
Archamiini, Fraser & Mabuchi, 2014Cheilodipterini Bleeker 1856b Glossamiini, Fraser & Mabuchi, 2014Gymnapogonini Whitley 1941 Lepidamiini, Fraser & Mabuchi, 2014Ostorhinchini Whitley 1959 Pristiapogonini, Fraser & Mabuchi, 2014Rhabdamiini, Fraser & Mabuchi, 2014Siphamiini Smith 1955 Sphaeramiini, Fraser & Mabuchi, 2014Veruluxini, Fraser & Mabuchi, 2014Zoramia Jordan 1917
|
publication ID |
https://dx.doi.org/10.11646/zootaxa.3846.2.1 |
|
publication LSID |
lsid:zoobank.org:pub:3844E8F1-A20C-44B4-9B47-B170F5A7C0C2 |
|
persistent identifier |
https://treatment.plazi.org/id/CA3F4E7D-8104-0B0E-FF78-C3E3FD35D41B |
|
treatment provided by |
Felipe |
|
scientific name |
Archamiini Cheilodipterini Bleeker 1856b Glossamiini Gymnapogonini Whitley 1941 Lepidamiini Ostorhinchini Whitley 1959 Pristiapogonini Rhabdamiini Siphamiini Smith 1955 Sphaeramiini Veruluxini Zoramia Jordan 1917 |
| status |
|
Tribe Archamiini new name Fraser & Mabuchi
Type genus Archamia Gill 1863
Diagnosis. Members of the Apogoninae : VI+I,9 or VII+I,9; anal fin II,12–19; head and body with ctenoid scales; pored lateral-line scales 24–25; preopercle ridge smooth, edges serrate; three supraneurals; supramaxilla absent; basisphenoid present; one pair of uroneurals present or absent; three epurals; five free hypurals, 1–2 fused and 3–4 fused to terminal centrum; free parhypural; caudal fin forked; body translucent without bars and head tinged greenish yellow and small dark basicaudal spot or with dark or yellowish to reddish bars on body, dark basicaudal spot, small or large, compact or diffuse or head and body with one or two narrow yellowish to dark stripes.
Other characteristics. one or two supernumerary dorsal spines; branched first segmented dorsal and anal ray; ctenoid scales on predorsal, cheek, breast, two pelvic scales, and body; ctenoid scale on opercle and onto base of caudal fin; pored lateral-line scales with multiple pores; pectoral fin-rays 11–16; three supraneurals; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; unbranched procurrent rays, longest segmented; teeth on premaxilla, dentary, vomer, palatine, all villiform (one species present on ectopterygoid) or absent on palatine; six infraorbitals, bony shelf on third infraorbital; anterior ceratohyal smooth or notched; developed gill rakers 15–23; 10+14 vertebrae; 8 ribs; 8 epineurals; stomach and intestine blackish, peritoneum silvery with melanophores; low crest on PU2.
Distribution. Archamia and Taeniamia are widespread throughout the Indo-Pacific from the Red Sea, East Africa to Japan and Samoa.
Remarks. This tribe contains two genera, Archamia and Taeniamia , corresponding to the clade XI in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). All the members of the clade were formerly classified under Archamia , but Fraser (2013b) redescribed Archamia as monotypic and recognized a new genus, Taeniamia for the remaining species. The history of this species is given by Gon & Randall (2003). Our molecular results did not disagree with the idea of recognizing two species ( Taeniamia kagoshimanus Döderlein in Steindachner & Döderlein 1883 and T. sansibaricus Pfeffer 1893 ) that has been long confused with Taeniamia fucata ( Cantor 1849) ( Fraser 2013b) . This idea is supported also by the geographic variation in gill raker counts reported by Gon & Randall (2003). Prokofiev (2006) indicated a possible close relationship between the species of " Archamia " ( Archamia + Taeniamia ) and Kurtus gulliveri based on morphological characters. But their monophyly was significantly rejected by the AU test based on the present molecular data (H09; Table 6).
Archami- is the stem for this new tribe.
Tribe Cheilodipterini Bleeker 1856b
Type genus Cheilodipterus Lacepède 1801
Diagnosis. Members of the Apogoninae : VI+I,9; anal fin II,8; head and body with ctenoid scales; pored lateral-line scales 24–26; canine teeth on premaxilla and dentary; preopercle ridge smooth, edges serrate; three supraneurals; reduced supramaxilla; one pair of uroneurals; three epurals; five free hypurals; free parhypural; caudal fin forked or emarginate; dark stripes on head and body, no stripes in second dorsal or anal fin, basicaudal dark spot, band or no spot.
Other characteristics. one supernumerary dorsal spine; branched first segmented dorsal and anal ray; ctenoid scales on predorsal, cheek, breast, two pelvic scales, and body; ctenoid scale on opercle and onto base of caudal fin; pored lateral-line scales with multiple pores; pectoral fin-rays 11–14; three supraneurals; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; unbranched procurrent rays, longest segmented; villiform teeth on vomer and palatine; six infraorbitals, bony shelf on third infraorbital; basisphenoid present; anterior ceratohyal smooth or notched; 10+14 vertebrae; 8 ribs; 7–8 epineurals; stomach and intestine pale with pale peritoneum; low crest on PU2.
Distribution. Cheilodipterus is widespread throughout the Indo-Pacific from the Red Sea, East Africa to Japan and French Polynesia following Gon (1993). There have been several short-lived efforts to subdivide this genus with several names proposed ( Cheilodipterops Schultz 1940 ; Desmoamia Fowler & Bean 1930 ; Paramia Bleeker 1863 ). More molecular analysis among the 17 species should provide insight to relationships not apparent in the five species we sampled.
Remarks. This tribe contains one genus, Cheilodipterus , corresponding to the clade X in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). Canine or caninoid teeth on the premaxilla and dentary, a reduced supramaxilla, a single supernumerary dorsal spine, stripes on body and a diffuse or solid basicaudal spot in most species often with some surrounding yellow are characteristics of this tribe. Cheilodipteroidei, as a family, was recognized by Bleeker (1856b). Schultz (1940) recognized the Cheilodipteridae , but as a broad grouping of a number of families. Smith (1961) and Norman (1966) recognized this group as a subfamily Cheilodipterinae including Coranthus Smith 1961 (now Amioides ) and Paramia Bleeker 1863 (now Cheilodipterus ). Monophyly of Amioides and Cheilodipterus was significantly rejected by AU test (H13; Table 6). The shape and position of the reduced supramaxilla (see Fraser 1972) and canine or caninoid teeth are synapomorphies.
Tribe Glossamiini new name Fraser & Mabuchi
Type genus Glossamia Gill 1863
Diagnosis. Members of the Apogoninae : dorsal fin VI+I,8–10; anal fin II,8–10; pored lateral-line scales 24–50; preopercle ridge smooth edges smooth to serrate; three supraneurals; large supramaxilla present; basisphenoid present, reduced or absent; anterior pair of uroneurals; three epurals; five free hypurals; parhypural separate; two autogenous haemal spines; one supernumerary dorsal spine; caudal fin emarginate, truncate or rounded.
Other characteristics. first dorsal ray unbranched and first anal ray branched and segmented; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; unbranched procurrent rays, longest segmented; head and body with ctenoid scales or cycloid on nape, cheek, opercle, anterior pored lateral-line scales, ctenoid on breast, grading from cycloid to ctenoid posteriorly; pored lateral-line scales simple with one pore on upper side and one on lower side; pectoral fin-rays 13–16; developed gill rakers 7–15; 10+14 vertebrae; 8 ribs; 9 epineurals; teeth in multiple rows on premaxilla, dentary, vomer, palatine, all villiform, present or absent on ectopterygoid; six infraorbitals, bony shelf on third infraorbital; stomach and intestine pale with pale peritoneum; low crest on PU2.
Distribution. Species of Glossamia are found in tidal and flowing freshwaters of Australia and the island of New Guinea. One species is known from Australia (perhaps a species complex) and the rest on the island of New Guinea. The only described species of Yarica is found in tidal streams, flowing freshwater and lowland lakes from Myanmar, Andaman and Nicobar Islands, Thailand, Malaysia, Indonesia, Philippines, Australia, New Guinea, Solomon Islands to New Caledonia and out to Saipan.
Remarks. In the present molecular analyses, Glossamia did not form a robust monophyletic group with any other apogonines (see Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 and Table 5). For this genus, we gave tribe status. We included Yarica ( Fig. 7A View FIGURE 7 ) in this tribe based solely on morphological data. The present molecular analyses reproduced a relatively robust sister relationship between Yarica and Rhabdamia (clade IX). Species of Rhabdamia have a smaller mouth and slender translucent body with a forked caudal fin ( Fig. 7B View FIGURE 7 ) consistent with their marine reef pelagic habitat preferences and differ in their osteological characteristics as follows: 1 or 2 supraneurals; fused hypurals plate consisting of hypurals 1+2+3+4 to the terminal centrum; 1–2 rows of villiform teeth on premaxilla or some canine, 1 row of villiform teeth on dentary, vomer and palatine; 7 epineurals on ribs 1–7; shelf on third infraorbital reduced or absent; uroneurals reduced or absent; supramaxilla absent. The cephalic pores are much more complex for Rhabdamia and free neuromasts much less numerous than for Yarica ( Bergman 2004, Figs. 11, 29–30). Support for combining Glossamia and Yarica are similar body shapes, freshwater habitat preferences, a large supramaxilla of similar shape, a single supernumerary dorsal spine and 9 epineurals. In spite of non-monophyly in the obtained trees, AU test did not reject the monophyly of Glossamia and Yarica (H06, see Table 6). The wide spread distribution of Yarica indicates that this species has euryhaline characteristics imbedded in its life history.
Tribe Gymnapogonini Whitley 1941
Type genus Gymnapogon Regan 1905
Diagnosis. Members of the Apogoninae : VI+I,8–13; anal fin II,8–16; head and body naked or with large weakly ctenoid or cycloid scales; lateral-line scales 23–24 or lateral line a series of free neuromasts to base of caudal fin, interrupted mid-line lateral line present or absent; preopercle ridge smooth, edge with one or more spines at angle; 0–2 ossified supraneurals; supramaxilla, paired uroneurals and basisphenoid absent; 2–3 epurals; hypurals 1–2 fused, 3–4 separate or fused to urostyle; parhypural separate or fused to hypurals 1–2; caudal fin forked or rounded; pterosphenoid and parasphenoid joined excluding prootic from internal orbit rim.
Other characteristics. one supernumerary dorsal-fin spine; unbranched or branched first segmented dorsal-fin ray, branched first anal-fin ray; cycloid or weakly ctenoid scales on predorsal, cheek, breast, two pelvic scales, and body; cycloid scale on opercle and onto base of caudal fin; pectoral-fin rays 10–14; 9+8 segmented principal caudal rays, 13–15 branched, upper one or two and lower one or two unbranched; unbranched procurrent rays, longest segmented; some enlarged teeth on premaxilla and dentary, canine or villiform on vomer, villiform on palatine; six infraorbitals, bony shelf on third infraorbital present or absent; 1–2 postcleithra; anterior ceratohyal smooth or notched; 10+14 or 9+15 ( Cercamia ) vertebrae; 7–8 ribs; 0–6 epipleurals; low crest on PU2.
Distribution. Pseudamiops East Africa to Japan, Hawaii and French Polynesia, Gymnapogon widespread from the Red Sea, East Africa to Japan and French Polynesia, Lachneratus from East Africa to Hawaii and Tonga and Cercamia from the Eastern Indian Ocean to Japan and French Polynesia.
Remarks. This tribe contains four genera, Cercamia , Gymnapogon , Lachneratus and Pseudamiops , corresponding to the clade I in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). Species of the included genera are all translucent with many reductive morphologic characters. Although Pseudamiops was absent from the present analyses, we tentatively associated this genus with the Gymnapogonini based on its translucent body and the reductive morphological characters. Bergman (2004) noted: …"The cephalic lateralis system of Gymnapogon is very similar to that of Pseudamiops . A notable exception being that the preopercular and mandibular canal portions are confluent in Gymnapogon species. " Pseudamiops , with scales, lacks pored, notched or pitted lateral-line scales. Cercamia and Lachneratus have not previously been associated with Gymnapogon . Both genera have deciduous scales and both lack pored, notched or pitted lateral-line scales. Gymnapogon and Pseudamiops were formerly classified under Pseudamiinae , together with Pseudamia . As mentioned above in the remarks of the subfamily Pseudamiinae , the present molecular data significantly rejected their monophyly (H 14 in Table 6). In our molecular trees, two individuals of Cercamia cladara from two distantly distributed populations ( French Polynesia and Palau) were paraphyletic to Gymnapogon and genetically distant from each other, which may indicate needs of taxonomical revisions of them.
Tanaka (1915) described the new family Henicicthyidae for the new genus and single new species Henicichthys foraminosus . Tanaka's family name has been used only in the original publication. Whitley (1941) created the family Gymnapogonidae with Regan's genus as the type species. He noted that the oldest genus name '...becomes the root for the family name." Whitley's (1941) synonymy of the literature for Henicichthys has the last published use of this genus in 1939. Whitley (1941) recognized that Henicichthys foraminosus and Austalaphia annona Whitley 1936 are synonyms of Gymnapogon . The sole use of Gymnapogon has been continuous since 1941. Fowler (1944) and Lindberg (1971) used Gymnapogonidae . We regarded Gymnapogonini as an easily recognized tribal name among apogonids and is its preferred use.
Tribe Lepidamiini new name Fraser & Mabuchi
Type genus Lepidamia Gill 1863
Diagnosis. Members of the Apogoninae : dorsal fin VII(I)-I,9 or VIII-I,9; anal fin II,8; head and body with ctenoid scales; pored lateral-line scales 35–48; preopercle ridge smooth, edges serrate; three supraneurals; supramaxilla absent; basisphenoid present; one pair of reduced uroneurals present; three epurals; five free hypurals; parhypural separate; two autogenous haemal spines; two supernumerary dorsal spines, caudal fin forked.
Other characteristics. first fin ray in second dorsal fin branched and segmented; first anal-fin ray branched and segmented; ctenoid scales on predorsal, cheek, breast, four pelvic scales, and body; ctenoid scales on opercle and onto base of caudal fin; pored lateral-line scales with multiple pores; 15 branched, upper and lower unbranched; developed gill rakers 8–17; unbranched procurrent rays, longest segmented; 10+14 vertebrae; 8 ribs; 9 epipleurals; teeth in multiple rows on premaxilla, dentary, vomer, palatine, all villiform; six infraorbitals, bony shelf on third infraorbital; stomach and intestine pale, peritoneum pale; low crest on PU2.
Distribution. Lepidamia with four species has been collected only from continental marine waters from South Africa to China ( Gon 1995). Members of this tribe have not been collected in Australia or New Guinea. One specimen (USNM 175754, 96 mm SL) was collected in 1909 from Manila Bay, Luzon, Philippines. None have been reported elsewhere in the Philippines or again from Manila Bay. Perhaps the location is erroneous.
Remarks. We did not have tissue from any species. Based on known morphological characters including color patterns of the juveniles and adults, body shape and small body scales, we recognized Lepidamia in its own tribe for the present.
