Internal construction
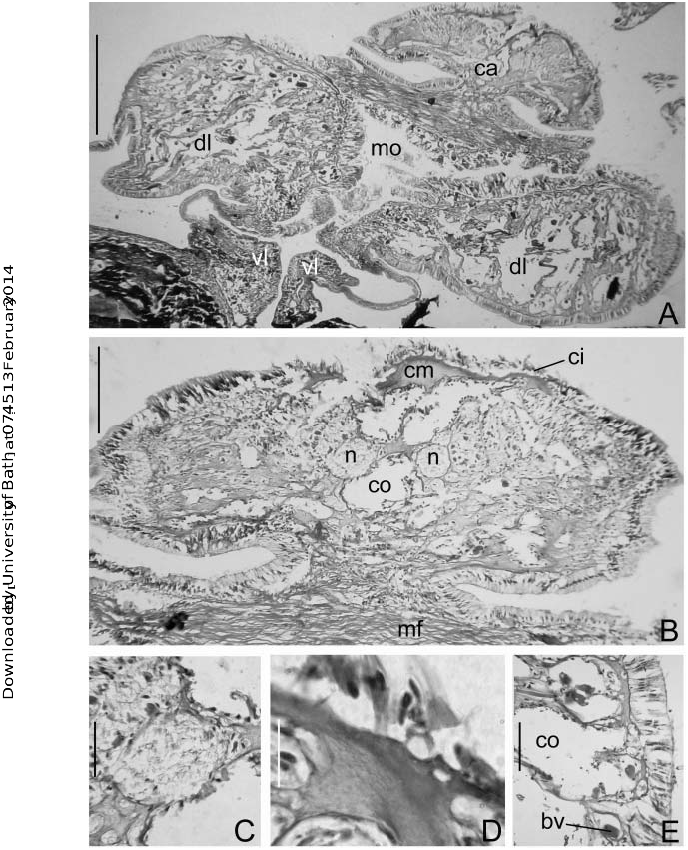
The caruncle of M. lobiferum is innervated directly from the cerebral ganglion (cg) ( Figures 4A,B View Figure 4 , 5B,C View Figure 5 ); and it is supported with hyaline cartilage constituted by a homogeneous eosinophilic matrix (cm) ( Figures 4C View Figure 4 , 5B,D View Figure 5 ). A blood vessel (bv) runs along the structure, covered by a broad coelomic chamber (co) and conjunctive tissue ( Figure 4C View Figure 4 ). Columnar epithelium (ce) is present at the base of the caruncle ( Figure 4B View Figure 4 ), and cubic epithelium (cue) with long ciliated cells (ci) towards the tip ( Figures 4D View Figure 4 , 5B View Figure 5 ). The anterior pair of nephridia (neph) crosses the coelomic cavity of the first thoracic chaetiger (I) and continues through a narrow intramuscular canal near the base of the caruncle but never reaching it or running through it ( Figure 4A View Figure 4 ).
The dorsal lips (dl) of M. carunculata sp. nov., M. lobiferum and M. pigmentum have radiolar appendages (ra) forming two lateral lamellae; they are as long as onequarter the length of the branchial crown ( Figure 2E View Figure 2 ). The lips of M. lobiferum are vascularized by a plexus of small blood vessels (bv), running along the lip as a strongly branched network; each blood vessel is surrounded by coelomic chambers ( Figures 5A,E View Figure 5 ). These blood vessels are derived from the basal branchial blood vessels, which further back communicate with the dorsal vessel. The dorsal lips have hyaline cartilage, but there is no extension of the branchial skeleton. The hyaline cartilage is composed of abundant collagen fibres forming a net with dispersed chondrocytes. A columnar epithelium covers the dorsal lips and it is covered by cuticle ( Figure 5E View Figure 5 ).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Megalomma carunculata
| Marı, Tovar-Herna, a Ana, ndez & Salazar-Vallejo, Sergio I. 2008 |
Megalomma carunculata
| Marı & Tovar-Herna & ndez & Salazar-Vallejo 2008 |
Megalomma carunculata
| Marı & Tovar-Herna & ndez & Salazar-Vallejo 2008 |
Megalomma carunculata
| Marı & Tovar-Herna & ndez & Salazar-Vallejo 2008 |
Megalomma carunculata
| Marı & Tovar-Herna & ndez & Salazar-Vallejo 2008 |
M. pigmentum
| Reish 1963 |
M. pigmentum
| Reish 1963 |
Megalomma
| Johansson 1925 |