Tribe Ostorhinchini Whitley 1959
Type genus Ostorhinchus Lacepède 1802
Diagnosis. Members of the Apogoninae : dorsal fin VI+I,9 or VII+I,9; anal fin II,8–9; head and body with ctenoid scales; pored lateral-line scales 6–26; preopercle ridge smooth, edges serrate; 2–3 supraneurals; supramaxilla absent; basisphenoid present; one pair of reduced uroneurals present or absent; three epurals; five free hypurals or 1–2 fused; parhypural separate; caudal fin forked; head and/or body with one or more light or dark stripes, bars rarely present, basicaudal spot or broad mark present or absent.
Other characteristics. one or two supernumerary dorsal spines; branched first segmented fin ray in second dorsal-fin; first anal-fin ray branched and segmented; ctenoid scales on predorsal, cheek, breast, two pelvic scales, and body; ctenoid scale on opercle and onto base of caudal fin; pored lateral-line scales with multiple pores; pectoral fin-rays 11–16; three supraneurals; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; unbranched procurrent rays, longest segmented; teeth in one or multiple rows on premaxilla, dentary, vomer, palatine, all villiform or absent on palatine, sometimes enlarged, but not canine on side of dentary; six infraorbitals, bony shelf on third infraorbital; anterior ceratohyal smooth or notched; developed gill rakers 10–26; 10+14 vertebrae; 8 ribs; 8 epipleurals; blackish stomachs and intestines; low crest on PU2.
Distribution. The restricted Ostorhinchus is widespread throughout the Indo-Pacific from the Red Sea, East Africa to Japan, Hawaii, Easter Island and French Polynesia.
Remarks. Monophyly of recent Ostorhinchus (species indicated by blue in Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 ) was significantly rejected by AU test (H 16 in Table 6). This tribe, however, corresponds to one of the five lineages of the recent Ostorhinchus , clade XII. It included most species of the recent Ostorhinchus : many striped species of Ostorhinchus (species of Ostorhinchus -5) and Brephamia parvula Smith & Radcliffe in Radcliffe 1912 ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). Other species of the recent Ostorhinchus were divided into four lineages, Ostorhinchus -1, 2, 3 and 4. They were all placed far from the clade XII. Except for Ostorhinchus -3 ( O. margaritophorus ), monophyly between this tribe (clade XII) and each of the remaining lineages was rejected by AU tests (H07, 08 and 11 in Table 6). Although the bootstrap values supporting the clade XII were not so high (<50%) both in the ML ( Fig. 2 View FIGURE 2 ) and MP ( Figs. 4 View FIGURE 4 and 5 View FIGURE 5 ) trees ( Table 4), and further the clade was divided into two separate clades in the BA tree ( Fig. 3 View FIGURE 3 ), we are treating this group as a single genus, based on the typical color pattern (stripes on body and/or head) shared by most of the included species. This group corresponds to the "striped group" of Ostorhinchus ( Ostorhinchus III ) in Mabuchi et al. (2006). One of the other recent Ostorhinchus lineages, O. margaritophorus , has long stripes, short bars between two long stripes on body and fused hypurals 1+2. Phylogenetic position of this species remains unsettled in the molecular trees ( Table 5), but monophyly between this species and the members of this tribe was not rejected by AU test (H01 in Table 6). Based on its featured color pattern, we tentatively included this species in this tribe. Apogon ( Brephamia) parvula has a very similar species within this tribe. It is Ostorhinchus neotes . Although they were not sister to each other in the molecular trees, their monophyly was not rejected by AU test (H 12 in Table 6). While recognition of Brephamia Jordan in Jordan & Jordan (1922) as a subgenus or genus needs further evaluation, we synonymized it under the restricted Ostorhinchus for the present. Likely synapomorphies may include color pattern groupings, probably at the subgenus level. At the genus level, with the exception of the loss of tiny first dorsal-fin spines, all species have 7 visible first dorsal spines, a serrated preopercular edge and most have blackish stomachs and intestines.
There are two available names ( Gronovichthys Whitley 1929 ; Lovamia Whitley 1930 ) which could be used in the future. There are at least 93 species in the group. We have sampled 31 species. Whitley (1959) first used the present name at family level Ostorhinchidae. The name appeared once. We use the name at the tribal level.
Tribe Pristiapogonini new name Fraser & Mabuchi
Type genus Pristiapogon Klunzinger 1870
Diagnosis. Members of the Apogoninae : dorsal fin VI+I,9 or VII+I,9; anal fin II,8; head and body with ctenoid scales; pored lateral-line scales 23–25; preopercle ridge serrate, edges serrate; infraorbitals serrate; three supraneurals; supramaxilla absent; basisphenoid present; one pair of uroneurals or absent; three epurals; five free hypurals; parhypural separate; one or two supernumerary dorsal spines; caudal fin forked.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal-fin ray branched and segmented; ctenoid scales on predorsal, cheek, breast, two pelvic scales, and body; ctenoid scales on opercle and onto base of caudal fin; pored lateral-line scales with multiple pores; pectoral fin-rays 12–16; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; unbranched procurrent rays, longest segmented; teeth in one or multiple rows on premaxilla, dentary, vomer, palatine, all villiform or absent on palatine; six infraorbitals, bony shelf on third infraorbital; anterior ceratohyal smooth or notched; developed gill rakers 10–19; 10+14 vertebrae; 8 ribs; 8 epipleurals; low crest on PU2.
Distribution. Pristiapogon is wide spread from East Africa, Red Sea, Indian Ocean Islands, Indonesia, Philippines, Pacific islands to Hawaii, Japan, French Polynesia and Australia. One species of Pristicon Fraser 1972 is restricted to the West Pacific. Another species reaches out to some islands on the Pacific Plate. One species of Pristicon has been reported from the west coast of India, with a gap between India and Indonesia ( Suresh & Thomas 2007).
Remarks. This tribe contains two genera, Pristiapogon and Pristicon , corresponding to the clade VI in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). Although bootstrap values supporting the tribe were not so high (62 and 56% BPs in ML and MP analyses, respectively), this tribe is morphologically well-defined: relatively large body with serrations on the preopercle ridge (a likely synapomorphy), edges and infraorbitals (a likely synapomorphy). Species of Pristiapogon usually have a darkish single stripe and/or a variable basicaudal spot while Pristicon have bars or saddles under the dorsal fins and spots at the base of the caudal fin or on the opercle.
Tribe Rhabdamiini new name Fraser & Mabuchi
Type genus Rhabdamia Weber 1909
Diagnosis. Members of the Apogoninae : dorsal fin VI+I,9 or VII+I,10–11; anal fin II,11–13; head and body with weakly ctenoid or cycloid; pored lateral-line scales 23–24; preopercle ridge smooth, edges smooth; two supraneurals; supramaxilla absent; basisphenoid present or absent; anterior pair of uroneurals reduced or absent; three epurals; hypurals 1+2+3+4 fused, the plate fused to terminal centrum; parhypural separate; no autogenous haemal spines; 1–2 supernumerary dorsal spines; caudal fin forked.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal-fin ray branched and segmented; pectoral-fin rays 13–17; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; developed gill rakers 17–31; unbranched procurrent rays, longest segmented; villiform teeth in one or two rows on premaxilla, or with a few canines ( Bentuviaichthys ), one villiform row on dentary and vomer, one villiform row or absent on palatine; six infraorbitals, bony shelf reduce or absent on third infraorbital; stomach and intestine pale with melanophores and silvery peritoneum with melanophores; 10+14 vertebrae; low crest on PU2.
Distribution. No reviews of the species have been made, but members have been reported from East Africa, the Red Sea, islands in the Indian Ocean, the West Pacific and out onto the Pacific Plate.
Remarks. This tribe contains one genus Rhabdamia (including two subgenera Rhabdamia and Bentuviaichthys sensu Fraser 1972 ). There are six names (Appendix A). Three species are streamlined apogonines exploiting the near pelagic zone of coral reefs for food. Likely synapomorphies include smooth preopercle ridge and edges, reduced dentition, fused hypurals, and two supraneurals. All species are mostly translucent. In the present molecular analyses, they formed a robust monophyletic group (clade IX) with Yarica hyalosoma ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4) (see also Fig. 7 View FIGURE 7 ), but it is very unlikely based on morphological evidence (for details, see the remarks of the tribe Glossamiini ). In Fraser (1972), subgenus Rhabdamia was classified under the genus Rhabdamia together with the subgenera Bentuviaichthys and Verulux Fraser 1972 . The last lineage, Verulux (clade V), was placed far from the first lineage, Rhabdamia , in the present molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 ). Although monophyly between the two lineages was not rejected by AU test (H05 in Table 6), we recognized each of them in its own tribe for the present (for Verulux , see the tribe Veruluxini ). The present molecular analyses did not include Bentuviaichthys , but it was included in this tribe based on the following morphological characters: fused hypurals 1+2+3+4, epipleurals absent on last three ribs, single row of teeth on dentary, two supraneurals, slender, mostly translucent body and forked caudal fin. Two individuals of Rhabdamia gracilis from two distantly distributed populations ( Fiji and Ryukyu Islands) were paraphyletic to R. spilota Allen & Kuiter 1994 and genetically distant from each other, which may indicate needs of taxonomical revisions of them.
Tribe Siphamiini Smith 1955
Type genus Siphamia Weber 1909
Diagnosis. Members of the Apogoninae : dorsal fin VI–VII + I,7–11; anal fin II,7–11; pored lateral-line scales 0–24; preopercular ridge smooth, edges smooth to serrate; supraneurals 1–2; supramaxilla absent; basisphenoid absent; uroneurals absent; two epurals; hypurals 1+2 and 3+4 fused into two plates, upper plate fused to terminal centrum; parhypural separate; one or two supernumerary dorsal spines; caudal fin emarginate or forked; bacteria bioluminescent system from hyal region to along body above or past anal-fin base.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal-fin ray branched and segmented; ctenoid, cycloid or spinoid scales; median predorsal scales 0–6; pectoral rays 11–16; segmented principal caudal rays 9+8, 15 branched, upper and lower unbranched; unbranched procurrent rays, longest segmented; villiform teeth in one or multiple rows on premaxilla, dentary, vomer, palatine, or absent on palatine; developed gill rakers 6–18; lateral-line scales usually with a vertical row of free neuromasts; six infraorbitals, bony shelf on third infraorbital; anterior ceratohyal smooth or notched; developed gill rakers 10–26; 10+14 vertebrae; 8 ribs; 8 epipleurals; stomach, intestine and peritoneum generally pale with variously sized melanophores; low crest on PU2.
Distribution. Representatives of the tribe are found from East Africa, Red Sea, islands in the Indian Ocean, throughout the West Pacific to Japan, onto the Pacific Plate to French Polynesia and Australia ( Gon & Allen 2012).
Remarks. This tribe contains one genus Siphamia , corresponding to the clade VII in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). All species of Siphamia have bioluminescent bacteria in a specialized organ (a synapomorphy for the species) unique among apogonines. Smith (1955) proposed placing species of Siphamia in its own subfamily Siphamiinae . Our results based on analysis of five of the twenty-three species suggest that Siphamia roseigaster Ramsay & Ogilby 1887 could be recognized in its own genus Adenapogon McCulloch 1921 , because it was placed relatively far from the remaining species. Gon & Allen's (2012) results based on morphology suggest that two other Australian species belong in Adenapogon and that Fodifoa Whitley 1936 is available for another group of species. We defer to Ofer Gon who is continuing to work on relationships within this tribe ( Gon & Allen 2012).
Tribe Sphaeramiini new name Fraser & Mabuchi
Type genus Sphaeramia Fowler & Bean 1930
Diagnosis. Members of the Apogoninae : dorsal fin VII+I,9–10, VII(I)+I,9–14 or VIII+I,9–10 with eighth spine reduced to a tiny visible spine or a nubbin hidden under skin supported by a free sixth distal radial; anal fin II,8–13; head and body with ctenoid scales; pored lateral-line scales 23–27; preopercle ridge smooth, edges serrate; 2–3 supraneurals; supramaxilla absent; basisphenoid present; one pair of uroneurals; three epurals; five free hypurals or 1+2 fused and 3+4 fused with 3+4 fused to terminal centrum; parhypural separate; two autogenous haemal spines; two supernumerary dorsal spines; caudal fin forked, emarginate or truncate.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal-fin ray branched and segmented; ctenoid scales on predorsal, cheek, breast, two pelvic scales, and body; ctenoid scales on opercle and onto base of caudal fin; 1–3 predorsal scales cycloid or ctenoid; pored lateral-line scales with multiple pores; pectoral-fin rays 13–17; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; developed gill rakers 7–38; unbranched procurrent rays, longest segmented or spinous; third or fourth dorsal spine longest; 10+14 or 10+15 vertebrae; 8 ribs; 8–9 epipleurals; teeth in one or multiple rows on premaxilla, dentary, vomer, palatine, all villiform; six infraorbitals, bony shelf on third infraorbital; stomach and intestine pale or black with pale peritoneum; low crest on PU2.
Distribution. Species of Apogonichthyoides are found throughout the Indo-West Pacific from East Africa, Red Sea, India, some island of the Indian Ocean, throughout Indonesia, Philippines to Japan, New Guinea, Australia and the Solomon Islands. Some species have restricted distribution and some are rare in collections. Species of Jaydia Smith 1961 have a more continental distribution, mostly caught in trawls, and are widespread from Africa to Japan, New Guinea, Australia and larger islands in the Coral Sea. Representatives of Nectamia are found on reef habitats from East Africa, Red Sea, islands in the Indian Ocean, throughout the West Pacific to Japan, onto the Pacific Plate out to the Line Islands and French Polynesia. Sphaeramia , represented by two species, one found in mangrove habitats and one on coral reefs, also have wide distribution in the Indo-West Pacific. Quinca , represented by a single species, is known from northwestern coast of Australia.
Remarks. This tribe contains six genera, Apogonichthyoides , Jaydia , Nectamia , Pterapogon Koumans 1933 , Quinca and Sphaeramia , almost corresponding to the clade IV in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). First three of the six genera were formerly classified under Ostorhinchus , corresponding to "barred group" of Ostorhinchus ( Ostorhinchus II ) in Mabuchi et al. (2006). Two of the remaining three genera ( Pterapogon and Sphaeramia ) have distinct bar(s) on body. In addition to the six genera, corresponding molecular clade included three species of Ostorhinchus -2 ( O. hoevenii Bleeker 1854b , O. ishigakiensis Ida & Moyer 1974 , and O. rueppellii Günther 1859 ). But there are no morphological characters that support this association. We defer taxonomical action on this possible relationship for the present. This clade further included another species of the former Ostorhinchus ( Ostorhinchus -1). For this species, the latest authors ( Allen & Erdmann 2012) provisionally used Apogonichthyoides as its genus name, and we followed it here, but such application of the name made this genus paraphyletic in our molecular trees. Based on its morphological features, we tentatively included it [ Apogonichthyoides (?) melas ] in this tribe, but further study including more Sphaeramiini species will be needed to clarify its taxonomic status. In the present molecular analyses, phylogenetic position of Pterapogon remained unsettled ( Table 5). It was sister to the clade VII ( Siphamiini ) in ML tree ( Fig. 2 View FIGURE 2 ), sister to Ostorhinchus margaritophorus in MP tree-A ( Fig. 4 View FIGURE 4 ), and nested within the clade XII ( Ostorhinchini ) in the MP tree-B ( Fig. 5 View FIGURE 5 ). It formed a polytomy with O. margaritophorus , clade VII, clade VIII, and a large clade including clades IX–XII in BA tree ( Fig. 3 View FIGURE 3 ). Pterapogon has, however, a synapomorphy (spinous procurrent caudal rays) uniquely held with Sphaeramia as well as similarities in color patterns and body shape, but differs with fused hypurals 1+2 and 3+4. Based on these morphological features, we tentatively included this species in the tribe Sphaeramiini . Monophyly of them was not rejected by AU test (H02 in Table 6), although its inclusion within Ostorhinchini was also not rejected (H03 in Table 6). We had no good DNA extracts from tissue of Quinca . This monotypic genus has morphological characteristics and color pattern suggestive of a relationship with Apogonichthyoides , and was provisionally place in the Sphaeramiini .
Tribe Veruluxini new name Fraser & Mabuchi
Type genus Verulux Fraser 1972
Diagnosis. Members of the Apogoninae : dorsal fin VI–I,9; anal fin II,9; head and body with weakly ctenoid or cycloid scales; pored lateral-line scales 24; preopercle ridge smooth, edges smooth; one supraneural; supramaxilla absent; basisphenoid present; uroneurals absent; two epurals; fused hypurals 1+2+3+4, fused to terminal centrum; no autogenous haemal spines; one supernumerary dorsal spine; bioluminescent organ under cleithrum; caudal fin forked.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal fin-ray branched and segmented; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; developed gill rakers 12–15; unbranched procurrent rays, longest segmented; pectoral fin-rays 14–16; 10+14 vertebrae; 8 ribs; 5–6 epipleurals; teeth in one row on premaxilla, dentary, vomer, palatine, all villiform, or absent on palatine; six infraorbitals, bony shelf absent on third infraorbital; stomach and intestine black with silvery peritoneum; low crest on PU2.
Distribution. The single described species is known from East Africa, Red Sea, islands in the Indian Ocean, throughout the West Pacific to Japan, onto the Pacific Plate to Marshall Islands, Vanuatu, New Caledonia and Australia.
Remarks. This tribe contains only one species, Verulux cypselurus , corresponding to the clade V in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). Representatives of Verulux (two individuals from Ryukyu Islands and Seychelles) were monophyletic separated by a moderate genetic distance, which may indicate presence of cryptic species. This lineage was recovered far from Rhabdamia (tribe Rhabdamiini ), although their monophyly (genus Rhabdamia sensu Fraser 1972 ) was not rejected by AU test (H05 in Table 6). The single described species has a bioluminescent organ under cleithrum unlike Rhabdamia .
Tribe Zoramiini new name Fraser & Mabuchi
Type genus Zoramia Jordan 1917
Diagnosis. Members of the Apogoninae : dorsal fin VI–I,9; anal fin II,8–9; head and body with ctenoid scales; pored lateral-line scales 24; preopercle ridge smooth, edges serrate; three supraneurals; supramaxilla absent; basisphenoid present; anterior pair of uroneurals; three epurals; five free hypurals; parhypural separate; two autogenous haemal spines; one supernumerary dorsal spine; caudal fin forked.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal-fin ray branched and segmented; second dorsal spine longest, very long and filamentous in some species of both genera; ctenoid scales on predorsal, cheek, breast, two pelvic scales, and body; ctenoid scales on opercle and onto base of caudal fin; cycloid or ctenoid predorsal scales1–3; pored lateral-line scales simple with one pore above and one below midline; pectoral-fin rays 13–17; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; developed gill rakers 7–38; unbranched procurrent rays, longest segmented; 10+14 vertebrae; 8 ribs; 7–8 epipleurals; teeth in one, two or multiple rows on premaxilla, dentary, vomer, palatine, all villiform; six infraorbitals, bony shelf absent or on third infraorbital; stomach and intestine black with silvery peritoneum; low crest on PU2.
Distribution. Species of Zoramia are known from East Africa, Red Sea, islands in the Indian Ocean, throughout the West Pacific to Japan, onto the Pacific Plate to Marshall Islands, Samoa, Tonga, Solomon Islands, New Caledonia and Australia. Species of Fibramia are known from East Africa, some islands in the Indian Ocean, Sri Lanka, throughout the West Pacific to Philippines, Solomon Islands, Santa Cruz Islands, Fiji, Vanuatu, New Caledonia, Australia, Samoa and Tonga.
Remarks. This tribe contains two genera, a new genus Fibramia described below and Zoramia , corresponding to the clade VIII in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). Both genera have species with elongate second dorsal spine, a single supernumerary dorsal spine, and can be found in tidal fresh and brackish water, near shore waters and in lagoons. Fibramia corresponds to the clade of Ostorhinchus - 4 in the present study, and that of Ostorhinchus I in Mabuchi et al. (2006).
Tribe Cheilodipterini Bleeker 1856b
Type genus Cheilodipterus Lacepède 1801
Diagnosis. Members of the Apogoninae : VI+I,9; anal fin II,8; head and body with ctenoid scales; pored lateral-line scales 24–26; canine teeth on premaxilla and dentary; preopercle ridge smooth, edges serrate; three supraneurals; reduced supramaxilla; one pair of uroneurals; three epurals; five free hypurals; free parhypural; caudal fin forked or emarginate; dark stripes on head and body, no stripes in second dorsal or anal fin, basicaudal dark spot, band or no spot.
Other characteristics. one supernumerary dorsal spine; branched first segmented dorsal and anal ray; ctenoid scales on predorsal, cheek, breast, two pelvic scales, and body; ctenoid scale on opercle and onto base of caudal fin; pored lateral-line scales with multiple pores; pectoral fin-rays 11–14; three supraneurals; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; unbranched procurrent rays, longest segmented; villiform teeth on vomer and palatine; six infraorbitals, bony shelf on third infraorbital; basisphenoid present; anterior ceratohyal smooth or notched; 10+14 vertebrae; 8 ribs; 7–8 epineurals; stomach and intestine pale with pale peritoneum; low crest on PU2.
Distribution. Cheilodipterus is widespread throughout the Indo-Pacific from the Red Sea, East Africa to Japan and French Polynesia following Gon (1993). There have been several short-lived efforts to subdivide this genus with several names proposed ( Cheilodipterops Schultz 1940 ; Desmoamia Fowler & Bean 1930 ; Paramia Bleeker 1863 ). More molecular analysis among the 17 species should provide insight to relationships not apparent in the five species we sampled.
Remarks. This tribe contains one genus, Cheilodipterus , corresponding to the clade X in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). Canine or caninoid teeth on the premaxilla and dentary, a reduced supramaxilla, a single supernumerary dorsal spine, stripes on body and a diffuse or solid basicaudal spot in most species often with some surrounding yellow are characteristics of this tribe. Cheilodipteroidei, as a family, was recognized by Bleeker (1856b). Schultz (1940) recognized the Cheilodipteridae , but as a broad grouping of a number of families. Smith (1961) and Norman (1966) recognized this group as a subfamily Cheilodipterinae including Coranthus Smith 1961 (now Amioides ) and Paramia Bleeker 1863 (now Cheilodipterus ). Monophyly of Amioides and Cheilodipterus was significantly rejected by AU test (H13; Table 6). The shape and position of the reduced supramaxilla (see Fraser 1972) and canine or caninoid teeth are synapomorphies.
Tribe Glossamiini new name Fraser & Mabuchi
Type genus Glossamia Gill 1863
Diagnosis. Members of the Apogoninae : dorsal fin VI+I,8–10; anal fin II,8–10; pored lateral-line scales 24–50; preopercle ridge smooth edges smooth to serrate; three supraneurals; large supramaxilla present; basisphenoid present, reduced or absent; anterior pair of uroneurals; three epurals; five free hypurals; parhypural separate; two autogenous haemal spines; one supernumerary dorsal spine; caudal fin emarginate, truncate or rounded.
Other characteristics. first dorsal ray unbranched and first anal ray branched and segmented; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; unbranched procurrent rays, longest segmented; head and body with ctenoid scales or cycloid on nape, cheek, opercle, anterior pored lateral-line scales, ctenoid on breast, grading from cycloid to ctenoid posteriorly; pored lateral-line scales simple with one pore on upper side and one on lower side; pectoral fin-rays 13–16; developed gill rakers 7–15; 10+14 vertebrae; 8 ribs; 9 epineurals; teeth in multiple rows on premaxilla, dentary, vomer, palatine, all villiform, present or absent on ectopterygoid; six infraorbitals, bony shelf on third infraorbital; stomach and intestine pale with pale peritoneum; low crest on PU2.
Distribution. Species of Glossamia are found in tidal and flowing freshwaters of Australia and the island of New Guinea. One species is known from Australia (perhaps a species complex) and the rest on the island of New Guinea. The only described species of Yarica is found in tidal streams, flowing freshwater and lowland lakes from Myanmar, Andaman and Nicobar Islands, Thailand, Malaysia, Indonesia, Philippines, Australia, New Guinea, Solomon Islands to New Caledonia and out to Saipan.
Remarks. In the present molecular analyses, Glossamia did not form a robust monophyletic group with any other apogonines (see Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 and Table 5). For this genus, we gave tribe status. We included Yarica ( Fig. 7A View FIGURE 7 ) in this tribe based solely on morphological data. The present molecular analyses reproduced a relatively robust sister relationship between Yarica and Rhabdamia (clade IX). Species of Rhabdamia have a smaller mouth and slender translucent body with a forked caudal fin ( Fig. 7B View FIGURE 7 ) consistent with their marine reef pelagic habitat preferences and differ in their osteological characteristics as follows: 1 or 2 supraneurals; fused hypurals plate consisting of hypurals 1+2+3+4 to the terminal centrum; 1–2 rows of villiform teeth on premaxilla or some canine, 1 row of villiform teeth on dentary, vomer and palatine; 7 epineurals on ribs 1–7; shelf on third infraorbital reduced or absent; uroneurals reduced or absent; supramaxilla absent. The cephalic pores are much more complex for Rhabdamia and free neuromasts much less numerous than for Yarica ( Bergman 2004, Figs. 11, 29–30). Support for combining Glossamia and Yarica are similar body shapes, freshwater habitat preferences, a large supramaxilla of similar shape, a single supernumerary dorsal spine and 9 epineurals. In spite of non-monophyly in the obtained trees, AU test did not reject the monophyly of Glossamia and Yarica (H06, see Table 6). The wide spread distribution of Yarica indicates that this species has euryhaline characteristics imbedded in its life history.
Tribe Gymnapogonini Whitley 1941
Type genus Gymnapogon Regan 1905
Diagnosis. Members of the Apogoninae : VI+I,8–13; anal fin II,8–16; head and body naked or with large weakly ctenoid or cycloid scales; lateral-line scales 23–24 or lateral line a series of free neuromasts to base of caudal fin, interrupted mid-line lateral line present or absent; preopercle ridge smooth, edge with one or more spines at angle; 0–2 ossified supraneurals; supramaxilla, paired uroneurals and basisphenoid absent; 2–3 epurals; hypurals 1–2 fused, 3–4 separate or fused to urostyle; parhypural separate or fused to hypurals 1–2; caudal fin forked or rounded; pterosphenoid and parasphenoid joined excluding prootic from internal orbit rim.
Other characteristics. one supernumerary dorsal-fin spine; unbranched or branched first segmented dorsal-fin ray, branched first anal-fin ray; cycloid or weakly ctenoid scales on predorsal, cheek, breast, two pelvic scales, and body; cycloid scale on opercle and onto base of caudal fin; pectoral-fin rays 10–14; 9+8 segmented principal caudal rays, 13–15 branched, upper one or two and lower one or two unbranched; unbranched procurrent rays, longest segmented; some enlarged teeth on premaxilla and dentary, canine or villiform on vomer, villiform on palatine; six infraorbitals, bony shelf on third infraorbital present or absent; 1–2 postcleithra; anterior ceratohyal smooth or notched; 10+14 or 9+15 ( Cercamia ) vertebrae; 7–8 ribs; 0–6 epipleurals; low crest on PU2.
Distribution. Pseudamiops East Africa to Japan, Hawaii and French Polynesia, Gymnapogon widespread from the Red Sea, East Africa to Japan and French Polynesia, Lachneratus from East Africa to Hawaii and Tonga and Cercamia from the Eastern Indian Ocean to Japan and French Polynesia.
Remarks. This tribe contains four genera, Cercamia , Gymnapogon , Lachneratus and Pseudamiops , corresponding to the clade I in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). Species of the included genera are all translucent with many reductive morphologic characters. Although Pseudamiops was absent from the present analyses, we tentatively associated this genus with the Gymnapogonini based on its translucent body and the reductive morphological characters. Bergman (2004) noted: …"The cephalic lateralis system of Gymnapogon is very similar to that of Pseudamiops . A notable exception being that the preopercular and mandibular canal portions are confluent in Gymnapogon species. " Pseudamiops , with scales, lacks pored, notched or pitted lateral-line scales. Cercamia and Lachneratus have not previously been associated with Gymnapogon . Both genera have deciduous scales and both lack pored, notched or pitted lateral-line scales. Gymnapogon and Pseudamiops were formerly classified under Pseudamiinae , together with Pseudamia . As mentioned above in the remarks of the subfamily Pseudamiinae , the present molecular data significantly rejected their monophyly (H 14 in Table 6). In our molecular trees, two individuals of Cercamia cladara from two distantly distributed populations ( French Polynesia and Palau) were paraphyletic to Gymnapogon and genetically distant from each other, which may indicate needs of taxonomical revisions of them.
Tanaka (1915) described the new family Henicicthyidae for the new genus and single new species Henicichthys foraminosus . Tanaka's family name has been used only in the original publication. Whitley (1941) created the family Gymnapogonidae with Regan's genus as the type species. He noted that the oldest genus name '...becomes the root for the family name." Whitley's (1941) synonymy of the literature for Henicichthys has the last published use of this genus in 1939. Whitley (1941) recognized that Henicichthys foraminosus and Austalaphia annona Whitley 1936 are synonyms of Gymnapogon . The sole use of Gymnapogon has been continuous since 1941. Fowler (1944) and Lindberg (1971) used Gymnapogonidae . We regarded Gymnapogonini as an easily recognized tribal name among apogonids and is its preferred use.
Tribe Lepidamiini new name Fraser & Mabuchi
Type genus Lepidamia Gill 1863
Diagnosis. Members of the Apogoninae : dorsal fin VII(I)-I,9 or VIII-I,9; anal fin II,8; head and body with ctenoid scales; pored lateral-line scales 35–48; preopercle ridge smooth, edges serrate; three supraneurals; supramaxilla absent; basisphenoid present; one pair of reduced uroneurals present; three epurals; five free hypurals; parhypural separate; two autogenous haemal spines; two supernumerary dorsal spines, caudal fin forked.
Other characteristics. first fin ray in second dorsal fin branched and segmented; first anal-fin ray branched and segmented; ctenoid scales on predorsal, cheek, breast, four pelvic scales, and body; ctenoid scales on opercle and onto base of caudal fin; pored lateral-line scales with multiple pores; 15 branched, upper and lower unbranched; developed gill rakers 8–17; unbranched procurrent rays, longest segmented; 10+14 vertebrae; 8 ribs; 9 epipleurals; teeth in multiple rows on premaxilla, dentary, vomer, palatine, all villiform; six infraorbitals, bony shelf on third infraorbital; stomach and intestine pale, peritoneum pale; low crest on PU2.
Distribution. Lepidamia with four species has been collected only from continental marine waters from South Africa to China ( Gon 1995). Members of this tribe have not been collected in Australia or New Guinea. One specimen (USNM 175754, 96 mm SL) was collected in 1909 from Manila Bay, Luzon, Philippines. None have been reported elsewhere in the Philippines or again from Manila Bay. Perhaps the location is erroneous.
Remarks. We did not have tissue from any species. Based on known morphological characters including color patterns of the juveniles and adults, body shape and small body scales, we recognized Lepidamia in its own tribe for the present.
Tribe Ostorhinchini Whitley 1959
Type genus Ostorhinchus Lacepède 1802
Diagnosis. Members of the Apogoninae : dorsal fin VI+I,9 or VII+I,9; anal fin II,8–9; head and body with ctenoid scales; pored lateral-line scales 6–26; preopercle ridge smooth, edges serrate; 2–3 supraneurals; supramaxilla absent; basisphenoid present; one pair of reduced uroneurals present or absent; three epurals; five free hypurals or 1–2 fused; parhypural separate; caudal fin forked; head and/or body with one or more light or dark stripes, bars rarely present, basicaudal spot or broad mark present or absent.
Other characteristics. one or two supernumerary dorsal spines; branched first segmented fin ray in second dorsal-fin; first anal-fin ray branched and segmented; ctenoid scales on predorsal, cheek, breast, two pelvic scales, and body; ctenoid scale on opercle and onto base of caudal fin; pored lateral-line scales with multiple pores; pectoral fin-rays 11–16; three supraneurals; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; unbranched procurrent rays, longest segmented; teeth in one or multiple rows on premaxilla, dentary, vomer, palatine, all villiform or absent on palatine, sometimes enlarged, but not canine on side of dentary; six infraorbitals, bony shelf on third infraorbital; anterior ceratohyal smooth or notched; developed gill rakers 10–26; 10+14 vertebrae; 8 ribs; 8 epipleurals; blackish stomachs and intestines; low crest on PU2.
Distribution. The restricted Ostorhinchus is widespread throughout the Indo-Pacific from the Red Sea, East Africa to Japan, Hawaii, Easter Island and French Polynesia.
Remarks. Monophyly of recent Ostorhinchus (species indicated by blue in Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 ) was significantly rejected by AU test (H 16 in Table 6). This tribe, however, corresponds to one of the five lineages of the recent Ostorhinchus , clade XII. It included most species of the recent Ostorhinchus : many striped species of Ostorhinchus (species of Ostorhinchus -5) and Brephamia parvula Smith & Radcliffe in Radcliffe 1912 ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). Other species of the recent Ostorhinchus were divided into four lineages, Ostorhinchus -1, 2, 3 and 4. They were all placed far from the clade XII. Except for Ostorhinchus -3 ( O. margaritophorus ), monophyly between this tribe (clade XII) and each of the remaining lineages was rejected by AU tests (H07, 08 and 11 in Table 6). Although the bootstrap values supporting the clade XII were not so high (<50%) both in the ML ( Fig. 2 View FIGURE 2 ) and MP ( Figs. 4 View FIGURE 4 and 5 View FIGURE 5 ) trees ( Table 4), and further the clade was divided into two separate clades in the BA tree ( Fig. 3 View FIGURE 3 ), we are treating this group as a single genus, based on the typical color pattern (stripes on body and/or head) shared by most of the included species. This group corresponds to the "striped group" of Ostorhinchus ( Ostorhinchus III ) in Mabuchi et al. (2006). One of the other recent Ostorhinchus lineages, O. margaritophorus , has long stripes, short bars between two long stripes on body and fused hypurals 1+2. Phylogenetic position of this species remains unsettled in the molecular trees ( Table 5), but monophyly between this species and the members of this tribe was not rejected by AU test (H01 in Table 6). Based on its featured color pattern, we tentatively included this species in this tribe. Apogon ( Brephamia) parvula has a very similar species within this tribe. It is Ostorhinchus neotes . Although they were not sister to each other in the molecular trees, their monophyly was not rejected by AU test (H 12 in Table 6). While recognition of Brephamia Jordan in Jordan & Jordan (1922) as a subgenus or genus needs further evaluation, we synonymized it under the restricted Ostorhinchus for the present. Likely synapomorphies may include color pattern groupings, probably at the subgenus level. At the genus level, with the exception of the loss of tiny first dorsal-fin spines, all species have 7 visible first dorsal spines, a serrated preopercular edge and most have blackish stomachs and intestines.
There are two available names ( Gronovichthys Whitley 1929 ; Lovamia Whitley 1930 ) which could be used in the future. There are at least 93 species in the group. We have sampled 31 species. Whitley (1959) first used the present name at family level Ostorhinchidae. The name appeared once. We use the name at the tribal level.
Tribe Pristiapogonini new name Fraser & Mabuchi
Type genus Pristiapogon Klunzinger 1870
Diagnosis. Members of the Apogoninae : dorsal fin VI+I,9 or VII+I,9; anal fin II,8; head and body with ctenoid scales; pored lateral-line scales 23–25; preopercle ridge serrate, edges serrate; infraorbitals serrate; three supraneurals; supramaxilla absent; basisphenoid present; one pair of uroneurals or absent; three epurals; five free hypurals; parhypural separate; one or two supernumerary dorsal spines; caudal fin forked.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal-fin ray branched and segmented; ctenoid scales on predorsal, cheek, breast, two pelvic scales, and body; ctenoid scales on opercle and onto base of caudal fin; pored lateral-line scales with multiple pores; pectoral fin-rays 12–16; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; unbranched procurrent rays, longest segmented; teeth in one or multiple rows on premaxilla, dentary, vomer, palatine, all villiform or absent on palatine; six infraorbitals, bony shelf on third infraorbital; anterior ceratohyal smooth or notched; developed gill rakers 10–19; 10+14 vertebrae; 8 ribs; 8 epipleurals; low crest on PU2.
Distribution. Pristiapogon is wide spread from East Africa, Red Sea, Indian Ocean Islands, Indonesia, Philippines, Pacific islands to Hawaii, Japan, French Polynesia and Australia. One species of Pristicon Fraser 1972 is restricted to the West Pacific. Another species reaches out to some islands on the Pacific Plate. One species of Pristicon has been reported from the west coast of India, with a gap between India and Indonesia ( Suresh & Thomas 2007).
Remarks. This tribe contains two genera, Pristiapogon and Pristicon , corresponding to the clade VI in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). Although bootstrap values supporting the tribe were not so high (62 and 56% BPs in ML and MP analyses, respectively), this tribe is morphologically well-defined: relatively large body with serrations on the preopercle ridge (a likely synapomorphy), edges and infraorbitals (a likely synapomorphy). Species of Pristiapogon usually have a darkish single stripe and/or a variable basicaudal spot while Pristicon have bars or saddles under the dorsal fins and spots at the base of the caudal fin or on the opercle.
Tribe Rhabdamiini new name Fraser & Mabuchi
Type genus Rhabdamia Weber 1909
Diagnosis. Members of the Apogoninae : dorsal fin VI+I,9 or VII+I,10–11; anal fin II,11–13; head and body with weakly ctenoid or cycloid; pored lateral-line scales 23–24; preopercle ridge smooth, edges smooth; two supraneurals; supramaxilla absent; basisphenoid present or absent; anterior pair of uroneurals reduced or absent; three epurals; hypurals 1+2+3+4 fused, the plate fused to terminal centrum; parhypural separate; no autogenous haemal spines; 1–2 supernumerary dorsal spines; caudal fin forked.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal-fin ray branched and segmented; pectoral-fin rays 13–17; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; developed gill rakers 17–31; unbranched procurrent rays, longest segmented; villiform teeth in one or two rows on premaxilla, or with a few canines ( Bentuviaichthys ), one villiform row on dentary and vomer, one villiform row or absent on palatine; six infraorbitals, bony shelf reduce or absent on third infraorbital; stomach and intestine pale with melanophores and silvery peritoneum with melanophores; 10+14 vertebrae; low crest on PU2.
Distribution. No reviews of the species have been made, but members have been reported from East Africa, the Red Sea, islands in the Indian Ocean, the West Pacific and out onto the Pacific Plate.
Remarks. This tribe contains one genus Rhabdamia (including two subgenera Rhabdamia and Bentuviaichthys sensu Fraser 1972 ). There are six names (Appendix A). Three species are streamlined apogonines exploiting the near pelagic zone of coral reefs for food. Likely synapomorphies include smooth preopercle ridge and edges, reduced dentition, fused hypurals, and two supraneurals. All species are mostly translucent. In the present molecular analyses, they formed a robust monophyletic group (clade IX) with Yarica hyalosoma ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4) (see also Fig. 7 View FIGURE 7 ), but it is very unlikely based on morphological evidence (for details, see the remarks of the tribe Glossamiini ). In Fraser (1972), subgenus Rhabdamia was classified under the genus Rhabdamia together with the subgenera Bentuviaichthys and Verulux Fraser 1972 . The last lineage, Verulux (clade V), was placed far from the first lineage, Rhabdamia , in the present molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 ). Although monophyly between the two lineages was not rejected by AU test (H05 in Table 6), we recognized each of them in its own tribe for the present (for Verulux , see the tribe Veruluxini ). The present molecular analyses did not include Bentuviaichthys , but it was included in this tribe based on the following morphological characters: fused hypurals 1+2+3+4, epipleurals absent on last three ribs, single row of teeth on dentary, two supraneurals, slender, mostly translucent body and forked caudal fin. Two individuals of Rhabdamia gracilis from two distantly distributed populations ( Fiji and Ryukyu Islands) were paraphyletic to R. spilota Allen & Kuiter 1994 and genetically distant from each other, which may indicate needs of taxonomical revisions of them.
Tribe Siphamiini Smith 1955
Type genus Siphamia Weber 1909
Diagnosis. Members of the Apogoninae : dorsal fin VI–VII + I,7–11; anal fin II,7–11; pored lateral-line scales 0–24; preopercular ridge smooth, edges smooth to serrate; supraneurals 1–2; supramaxilla absent; basisphenoid absent; uroneurals absent; two epurals; hypurals 1+2 and 3+4 fused into two plates, upper plate fused to terminal centrum; parhypural separate; one or two supernumerary dorsal spines; caudal fin emarginate or forked; bacteria bioluminescent system from hyal region to along body above or past anal-fin base.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal-fin ray branched and segmented; ctenoid, cycloid or spinoid scales; median predorsal scales 0–6; pectoral rays 11–16; segmented principal caudal rays 9+8, 15 branched, upper and lower unbranched; unbranched procurrent rays, longest segmented; villiform teeth in one or multiple rows on premaxilla, dentary, vomer, palatine, or absent on palatine; developed gill rakers 6–18; lateral-line scales usually with a vertical row of free neuromasts; six infraorbitals, bony shelf on third infraorbital; anterior ceratohyal smooth or notched; developed gill rakers 10–26; 10+14 vertebrae; 8 ribs; 8 epipleurals; stomach, intestine and peritoneum generally pale with variously sized melanophores; low crest on PU2.
Distribution. Representatives of the tribe are found from East Africa, Red Sea, islands in the Indian Ocean, throughout the West Pacific to Japan, onto the Pacific Plate to French Polynesia and Australia ( Gon & Allen 2012).
Remarks. This tribe contains one genus Siphamia , corresponding to the clade VII in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). All species of Siphamia have bioluminescent bacteria in a specialized organ (a synapomorphy for the species) unique among apogonines. Smith (1955) proposed placing species of Siphamia in its own subfamily Siphamiinae . Our results based on analysis of five of the twenty-three species suggest that Siphamia roseigaster Ramsay & Ogilby 1887 could be recognized in its own genus Adenapogon McCulloch 1921 , because it was placed relatively far from the remaining species. Gon & Allen's (2012) results based on morphology suggest that two other Australian species belong in Adenapogon and that Fodifoa Whitley 1936 is available for another group of species. We defer to Ofer Gon who is continuing to work on relationships within this tribe ( Gon & Allen 2012).
Tribe Sphaeramiini new name Fraser & Mabuchi
Type genus Sphaeramia Fowler & Bean 1930
Diagnosis. Members of the Apogoninae : dorsal fin VII+I,9–10, VII(I)+I,9–14 or VIII+I,9–10 with eighth spine reduced to a tiny visible spine or a nubbin hidden under skin supported by a free sixth distal radial; anal fin II,8–13; head and body with ctenoid scales; pored lateral-line scales 23–27; preopercle ridge smooth, edges serrate; 2–3 supraneurals; supramaxilla absent; basisphenoid present; one pair of uroneurals; three epurals; five free hypurals or 1+2 fused and 3+4 fused with 3+4 fused to terminal centrum; parhypural separate; two autogenous haemal spines; two supernumerary dorsal spines; caudal fin forked, emarginate or truncate.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal-fin ray branched and segmented; ctenoid scales on predorsal, cheek, breast, two pelvic scales, and body; ctenoid scales on opercle and onto base of caudal fin; 1–3 predorsal scales cycloid or ctenoid; pored lateral-line scales with multiple pores; pectoral-fin rays 13–17; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; developed gill rakers 7–38; unbranched procurrent rays, longest segmented or spinous; third or fourth dorsal spine longest; 10+14 or 10+15 vertebrae; 8 ribs; 8–9 epipleurals; teeth in one or multiple rows on premaxilla, dentary, vomer, palatine, all villiform; six infraorbitals, bony shelf on third infraorbital; stomach and intestine pale or black with pale peritoneum; low crest on PU2.
Distribution. Species of Apogonichthyoides are found throughout the Indo-West Pacific from East Africa, Red Sea, India, some island of the Indian Ocean, throughout Indonesia, Philippines to Japan, New Guinea, Australia and the Solomon Islands. Some species have restricted distribution and some are rare in collections. Species of Jaydia Smith 1961 have a more continental distribution, mostly caught in trawls, and are widespread from Africa to Japan, New Guinea, Australia and larger islands in the Coral Sea. Representatives of Nectamia are found on reef habitats from East Africa, Red Sea, islands in the Indian Ocean, throughout the West Pacific to Japan, onto the Pacific Plate out to the Line Islands and French Polynesia. Sphaeramia , represented by two species, one found in mangrove habitats and one on coral reefs, also have wide distribution in the Indo-West Pacific. Quinca , represented by a single species, is known from northwestern coast of Australia.
Remarks. This tribe contains six genera, Apogonichthyoides , Jaydia , Nectamia , Pterapogon Koumans 1933 , Quinca and Sphaeramia , almost corresponding to the clade IV in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). First three of the six genera were formerly classified under Ostorhinchus , corresponding to "barred group" of Ostorhinchus ( Ostorhinchus II ) in Mabuchi et al. (2006). Two of the remaining three genera ( Pterapogon and Sphaeramia ) have distinct bar(s) on body. In addition to the six genera, corresponding molecular clade included three species of Ostorhinchus -2 ( O. hoevenii Bleeker 1854b , O. ishigakiensis Ida & Moyer 1974 , and O. rueppellii Günther 1859 ). But there are no morphological characters that support this association. We defer taxonomical action on this possible relationship for the present. This clade further included another species of the former Ostorhinchus ( Ostorhinchus -1). For this species, the latest authors ( Allen & Erdmann 2012) provisionally used Apogonichthyoides as its genus name, and we followed it here, but such application of the name made this genus paraphyletic in our molecular trees. Based on its morphological features, we tentatively included it [ Apogonichthyoides (?) melas ] in this tribe, but further study including more Sphaeramiini species will be needed to clarify its taxonomic status. In the present molecular analyses, phylogenetic position of Pterapogon remained unsettled ( Table 5). It was sister to the clade VII ( Siphamiini ) in ML tree ( Fig. 2 View FIGURE 2 ), sister to Ostorhinchus margaritophorus in MP tree-A ( Fig. 4 View FIGURE 4 ), and nested within the clade XII ( Ostorhinchini ) in the MP tree-B ( Fig. 5 View FIGURE 5 ). It formed a polytomy with O. margaritophorus , clade VII, clade VIII, and a large clade including clades IX–XII in BA tree ( Fig. 3 View FIGURE 3 ). Pterapogon has, however, a synapomorphy (spinous procurrent caudal rays) uniquely held with Sphaeramia as well as similarities in color patterns and body shape, but differs with fused hypurals 1+2 and 3+4. Based on these morphological features, we tentatively included this species in the tribe Sphaeramiini . Monophyly of them was not rejected by AU test (H02 in Table 6), although its inclusion within Ostorhinchini was also not rejected (H03 in Table 6). We had no good DNA extracts from tissue of Quinca . This monotypic genus has morphological characteristics and color pattern suggestive of a relationship with Apogonichthyoides , and was provisionally place in the Sphaeramiini .
Tribe Veruluxini new name Fraser & Mabuchi
Type genus Verulux Fraser 1972
Diagnosis. Members of the Apogoninae : dorsal fin VI–I,9; anal fin II,9; head and body with weakly ctenoid or cycloid scales; pored lateral-line scales 24; preopercle ridge smooth, edges smooth; one supraneural; supramaxilla absent; basisphenoid present; uroneurals absent; two epurals; fused hypurals 1+2+3+4, fused to terminal centrum; no autogenous haemal spines; one supernumerary dorsal spine; bioluminescent organ under cleithrum; caudal fin forked.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal fin-ray branched and segmented; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; developed gill rakers 12–15; unbranched procurrent rays, longest segmented; pectoral fin-rays 14–16; 10+14 vertebrae; 8 ribs; 5–6 epipleurals; teeth in one row on premaxilla, dentary, vomer, palatine, all villiform, or absent on palatine; six infraorbitals, bony shelf absent on third infraorbital; stomach and intestine black with silvery peritoneum; low crest on PU2.
Distribution. The single described species is known from East Africa, Red Sea, islands in the Indian Ocean, throughout the West Pacific to Japan, onto the Pacific Plate to Marshall Islands, Vanuatu, New Caledonia and Australia.
Remarks. This tribe contains only one species, Verulux cypselurus , corresponding to the clade V in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). Representatives of Verulux (two individuals from Ryukyu Islands and Seychelles) were monophyletic separated by a moderate genetic distance, which may indicate presence of cryptic species. This lineage was recovered far from Rhabdamia (tribe Rhabdamiini ), although their monophyly (genus Rhabdamia sensu Fraser 1972 ) was not rejected by AU test (H05 in Table 6). The single described species has a bioluminescent organ under cleithrum unlike Rhabdamia .
Tribe Zoramiini new name Fraser & Mabuchi
Type genus Zoramia Jordan 1917
Diagnosis. Members of the Apogoninae : dorsal fin VI–I,9; anal fin II,8–9; head and body with ctenoid scales; pored lateral-line scales 24; preopercle ridge smooth, edges serrate; three supraneurals; supramaxilla absent; basisphenoid present; anterior pair of uroneurals; three epurals; five free hypurals; parhypural separate; two autogenous haemal spines; one supernumerary dorsal spine; caudal fin forked.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal-fin ray branched and segmented; second dorsal spine longest, very long and filamentous in some species of both genera; ctenoid scales on predorsal, cheek, breast, two pelvic scales, and body; ctenoid scales on opercle and onto base of caudal fin; cycloid or ctenoid predorsal scales1–3; pored lateral-line scales simple with one pore above and one below midline; pectoral-fin rays 13–17; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; developed gill rakers 7–38; unbranched procurrent rays, longest segmented; 10+14 vertebrae; 8 ribs; 7–8 epipleurals; teeth in one, two or multiple rows on premaxilla, dentary, vomer, palatine, all villiform; six infraorbitals, bony shelf absent or on third infraorbital; stomach and intestine black with silvery peritoneum; low crest on PU2.
Distribution. Species of Zoramia are known from East Africa, Red Sea, islands in the Indian Ocean, throughout the West Pacific to Japan, onto the Pacific Plate to Marshall Islands, Samoa, Tonga, Solomon Islands, New Caledonia and Australia. Species of Fibramia are known from East Africa, some islands in the Indian Ocean, Sri Lanka, throughout the West Pacific to Philippines, Solomon Islands, Santa Cruz Islands, Fiji, Vanuatu, New Caledonia, Australia, Samoa and Tonga.
Remarks. This tribe contains two genera, a new genus Fibramia described below and Zoramia , corresponding to the clade VIII in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). Both genera have species with elongate second dorsal spine, a single supernumerary dorsal spine, and can be found in tidal fresh and brackish water, near shore waters and in lagoons. Fibramia corresponds to the clade of Ostorhinchus - 4 in the present study, and that of Ostorhinchus I in Mabuchi et al. (2006).
Tribe Glossamiini new name Fraser & Mabuchi
Type genus Glossamia Gill 1863
Diagnosis. Members of the Apogoninae : dorsal fin VI+I,8–10; anal fin II,8–10; pored lateral-line scales 24–50; preopercle ridge smooth edges smooth to serrate; three supraneurals; large supramaxilla present; basisphenoid present, reduced or absent; anterior pair of uroneurals; three epurals; five free hypurals; parhypural separate; two autogenous haemal spines; one supernumerary dorsal spine; caudal fin emarginate, truncate or rounded.
Other characteristics. first dorsal ray unbranched and first anal ray branched and segmented; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; unbranched procurrent rays, longest segmented; head and body with ctenoid scales or cycloid on nape, cheek, opercle, anterior pored lateral-line scales, ctenoid on breast, grading from cycloid to ctenoid posteriorly; pored lateral-line scales simple with one pore on upper side and one on lower side; pectoral fin-rays 13–16; developed gill rakers 7–15; 10+14 vertebrae; 8 ribs; 9 epineurals; teeth in multiple rows on premaxilla, dentary, vomer, palatine, all villiform, present or absent on ectopterygoid; six infraorbitals, bony shelf on third infraorbital; stomach and intestine pale with pale peritoneum; low crest on PU2.
Distribution. Species of Glossamia are found in tidal and flowing freshwaters of Australia and the island of New Guinea. One species is known from Australia (perhaps a species complex) and the rest on the island of New Guinea. The only described species of Yarica is found in tidal streams, flowing freshwater and lowland lakes from Myanmar, Andaman and Nicobar Islands, Thailand, Malaysia, Indonesia, Philippines, Australia, New Guinea, Solomon Islands to New Caledonia and out to Saipan.
Remarks. In the present molecular analyses, Glossamia did not form a robust monophyletic group with any other apogonines (see Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 and Table 5). For this genus, we gave tribe status. We included Yarica ( Fig. 7A View FIGURE 7 ) in this tribe based solely on morphological data. The present molecular analyses reproduced a relatively robust sister relationship between Yarica and Rhabdamia (clade IX). Species of Rhabdamia have a smaller mouth and slender translucent body with a forked caudal fin ( Fig. 7B View FIGURE 7 ) consistent with their marine reef pelagic habitat preferences and differ in their osteological characteristics as follows: 1 or 2 supraneurals; fused hypurals plate consisting of hypurals 1+2+3+4 to the terminal centrum; 1–2 rows of villiform teeth on premaxilla or some canine, 1 row of villiform teeth on dentary, vomer and palatine; 7 epineurals on ribs 1–7; shelf on third infraorbital reduced or absent; uroneurals reduced or absent; supramaxilla absent. The cephalic pores are much more complex for Rhabdamia and free neuromasts much less numerous than for Yarica ( Bergman 2004, Figs. 11, 29–30). Support for combining Glossamia and Yarica are similar body shapes, freshwater habitat preferences, a large supramaxilla of similar shape, a single supernumerary dorsal spine and 9 epineurals. In spite of non-monophyly in the obtained trees, AU test did not reject the monophyly of Glossamia and Yarica (H06, see Table 6). The wide spread distribution of Yarica indicates that this species has euryhaline characteristics imbedded in its life history.
Tribe Gymnapogonini Whitley 1941
Type genus Gymnapogon Regan 1905
Diagnosis. Members of the Apogoninae : VI+I,8–13; anal fin II,8–16; head and body naked or with large weakly ctenoid or cycloid scales; lateral-line scales 23–24 or lateral line a series of free neuromasts to base of caudal fin, interrupted mid-line lateral line present or absent; preopercle ridge smooth, edge with one or more spines at angle; 0–2 ossified supraneurals; supramaxilla, paired uroneurals and basisphenoid absent; 2–3 epurals; hypurals 1–2 fused, 3–4 separate or fused to urostyle; parhypural separate or fused to hypurals 1–2; caudal fin forked or rounded; pterosphenoid and parasphenoid joined excluding prootic from internal orbit rim.
Other characteristics. one supernumerary dorsal-fin spine; unbranched or branched first segmented dorsal-fin ray, branched first anal-fin ray; cycloid or weakly ctenoid scales on predorsal, cheek, breast, two pelvic scales, and body; cycloid scale on opercle and onto base of caudal fin; pectoral-fin rays 10–14; 9+8 segmented principal caudal rays, 13–15 branched, upper one or two and lower one or two unbranched; unbranched procurrent rays, longest segmented; some enlarged teeth on premaxilla and dentary, canine or villiform on vomer, villiform on palatine; six infraorbitals, bony shelf on third infraorbital present or absent; 1–2 postcleithra; anterior ceratohyal smooth or notched; 10+14 or 9+15 ( Cercamia ) vertebrae; 7–8 ribs; 0–6 epipleurals; low crest on PU2.
Distribution. Pseudamiops East Africa to Japan, Hawaii and French Polynesia, Gymnapogon widespread from the Red Sea, East Africa to Japan and French Polynesia, Lachneratus from East Africa to Hawaii and Tonga and Cercamia from the Eastern Indian Ocean to Japan and French Polynesia.
Remarks. This tribe contains four genera, Cercamia , Gymnapogon , Lachneratus and Pseudamiops , corresponding to the clade I in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). Species of the included genera are all translucent with many reductive morphologic characters. Although Pseudamiops was absent from the present analyses, we tentatively associated this genus with the Gymnapogonini based on its translucent body and the reductive morphological characters. Bergman (2004) noted: …"The cephalic lateralis system of Gymnapogon is very similar to that of Pseudamiops . A notable exception being that the preopercular and mandibular canal portions are confluent in Gymnapogon species. " Pseudamiops , with scales, lacks pored, notched or pitted lateral-line scales. Cercamia and Lachneratus have not previously been associated with Gymnapogon . Both genera have deciduous scales and both lack pored, notched or pitted lateral-line scales. Gymnapogon and Pseudamiops were formerly classified under Pseudamiinae , together with Pseudamia . As mentioned above in the remarks of the subfamily Pseudamiinae , the present molecular data significantly rejected their monophyly (H 14 in Table 6). In our molecular trees, two individuals of Cercamia cladara from two distantly distributed populations ( French Polynesia and Palau) were paraphyletic to Gymnapogon and genetically distant from each other, which may indicate needs of taxonomical revisions of them.
Tanaka (1915) described the new family Henicicthyidae for the new genus and single new species Henicichthys foraminosus . Tanaka's family name has been used only in the original publication. Whitley (1941) created the family Gymnapogonidae with Regan's genus as the type species. He noted that the oldest genus name '...becomes the root for the family name." Whitley's (1941) synonymy of the literature for Henicichthys has the last published use of this genus in 1939. Whitley (1941) recognized that Henicichthys foraminosus and Austalaphia annona Whitley 1936 are synonyms of Gymnapogon . The sole use of Gymnapogon has been continuous since 1941. Fowler (1944) and Lindberg (1971) used Gymnapogonidae . We regarded Gymnapogonini as an easily recognized tribal name among apogonids and is its preferred use.
Tribe Lepidamiini new name Fraser & Mabuchi
Type genus Lepidamia Gill 1863
Diagnosis. Members of the Apogoninae : dorsal fin VII(I)-I,9 or VIII-I,9; anal fin II,8; head and body with ctenoid scales; pored lateral-line scales 35–48; preopercle ridge smooth, edges serrate; three supraneurals; supramaxilla absent; basisphenoid present; one pair of reduced uroneurals present; three epurals; five free hypurals; parhypural separate; two autogenous haemal spines; two supernumerary dorsal spines, caudal fin forked.
Other characteristics. first fin ray in second dorsal fin branched and segmented; first anal-fin ray branched and segmented; ctenoid scales on predorsal, cheek, breast, four pelvic scales, and body; ctenoid scales on opercle and onto base of caudal fin; pored lateral-line scales with multiple pores; 15 branched, upper and lower unbranched; developed gill rakers 8–17; unbranched procurrent rays, longest segmented; 10+14 vertebrae; 8 ribs; 9 epipleurals; teeth in multiple rows on premaxilla, dentary, vomer, palatine, all villiform; six infraorbitals, bony shelf on third infraorbital; stomach and intestine pale, peritoneum pale; low crest on PU2.
Distribution. Lepidamia with four species has been collected only from continental marine waters from South Africa to China ( Gon 1995). Members of this tribe have not been collected in Australia or New Guinea. One specimen (USNM 175754, 96 mm SL) was collected in 1909 from Manila Bay, Luzon, Philippines. None have been reported elsewhere in the Philippines or again from Manila Bay. Perhaps the location is erroneous.
Remarks. We did not have tissue from any species. Based on known morphological characters including color patterns of the juveniles and adults, body shape and small body scales, we recognized Lepidamia in its own tribe for the present.
Tribe Ostorhinchini Whitley 1959
Type genus Ostorhinchus Lacepède 1802
Diagnosis. Members of the Apogoninae : dorsal fin VI+I,9 or VII+I,9; anal fin II,8–9; head and body with ctenoid scales; pored lateral-line scales 6–26; preopercle ridge smooth, edges serrate; 2–3 supraneurals; supramaxilla absent; basisphenoid present; one pair of reduced uroneurals present or absent; three epurals; five free hypurals or 1–2 fused; parhypural separate; caudal fin forked; head and/or body with one or more light or dark stripes, bars rarely present, basicaudal spot or broad mark present or absent.
Other characteristics. one or two supernumerary dorsal spines; branched first segmented fin ray in second dorsal-fin; first anal-fin ray branched and segmented; ctenoid scales on predorsal, cheek, breast, two pelvic scales, and body; ctenoid scale on opercle and onto base of caudal fin; pored lateral-line scales with multiple pores; pectoral fin-rays 11–16; three supraneurals; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; unbranched procurrent rays, longest segmented; teeth in one or multiple rows on premaxilla, dentary, vomer, palatine, all villiform or absent on palatine, sometimes enlarged, but not canine on side of dentary; six infraorbitals, bony shelf on third infraorbital; anterior ceratohyal smooth or notched; developed gill rakers 10–26; 10+14 vertebrae; 8 ribs; 8 epipleurals; blackish stomachs and intestines; low crest on PU2.
Distribution. The restricted Ostorhinchus is widespread throughout the Indo-Pacific from the Red Sea, East Africa to Japan, Hawaii, Easter Island and French Polynesia.
Remarks. Monophyly of recent Ostorhinchus (species indicated by blue in Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 ) was significantly rejected by AU test (H 16 in Table 6). This tribe, however, corresponds to one of the five lineages of the recent Ostorhinchus , clade XII. It included most species of the recent Ostorhinchus : many striped species of Ostorhinchus (species of Ostorhinchus -5) and Brephamia parvula Smith & Radcliffe in Radcliffe 1912 ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). Other species of the recent Ostorhinchus were divided into four lineages, Ostorhinchus -1, 2, 3 and 4. They were all placed far from the clade XII. Except for Ostorhinchus -3 ( O. margaritophorus ), monophyly between this tribe (clade XII) and each of the remaining lineages was rejected by AU tests (H07, 08 and 11 in Table 6). Although the bootstrap values supporting the clade XII were not so high (<50%) both in the ML ( Fig. 2 View FIGURE 2 ) and MP ( Figs. 4 View FIGURE 4 and 5 View FIGURE 5 ) trees ( Table 4), and further the clade was divided into two separate clades in the BA tree ( Fig. 3 View FIGURE 3 ), we are treating this group as a single genus, based on the typical color pattern (stripes on body and/or head) shared by most of the included species. This group corresponds to the "striped group" of Ostorhinchus ( Ostorhinchus III ) in Mabuchi et al. (2006). One of the other recent Ostorhinchus lineages, O. margaritophorus , has long stripes, short bars between two long stripes on body and fused hypurals 1+2. Phylogenetic position of this species remains unsettled in the molecular trees ( Table 5), but monophyly between this species and the members of this tribe was not rejected by AU test (H01 in Table 6). Based on its featured color pattern, we tentatively included this species in this tribe. Apogon ( Brephamia) parvula has a very similar species within this tribe. It is Ostorhinchus neotes . Although they were not sister to each other in the molecular trees, their monophyly was not rejected by AU test (H 12 in Table 6). While recognition of Brephamia Jordan in Jordan & Jordan (1922) as a subgenus or genus needs further evaluation, we synonymized it under the restricted Ostorhinchus for the present. Likely synapomorphies may include color pattern groupings, probably at the subgenus level. At the genus level, with the exception of the loss of tiny first dorsal-fin spines, all species have 7 visible first dorsal spines, a serrated preopercular edge and most have blackish stomachs and intestines.
There are two available names ( Gronovichthys Whitley 1929 ; Lovamia Whitley 1930 ) which could be used in the future. There are at least 93 species in the group. We have sampled 31 species. Whitley (1959) first used the present name at family level Ostorhinchidae. The name appeared once. We use the name at the tribal level.
Tribe Pristiapogonini new name Fraser & Mabuchi
Type genus Pristiapogon Klunzinger 1870
Diagnosis. Members of the Apogoninae : dorsal fin VI+I,9 or VII+I,9; anal fin II,8; head and body with ctenoid scales; pored lateral-line scales 23–25; preopercle ridge serrate, edges serrate; infraorbitals serrate; three supraneurals; supramaxilla absent; basisphenoid present; one pair of uroneurals or absent; three epurals; five free hypurals; parhypural separate; one or two supernumerary dorsal spines; caudal fin forked.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal-fin ray branched and segmented; ctenoid scales on predorsal, cheek, breast, two pelvic scales, and body; ctenoid scales on opercle and onto base of caudal fin; pored lateral-line scales with multiple pores; pectoral fin-rays 12–16; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; unbranched procurrent rays, longest segmented; teeth in one or multiple rows on premaxilla, dentary, vomer, palatine, all villiform or absent on palatine; six infraorbitals, bony shelf on third infraorbital; anterior ceratohyal smooth or notched; developed gill rakers 10–19; 10+14 vertebrae; 8 ribs; 8 epipleurals; low crest on PU2.
Distribution. Pristiapogon is wide spread from East Africa, Red Sea, Indian Ocean Islands, Indonesia, Philippines, Pacific islands to Hawaii, Japan, French Polynesia and Australia. One species of Pristicon Fraser 1972 is restricted to the West Pacific. Another species reaches out to some islands on the Pacific Plate. One species of Pristicon has been reported from the west coast of India, with a gap between India and Indonesia ( Suresh & Thomas 2007).
Remarks. This tribe contains two genera, Pristiapogon and Pristicon , corresponding to the clade VI in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). Although bootstrap values supporting the tribe were not so high (62 and 56% BPs in ML and MP analyses, respectively), this tribe is morphologically well-defined: relatively large body with serrations on the preopercle ridge (a likely synapomorphy), edges and infraorbitals (a likely synapomorphy). Species of Pristiapogon usually have a darkish single stripe and/or a variable basicaudal spot while Pristicon have bars or saddles under the dorsal fins and spots at the base of the caudal fin or on the opercle.
Tribe Rhabdamiini new name Fraser & Mabuchi
Type genus Rhabdamia Weber 1909
Diagnosis. Members of the Apogoninae : dorsal fin VI+I,9 or VII+I,10–11; anal fin II,11–13; head and body with weakly ctenoid or cycloid; pored lateral-line scales 23–24; preopercle ridge smooth, edges smooth; two supraneurals; supramaxilla absent; basisphenoid present or absent; anterior pair of uroneurals reduced or absent; three epurals; hypurals 1+2+3+4 fused, the plate fused to terminal centrum; parhypural separate; no autogenous haemal spines; 1–2 supernumerary dorsal spines; caudal fin forked.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal-fin ray branched and segmented; pectoral-fin rays 13–17; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; developed gill rakers 17–31; unbranched procurrent rays, longest segmented; villiform teeth in one or two rows on premaxilla, or with a few canines ( Bentuviaichthys ), one villiform row on dentary and vomer, one villiform row or absent on palatine; six infraorbitals, bony shelf reduce or absent on third infraorbital; stomach and intestine pale with melanophores and silvery peritoneum with melanophores; 10+14 vertebrae; low crest on PU2.
Distribution. No reviews of the species have been made, but members have been reported from East Africa, the Red Sea, islands in the Indian Ocean, the West Pacific and out onto the Pacific Plate.
Remarks. This tribe contains one genus Rhabdamia (including two subgenera Rhabdamia and Bentuviaichthys sensu Fraser 1972 ). There are six names (Appendix A). Three species are streamlined apogonines exploiting the near pelagic zone of coral reefs for food. Likely synapomorphies include smooth preopercle ridge and edges, reduced dentition, fused hypurals, and two supraneurals. All species are mostly translucent. In the present molecular analyses, they formed a robust monophyletic group (clade IX) with Yarica hyalosoma ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4) (see also Fig. 7 View FIGURE 7 ), but it is very unlikely based on morphological evidence (for details, see the remarks of the tribe Glossamiini ). In Fraser (1972), subgenus Rhabdamia was classified under the genus Rhabdamia together with the subgenera Bentuviaichthys and Verulux Fraser 1972 . The last lineage, Verulux (clade V), was placed far from the first lineage, Rhabdamia , in the present molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 ). Although monophyly between the two lineages was not rejected by AU test (H05 in Table 6), we recognized each of them in its own tribe for the present (for Verulux , see the tribe Veruluxini ). The present molecular analyses did not include Bentuviaichthys , but it was included in this tribe based on the following morphological characters: fused hypurals 1+2+3+4, epipleurals absent on last three ribs, single row of teeth on dentary, two supraneurals, slender, mostly translucent body and forked caudal fin. Two individuals of Rhabdamia gracilis from two distantly distributed populations ( Fiji and Ryukyu Islands) were paraphyletic to R. spilota Allen & Kuiter 1994 and genetically distant from each other, which may indicate needs of taxonomical revisions of them.
Tribe Siphamiini Smith 1955
Type genus Siphamia Weber 1909
Diagnosis. Members of the Apogoninae : dorsal fin VI–VII + I,7–11; anal fin II,7–11; pored lateral-line scales 0–24; preopercular ridge smooth, edges smooth to serrate; supraneurals 1–2; supramaxilla absent; basisphenoid absent; uroneurals absent; two epurals; hypurals 1+2 and 3+4 fused into two plates, upper plate fused to terminal centrum; parhypural separate; one or two supernumerary dorsal spines; caudal fin emarginate or forked; bacteria bioluminescent system from hyal region to along body above or past anal-fin base.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal-fin ray branched and segmented; ctenoid, cycloid or spinoid scales; median predorsal scales 0–6; pectoral rays 11–16; segmented principal caudal rays 9+8, 15 branched, upper and lower unbranched; unbranched procurrent rays, longest segmented; villiform teeth in one or multiple rows on premaxilla, dentary, vomer, palatine, or absent on palatine; developed gill rakers 6–18; lateral-line scales usually with a vertical row of free neuromasts; six infraorbitals, bony shelf on third infraorbital; anterior ceratohyal smooth or notched; developed gill rakers 10–26; 10+14 vertebrae; 8 ribs; 8 epipleurals; stomach, intestine and peritoneum generally pale with variously sized melanophores; low crest on PU2.
Distribution. Representatives of the tribe are found from East Africa, Red Sea, islands in the Indian Ocean, throughout the West Pacific to Japan, onto the Pacific Plate to French Polynesia and Australia ( Gon & Allen 2012).
Remarks. This tribe contains one genus Siphamia , corresponding to the clade VII in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). All species of Siphamia have bioluminescent bacteria in a specialized organ (a synapomorphy for the species) unique among apogonines. Smith (1955) proposed placing species of Siphamia in its own subfamily Siphamiinae . Our results based on analysis of five of the twenty-three species suggest that Siphamia roseigaster Ramsay & Ogilby 1887 could be recognized in its own genus Adenapogon McCulloch 1921 , because it was placed relatively far from the remaining species. Gon & Allen's (2012) results based on morphology suggest that two other Australian species belong in Adenapogon and that Fodifoa Whitley 1936 is available for another group of species. We defer to Ofer Gon who is continuing to work on relationships within this tribe ( Gon & Allen 2012).
Tribe Sphaeramiini new name Fraser & Mabuchi
Type genus Sphaeramia Fowler & Bean 1930
Diagnosis. Members of the Apogoninae : dorsal fin VII+I,9–10, VII(I)+I,9–14 or VIII+I,9–10 with eighth spine reduced to a tiny visible spine or a nubbin hidden under skin supported by a free sixth distal radial; anal fin II,8–13; head and body with ctenoid scales; pored lateral-line scales 23–27; preopercle ridge smooth, edges serrate; 2–3 supraneurals; supramaxilla absent; basisphenoid present; one pair of uroneurals; three epurals; five free hypurals or 1+2 fused and 3+4 fused with 3+4 fused to terminal centrum; parhypural separate; two autogenous haemal spines; two supernumerary dorsal spines; caudal fin forked, emarginate or truncate.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal-fin ray branched and segmented; ctenoid scales on predorsal, cheek, breast, two pelvic scales, and body; ctenoid scales on opercle and onto base of caudal fin; 1–3 predorsal scales cycloid or ctenoid; pored lateral-line scales with multiple pores; pectoral-fin rays 13–17; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; developed gill rakers 7–38; unbranched procurrent rays, longest segmented or spinous; third or fourth dorsal spine longest; 10+14 or 10+15 vertebrae; 8 ribs; 8–9 epipleurals; teeth in one or multiple rows on premaxilla, dentary, vomer, palatine, all villiform; six infraorbitals, bony shelf on third infraorbital; stomach and intestine pale or black with pale peritoneum; low crest on PU2.
Distribution. Species of Apogonichthyoides are found throughout the Indo-West Pacific from East Africa, Red Sea, India, some island of the Indian Ocean, throughout Indonesia, Philippines to Japan, New Guinea, Australia and the Solomon Islands. Some species have restricted distribution and some are rare in collections. Species of Jaydia Smith 1961 have a more continental distribution, mostly caught in trawls, and are widespread from Africa to Japan, New Guinea, Australia and larger islands in the Coral Sea. Representatives of Nectamia are found on reef habitats from East Africa, Red Sea, islands in the Indian Ocean, throughout the West Pacific to Japan, onto the Pacific Plate out to the Line Islands and French Polynesia. Sphaeramia , represented by two species, one found in mangrove habitats and one on coral reefs, also have wide distribution in the Indo-West Pacific. Quinca , represented by a single species, is known from northwestern coast of Australia.
Remarks. This tribe contains six genera, Apogonichthyoides , Jaydia , Nectamia , Pterapogon Koumans 1933 , Quinca and Sphaeramia , almost corresponding to the clade IV in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). First three of the six genera were formerly classified under Ostorhinchus , corresponding to "barred group" of Ostorhinchus ( Ostorhinchus II ) in Mabuchi et al. (2006). Two of the remaining three genera ( Pterapogon and Sphaeramia ) have distinct bar(s) on body. In addition to the six genera, corresponding molecular clade included three species of Ostorhinchus -2 ( O. hoevenii Bleeker 1854b , O. ishigakiensis Ida & Moyer 1974 , and O. rueppellii Günther 1859 ). But there are no morphological characters that support this association. We defer taxonomical action on this possible relationship for the present. This clade further included another species of the former Ostorhinchus ( Ostorhinchus -1). For this species, the latest authors ( Allen & Erdmann 2012) provisionally used Apogonichthyoides as its genus name, and we followed it here, but such application of the name made this genus paraphyletic in our molecular trees. Based on its morphological features, we tentatively included it [ Apogonichthyoides (?) melas ] in this tribe, but further study including more Sphaeramiini species will be needed to clarify its taxonomic status. In the present molecular analyses, phylogenetic position of Pterapogon remained unsettled ( Table 5). It was sister to the clade VII ( Siphamiini ) in ML tree ( Fig. 2 View FIGURE 2 ), sister to Ostorhinchus margaritophorus in MP tree-A ( Fig. 4 View FIGURE 4 ), and nested within the clade XII ( Ostorhinchini ) in the MP tree-B ( Fig. 5 View FIGURE 5 ). It formed a polytomy with O. margaritophorus , clade VII, clade VIII, and a large clade including clades IX–XII in BA tree ( Fig. 3 View FIGURE 3 ). Pterapogon has, however, a synapomorphy (spinous procurrent caudal rays) uniquely held with Sphaeramia as well as similarities in color patterns and body shape, but differs with fused hypurals 1+2 and 3+4. Based on these morphological features, we tentatively included this species in the tribe Sphaeramiini . Monophyly of them was not rejected by AU test (H02 in Table 6), although its inclusion within Ostorhinchini was also not rejected (H03 in Table 6). We had no good DNA extracts from tissue of Quinca . This monotypic genus has morphological characteristics and color pattern suggestive of a relationship with Apogonichthyoides , and was provisionally place in the Sphaeramiini .
Tribe Veruluxini new name Fraser & Mabuchi
Type genus Verulux Fraser 1972
Diagnosis. Members of the Apogoninae : dorsal fin VI–I,9; anal fin II,9; head and body with weakly ctenoid or cycloid scales; pored lateral-line scales 24; preopercle ridge smooth, edges smooth; one supraneural; supramaxilla absent; basisphenoid present; uroneurals absent; two epurals; fused hypurals 1+2+3+4, fused to terminal centrum; no autogenous haemal spines; one supernumerary dorsal spine; bioluminescent organ under cleithrum; caudal fin forked.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal fin-ray branched and segmented; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; developed gill rakers 12–15; unbranched procurrent rays, longest segmented; pectoral fin-rays 14–16; 10+14 vertebrae; 8 ribs; 5–6 epipleurals; teeth in one row on premaxilla, dentary, vomer, palatine, all villiform, or absent on palatine; six infraorbitals, bony shelf absent on third infraorbital; stomach and intestine black with silvery peritoneum; low crest on PU2.
Distribution. The single described species is known from East Africa, Red Sea, islands in the Indian Ocean, throughout the West Pacific to Japan, onto the Pacific Plate to Marshall Islands, Vanuatu, New Caledonia and Australia.
Remarks. This tribe contains only one species, Verulux cypselurus , corresponding to the clade V in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). Representatives of Verulux (two individuals from Ryukyu Islands and Seychelles) were monophyletic separated by a moderate genetic distance, which may indicate presence of cryptic species. This lineage was recovered far from Rhabdamia (tribe Rhabdamiini ), although their monophyly (genus Rhabdamia sensu Fraser 1972 ) was not rejected by AU test (H05 in Table 6). The single described species has a bioluminescent organ under cleithrum unlike Rhabdamia .
Tribe Zoramiini new name Fraser & Mabuchi
Type genus Zoramia Jordan 1917
Diagnosis. Members of the Apogoninae : dorsal fin VI–I,9; anal fin II,8–9; head and body with ctenoid scales; pored lateral-line scales 24; preopercle ridge smooth, edges serrate; three supraneurals; supramaxilla absent; basisphenoid present; anterior pair of uroneurals; three epurals; five free hypurals; parhypural separate; two autogenous haemal spines; one supernumerary dorsal spine; caudal fin forked.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal-fin ray branched and segmented; second dorsal spine longest, very long and filamentous in some species of both genera; ctenoid scales on predorsal, cheek, breast, two pelvic scales, and body; ctenoid scales on opercle and onto base of caudal fin; cycloid or ctenoid predorsal scales1–3; pored lateral-line scales simple with one pore above and one below midline; pectoral-fin rays 13–17; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; developed gill rakers 7–38; unbranched procurrent rays, longest segmented; 10+14 vertebrae; 8 ribs; 7–8 epipleurals; teeth in one, two or multiple rows on premaxilla, dentary, vomer, palatine, all villiform; six infraorbitals, bony shelf absent or on third infraorbital; stomach and intestine black with silvery peritoneum; low crest on PU2.
Distribution. Species of Zoramia are known from East Africa, Red Sea, islands in the Indian Ocean, throughout the West Pacific to Japan, onto the Pacific Plate to Marshall Islands, Samoa, Tonga, Solomon Islands, New Caledonia and Australia. Species of Fibramia are known from East Africa, some islands in the Indian Ocean, Sri Lanka, throughout the West Pacific to Philippines, Solomon Islands, Santa Cruz Islands, Fiji, Vanuatu, New Caledonia, Australia, Samoa and Tonga.
Remarks. This tribe contains two genera, a new genus Fibramia described below and Zoramia , corresponding to the clade VIII in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). Both genera have species with elongate second dorsal spine, a single supernumerary dorsal spine, and can be found in tidal fresh and brackish water, near shore waters and in lagoons. Fibramia corresponds to the clade of Ostorhinchus - 4 in the present study, and that of Ostorhinchus I in Mabuchi et al. (2006).
Tribe Gymnapogonini Whitley 1941
Type genus Gymnapogon Regan 1905
Diagnosis. Members of the Apogoninae : VI+I,8–13; anal fin II,8–16; head and body naked or with large weakly ctenoid or cycloid scales; lateral-line scales 23–24 or lateral line a series of free neuromasts to base of caudal fin, interrupted mid-line lateral line present or absent; preopercle ridge smooth, edge with one or more spines at angle; 0–2 ossified supraneurals; supramaxilla, paired uroneurals and basisphenoid absent; 2–3 epurals; hypurals 1–2 fused, 3–4 separate or fused to urostyle; parhypural separate or fused to hypurals 1–2; caudal fin forked or rounded; pterosphenoid and parasphenoid joined excluding prootic from internal orbit rim.
Other characteristics. one supernumerary dorsal-fin spine; unbranched or branched first segmented dorsal-fin ray, branched first anal-fin ray; cycloid or weakly ctenoid scales on predorsal, cheek, breast, two pelvic scales, and body; cycloid scale on opercle and onto base of caudal fin; pectoral-fin rays 10–14; 9+8 segmented principal caudal rays, 13–15 branched, upper one or two and lower one or two unbranched; unbranched procurrent rays, longest segmented; some enlarged teeth on premaxilla and dentary, canine or villiform on vomer, villiform on palatine; six infraorbitals, bony shelf on third infraorbital present or absent; 1–2 postcleithra; anterior ceratohyal smooth or notched; 10+14 or 9+15 ( Cercamia ) vertebrae; 7–8 ribs; 0–6 epipleurals; low crest on PU2.
Distribution. Pseudamiops East Africa to Japan, Hawaii and French Polynesia, Gymnapogon widespread from the Red Sea, East Africa to Japan and French Polynesia, Lachneratus from East Africa to Hawaii and Tonga and Cercamia from the Eastern Indian Ocean to Japan and French Polynesia.
Remarks. This tribe contains four genera, Cercamia , Gymnapogon , Lachneratus and Pseudamiops , corresponding to the clade I in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). Species of the included genera are all translucent with many reductive morphologic characters. Although Pseudamiops was absent from the present analyses, we tentatively associated this genus with the Gymnapogonini based on its translucent body and the reductive morphological characters. Bergman (2004) noted: …"The cephalic lateralis system of Gymnapogon is very similar to that of Pseudamiops . A notable exception being that the preopercular and mandibular canal portions are confluent in Gymnapogon species. " Pseudamiops , with scales, lacks pored, notched or pitted lateral-line scales. Cercamia and Lachneratus have not previously been associated with Gymnapogon . Both genera have deciduous scales and both lack pored, notched or pitted lateral-line scales. Gymnapogon and Pseudamiops were formerly classified under Pseudamiinae , together with Pseudamia . As mentioned above in the remarks of the subfamily Pseudamiinae , the present molecular data significantly rejected their monophyly (H 14 in Table 6). In our molecular trees, two individuals of Cercamia cladara from two distantly distributed populations ( French Polynesia and Palau) were paraphyletic to Gymnapogon and genetically distant from each other, which may indicate needs of taxonomical revisions of them.
Tanaka (1915) described the new family Henicicthyidae for the new genus and single new species Henicichthys foraminosus . Tanaka's family name has been used only in the original publication. Whitley (1941) created the family Gymnapogonidae with Regan's genus as the type species. He noted that the oldest genus name '...becomes the root for the family name." Whitley's (1941) synonymy of the literature for Henicichthys has the last published use of this genus in 1939. Whitley (1941) recognized that Henicichthys foraminosus and Austalaphia annona Whitley 1936 are synonyms of Gymnapogon . The sole use of Gymnapogon has been continuous since 1941. Fowler (1944) and Lindberg (1971) used Gymnapogonidae . We regarded Gymnapogonini as an easily recognized tribal name among apogonids and is its preferred use.
Tribe Lepidamiini new name Fraser & Mabuchi
Type genus Lepidamia Gill 1863
Diagnosis. Members of the Apogoninae : dorsal fin VII(I)-I,9 or VIII-I,9; anal fin II,8; head and body with ctenoid scales; pored lateral-line scales 35–48; preopercle ridge smooth, edges serrate; three supraneurals; supramaxilla absent; basisphenoid present; one pair of reduced uroneurals present; three epurals; five free hypurals; parhypural separate; two autogenous haemal spines; two supernumerary dorsal spines, caudal fin forked.
Other characteristics. first fin ray in second dorsal fin branched and segmented; first anal-fin ray branched and segmented; ctenoid scales on predorsal, cheek, breast, four pelvic scales, and body; ctenoid scales on opercle and onto base of caudal fin; pored lateral-line scales with multiple pores; 15 branched, upper and lower unbranched; developed gill rakers 8–17; unbranched procurrent rays, longest segmented; 10+14 vertebrae; 8 ribs; 9 epipleurals; teeth in multiple rows on premaxilla, dentary, vomer, palatine, all villiform; six infraorbitals, bony shelf on third infraorbital; stomach and intestine pale, peritoneum pale; low crest on PU2.
Distribution. Lepidamia with four species has been collected only from continental marine waters from South Africa to China ( Gon 1995). Members of this tribe have not been collected in Australia or New Guinea. One specimen (USNM 175754, 96 mm SL) was collected in 1909 from Manila Bay, Luzon, Philippines. None have been reported elsewhere in the Philippines or again from Manila Bay. Perhaps the location is erroneous.
Remarks. We did not have tissue from any species. Based on known morphological characters including color patterns of the juveniles and adults, body shape and small body scales, we recognized Lepidamia in its own tribe for the present.
Tribe Ostorhinchini Whitley 1959
Type genus Ostorhinchus Lacepède 1802
Diagnosis. Members of the Apogoninae : dorsal fin VI+I,9 or VII+I,9; anal fin II,8–9; head and body with ctenoid scales; pored lateral-line scales 6–26; preopercle ridge smooth, edges serrate; 2–3 supraneurals; supramaxilla absent; basisphenoid present; one pair of reduced uroneurals present or absent; three epurals; five free hypurals or 1–2 fused; parhypural separate; caudal fin forked; head and/or body with one or more light or dark stripes, bars rarely present, basicaudal spot or broad mark present or absent.
Other characteristics. one or two supernumerary dorsal spines; branched first segmented fin ray in second dorsal-fin; first anal-fin ray branched and segmented; ctenoid scales on predorsal, cheek, breast, two pelvic scales, and body; ctenoid scale on opercle and onto base of caudal fin; pored lateral-line scales with multiple pores; pectoral fin-rays 11–16; three supraneurals; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; unbranched procurrent rays, longest segmented; teeth in one or multiple rows on premaxilla, dentary, vomer, palatine, all villiform or absent on palatine, sometimes enlarged, but not canine on side of dentary; six infraorbitals, bony shelf on third infraorbital; anterior ceratohyal smooth or notched; developed gill rakers 10–26; 10+14 vertebrae; 8 ribs; 8 epipleurals; blackish stomachs and intestines; low crest on PU2.
Distribution. The restricted Ostorhinchus is widespread throughout the Indo-Pacific from the Red Sea, East Africa to Japan, Hawaii, Easter Island and French Polynesia.
Remarks. Monophyly of recent Ostorhinchus (species indicated by blue in Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 ) was significantly rejected by AU test (H 16 in Table 6). This tribe, however, corresponds to one of the five lineages of the recent Ostorhinchus , clade XII. It included most species of the recent Ostorhinchus : many striped species of Ostorhinchus (species of Ostorhinchus -5) and Brephamia parvula Smith & Radcliffe in Radcliffe 1912 ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). Other species of the recent Ostorhinchus were divided into four lineages, Ostorhinchus -1, 2, 3 and 4. They were all placed far from the clade XII. Except for Ostorhinchus -3 ( O. margaritophorus ), monophyly between this tribe (clade XII) and each of the remaining lineages was rejected by AU tests (H07, 08 and 11 in Table 6). Although the bootstrap values supporting the clade XII were not so high (<50%) both in the ML ( Fig. 2 View FIGURE 2 ) and MP ( Figs. 4 View FIGURE 4 and 5 View FIGURE 5 ) trees ( Table 4), and further the clade was divided into two separate clades in the BA tree ( Fig. 3 View FIGURE 3 ), we are treating this group as a single genus, based on the typical color pattern (stripes on body and/or head) shared by most of the included species. This group corresponds to the "striped group" of Ostorhinchus ( Ostorhinchus III ) in Mabuchi et al. (2006). One of the other recent Ostorhinchus lineages, O. margaritophorus , has long stripes, short bars between two long stripes on body and fused hypurals 1+2. Phylogenetic position of this species remains unsettled in the molecular trees ( Table 5), but monophyly between this species and the members of this tribe was not rejected by AU test (H01 in Table 6). Based on its featured color pattern, we tentatively included this species in this tribe. Apogon ( Brephamia) parvula has a very similar species within this tribe. It is Ostorhinchus neotes . Although they were not sister to each other in the molecular trees, their monophyly was not rejected by AU test (H 12 in Table 6). While recognition of Brephamia Jordan in Jordan & Jordan (1922) as a subgenus or genus needs further evaluation, we synonymized it under the restricted Ostorhinchus for the present. Likely synapomorphies may include color pattern groupings, probably at the subgenus level. At the genus level, with the exception of the loss of tiny first dorsal-fin spines, all species have 7 visible first dorsal spines, a serrated preopercular edge and most have blackish stomachs and intestines.
There are two available names ( Gronovichthys Whitley 1929 ; Lovamia Whitley 1930 ) which could be used in the future. There are at least 93 species in the group. We have sampled 31 species. Whitley (1959) first used the present name at family level Ostorhinchidae. The name appeared once. We use the name at the tribal level.
Tribe Pristiapogonini new name Fraser & Mabuchi
Type genus Pristiapogon Klunzinger 1870
Diagnosis. Members of the Apogoninae : dorsal fin VI+I,9 or VII+I,9; anal fin II,8; head and body with ctenoid scales; pored lateral-line scales 23–25; preopercle ridge serrate, edges serrate; infraorbitals serrate; three supraneurals; supramaxilla absent; basisphenoid present; one pair of uroneurals or absent; three epurals; five free hypurals; parhypural separate; one or two supernumerary dorsal spines; caudal fin forked.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal-fin ray branched and segmented; ctenoid scales on predorsal, cheek, breast, two pelvic scales, and body; ctenoid scales on opercle and onto base of caudal fin; pored lateral-line scales with multiple pores; pectoral fin-rays 12–16; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; unbranched procurrent rays, longest segmented; teeth in one or multiple rows on premaxilla, dentary, vomer, palatine, all villiform or absent on palatine; six infraorbitals, bony shelf on third infraorbital; anterior ceratohyal smooth or notched; developed gill rakers 10–19; 10+14 vertebrae; 8 ribs; 8 epipleurals; low crest on PU2.
Distribution. Pristiapogon is wide spread from East Africa, Red Sea, Indian Ocean Islands, Indonesia, Philippines, Pacific islands to Hawaii, Japan, French Polynesia and Australia. One species of Pristicon Fraser 1972 is restricted to the West Pacific. Another species reaches out to some islands on the Pacific Plate. One species of Pristicon has been reported from the west coast of India, with a gap between India and Indonesia ( Suresh & Thomas 2007).
Remarks. This tribe contains two genera, Pristiapogon and Pristicon , corresponding to the clade VI in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). Although bootstrap values supporting the tribe were not so high (62 and 56% BPs in ML and MP analyses, respectively), this tribe is morphologically well-defined: relatively large body with serrations on the preopercle ridge (a likely synapomorphy), edges and infraorbitals (a likely synapomorphy). Species of Pristiapogon usually have a darkish single stripe and/or a variable basicaudal spot while Pristicon have bars or saddles under the dorsal fins and spots at the base of the caudal fin or on the opercle.
Tribe Rhabdamiini new name Fraser & Mabuchi
Type genus Rhabdamia Weber 1909
Diagnosis. Members of the Apogoninae : dorsal fin VI+I,9 or VII+I,10–11; anal fin II,11–13; head and body with weakly ctenoid or cycloid; pored lateral-line scales 23–24; preopercle ridge smooth, edges smooth; two supraneurals; supramaxilla absent; basisphenoid present or absent; anterior pair of uroneurals reduced or absent; three epurals; hypurals 1+2+3+4 fused, the plate fused to terminal centrum; parhypural separate; no autogenous haemal spines; 1–2 supernumerary dorsal spines; caudal fin forked.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal-fin ray branched and segmented; pectoral-fin rays 13–17; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; developed gill rakers 17–31; unbranched procurrent rays, longest segmented; villiform teeth in one or two rows on premaxilla, or with a few canines ( Bentuviaichthys ), one villiform row on dentary and vomer, one villiform row or absent on palatine; six infraorbitals, bony shelf reduce or absent on third infraorbital; stomach and intestine pale with melanophores and silvery peritoneum with melanophores; 10+14 vertebrae; low crest on PU2.
Distribution. No reviews of the species have been made, but members have been reported from East Africa, the Red Sea, islands in the Indian Ocean, the West Pacific and out onto the Pacific Plate.
Remarks. This tribe contains one genus Rhabdamia (including two subgenera Rhabdamia and Bentuviaichthys sensu Fraser 1972 ). There are six names (Appendix A). Three species are streamlined apogonines exploiting the near pelagic zone of coral reefs for food. Likely synapomorphies include smooth preopercle ridge and edges, reduced dentition, fused hypurals, and two supraneurals. All species are mostly translucent. In the present molecular analyses, they formed a robust monophyletic group (clade IX) with Yarica hyalosoma ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4) (see also Fig. 7 View FIGURE 7 ), but it is very unlikely based on morphological evidence (for details, see the remarks of the tribe Glossamiini ). In Fraser (1972), subgenus Rhabdamia was classified under the genus Rhabdamia together with the subgenera Bentuviaichthys and Verulux Fraser 1972 . The last lineage, Verulux (clade V), was placed far from the first lineage, Rhabdamia , in the present molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 ). Although monophyly between the two lineages was not rejected by AU test (H05 in Table 6), we recognized each of them in its own tribe for the present (for Verulux , see the tribe Veruluxini ). The present molecular analyses did not include Bentuviaichthys , but it was included in this tribe based on the following morphological characters: fused hypurals 1+2+3+4, epipleurals absent on last three ribs, single row of teeth on dentary, two supraneurals, slender, mostly translucent body and forked caudal fin. Two individuals of Rhabdamia gracilis from two distantly distributed populations ( Fiji and Ryukyu Islands) were paraphyletic to R. spilota Allen & Kuiter 1994 and genetically distant from each other, which may indicate needs of taxonomical revisions of them.
Tribe Siphamiini Smith 1955
Type genus Siphamia Weber 1909
Diagnosis. Members of the Apogoninae : dorsal fin VI–VII + I,7–11; anal fin II,7–11; pored lateral-line scales 0–24; preopercular ridge smooth, edges smooth to serrate; supraneurals 1–2; supramaxilla absent; basisphenoid absent; uroneurals absent; two epurals; hypurals 1+2 and 3+4 fused into two plates, upper plate fused to terminal centrum; parhypural separate; one or two supernumerary dorsal spines; caudal fin emarginate or forked; bacteria bioluminescent system from hyal region to along body above or past anal-fin base.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal-fin ray branched and segmented; ctenoid, cycloid or spinoid scales; median predorsal scales 0–6; pectoral rays 11–16; segmented principal caudal rays 9+8, 15 branched, upper and lower unbranched; unbranched procurrent rays, longest segmented; villiform teeth in one or multiple rows on premaxilla, dentary, vomer, palatine, or absent on palatine; developed gill rakers 6–18; lateral-line scales usually with a vertical row of free neuromasts; six infraorbitals, bony shelf on third infraorbital; anterior ceratohyal smooth or notched; developed gill rakers 10–26; 10+14 vertebrae; 8 ribs; 8 epipleurals; stomach, intestine and peritoneum generally pale with variously sized melanophores; low crest on PU2.
Distribution. Representatives of the tribe are found from East Africa, Red Sea, islands in the Indian Ocean, throughout the West Pacific to Japan, onto the Pacific Plate to French Polynesia and Australia ( Gon & Allen 2012).
Remarks. This tribe contains one genus Siphamia , corresponding to the clade VII in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). All species of Siphamia have bioluminescent bacteria in a specialized organ (a synapomorphy for the species) unique among apogonines. Smith (1955) proposed placing species of Siphamia in its own subfamily Siphamiinae . Our results based on analysis of five of the twenty-three species suggest that Siphamia roseigaster Ramsay & Ogilby 1887 could be recognized in its own genus Adenapogon McCulloch 1921 , because it was placed relatively far from the remaining species. Gon & Allen's (2012) results based on morphology suggest that two other Australian species belong in Adenapogon and that Fodifoa Whitley 1936 is available for another group of species. We defer to Ofer Gon who is continuing to work on relationships within this tribe ( Gon & Allen 2012).
Tribe Sphaeramiini new name Fraser & Mabuchi
Type genus Sphaeramia Fowler & Bean 1930
Diagnosis. Members of the Apogoninae : dorsal fin VII+I,9–10, VII(I)+I,9–14 or VIII+I,9–10 with eighth spine reduced to a tiny visible spine or a nubbin hidden under skin supported by a free sixth distal radial; anal fin II,8–13; head and body with ctenoid scales; pored lateral-line scales 23–27; preopercle ridge smooth, edges serrate; 2–3 supraneurals; supramaxilla absent; basisphenoid present; one pair of uroneurals; three epurals; five free hypurals or 1+2 fused and 3+4 fused with 3+4 fused to terminal centrum; parhypural separate; two autogenous haemal spines; two supernumerary dorsal spines; caudal fin forked, emarginate or truncate.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal-fin ray branched and segmented; ctenoid scales on predorsal, cheek, breast, two pelvic scales, and body; ctenoid scales on opercle and onto base of caudal fin; 1–3 predorsal scales cycloid or ctenoid; pored lateral-line scales with multiple pores; pectoral-fin rays 13–17; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; developed gill rakers 7–38; unbranched procurrent rays, longest segmented or spinous; third or fourth dorsal spine longest; 10+14 or 10+15 vertebrae; 8 ribs; 8–9 epipleurals; teeth in one or multiple rows on premaxilla, dentary, vomer, palatine, all villiform; six infraorbitals, bony shelf on third infraorbital; stomach and intestine pale or black with pale peritoneum; low crest on PU2.
Distribution. Species of Apogonichthyoides are found throughout the Indo-West Pacific from East Africa, Red Sea, India, some island of the Indian Ocean, throughout Indonesia, Philippines to Japan, New Guinea, Australia and the Solomon Islands. Some species have restricted distribution and some are rare in collections. Species of Jaydia Smith 1961 have a more continental distribution, mostly caught in trawls, and are widespread from Africa to Japan, New Guinea, Australia and larger islands in the Coral Sea. Representatives of Nectamia are found on reef habitats from East Africa, Red Sea, islands in the Indian Ocean, throughout the West Pacific to Japan, onto the Pacific Plate out to the Line Islands and French Polynesia. Sphaeramia , represented by two species, one found in mangrove habitats and one on coral reefs, also have wide distribution in the Indo-West Pacific. Quinca , represented by a single species, is known from northwestern coast of Australia.
Remarks. This tribe contains six genera, Apogonichthyoides , Jaydia , Nectamia , Pterapogon Koumans 1933 , Quinca and Sphaeramia , almost corresponding to the clade IV in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). First three of the six genera were formerly classified under Ostorhinchus , corresponding to "barred group" of Ostorhinchus ( Ostorhinchus II ) in Mabuchi et al. (2006). Two of the remaining three genera ( Pterapogon and Sphaeramia ) have distinct bar(s) on body. In addition to the six genera, corresponding molecular clade included three species of Ostorhinchus -2 ( O. hoevenii Bleeker 1854b , O. ishigakiensis Ida & Moyer 1974 , and O. rueppellii Günther 1859 ). But there are no morphological characters that support this association. We defer taxonomical action on this possible relationship for the present. This clade further included another species of the former Ostorhinchus ( Ostorhinchus -1). For this species, the latest authors ( Allen & Erdmann 2012) provisionally used Apogonichthyoides as its genus name, and we followed it here, but such application of the name made this genus paraphyletic in our molecular trees. Based on its morphological features, we tentatively included it [ Apogonichthyoides (?) melas ] in this tribe, but further study including more Sphaeramiini species will be needed to clarify its taxonomic status. In the present molecular analyses, phylogenetic position of Pterapogon remained unsettled ( Table 5). It was sister to the clade VII ( Siphamiini ) in ML tree ( Fig. 2 View FIGURE 2 ), sister to Ostorhinchus margaritophorus in MP tree-A ( Fig. 4 View FIGURE 4 ), and nested within the clade XII ( Ostorhinchini ) in the MP tree-B ( Fig. 5 View FIGURE 5 ). It formed a polytomy with O. margaritophorus , clade VII, clade VIII, and a large clade including clades IX–XII in BA tree ( Fig. 3 View FIGURE 3 ). Pterapogon has, however, a synapomorphy (spinous procurrent caudal rays) uniquely held with Sphaeramia as well as similarities in color patterns and body shape, but differs with fused hypurals 1+2 and 3+4. Based on these morphological features, we tentatively included this species in the tribe Sphaeramiini . Monophyly of them was not rejected by AU test (H02 in Table 6), although its inclusion within Ostorhinchini was also not rejected (H03 in Table 6). We had no good DNA extracts from tissue of Quinca . This monotypic genus has morphological characteristics and color pattern suggestive of a relationship with Apogonichthyoides , and was provisionally place in the Sphaeramiini .
Tribe Veruluxini new name Fraser & Mabuchi
Type genus Verulux Fraser 1972
Diagnosis. Members of the Apogoninae : dorsal fin VI–I,9; anal fin II,9; head and body with weakly ctenoid or cycloid scales; pored lateral-line scales 24; preopercle ridge smooth, edges smooth; one supraneural; supramaxilla absent; basisphenoid present; uroneurals absent; two epurals; fused hypurals 1+2+3+4, fused to terminal centrum; no autogenous haemal spines; one supernumerary dorsal spine; bioluminescent organ under cleithrum; caudal fin forked.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal fin-ray branched and segmented; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; developed gill rakers 12–15; unbranched procurrent rays, longest segmented; pectoral fin-rays 14–16; 10+14 vertebrae; 8 ribs; 5–6 epipleurals; teeth in one row on premaxilla, dentary, vomer, palatine, all villiform, or absent on palatine; six infraorbitals, bony shelf absent on third infraorbital; stomach and intestine black with silvery peritoneum; low crest on PU2.
Distribution. The single described species is known from East Africa, Red Sea, islands in the Indian Ocean, throughout the West Pacific to Japan, onto the Pacific Plate to Marshall Islands, Vanuatu, New Caledonia and Australia.
Remarks. This tribe contains only one species, Verulux cypselurus , corresponding to the clade V in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). Representatives of Verulux (two individuals from Ryukyu Islands and Seychelles) were monophyletic separated by a moderate genetic distance, which may indicate presence of cryptic species. This lineage was recovered far from Rhabdamia (tribe Rhabdamiini ), although their monophyly (genus Rhabdamia sensu Fraser 1972 ) was not rejected by AU test (H05 in Table 6). The single described species has a bioluminescent organ under cleithrum unlike Rhabdamia .
Tribe Zoramiini new name Fraser & Mabuchi
Type genus Zoramia Jordan 1917
Diagnosis. Members of the Apogoninae : dorsal fin VI–I,9; anal fin II,8–9; head and body with ctenoid scales; pored lateral-line scales 24; preopercle ridge smooth, edges serrate; three supraneurals; supramaxilla absent; basisphenoid present; anterior pair of uroneurals; three epurals; five free hypurals; parhypural separate; two autogenous haemal spines; one supernumerary dorsal spine; caudal fin forked.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal-fin ray branched and segmented; second dorsal spine longest, very long and filamentous in some species of both genera; ctenoid scales on predorsal, cheek, breast, two pelvic scales, and body; ctenoid scales on opercle and onto base of caudal fin; cycloid or ctenoid predorsal scales1–3; pored lateral-line scales simple with one pore above and one below midline; pectoral-fin rays 13–17; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; developed gill rakers 7–38; unbranched procurrent rays, longest segmented; 10+14 vertebrae; 8 ribs; 7–8 epipleurals; teeth in one, two or multiple rows on premaxilla, dentary, vomer, palatine, all villiform; six infraorbitals, bony shelf absent or on third infraorbital; stomach and intestine black with silvery peritoneum; low crest on PU2.
Distribution. Species of Zoramia are known from East Africa, Red Sea, islands in the Indian Ocean, throughout the West Pacific to Japan, onto the Pacific Plate to Marshall Islands, Samoa, Tonga, Solomon Islands, New Caledonia and Australia. Species of Fibramia are known from East Africa, some islands in the Indian Ocean, Sri Lanka, throughout the West Pacific to Philippines, Solomon Islands, Santa Cruz Islands, Fiji, Vanuatu, New Caledonia, Australia, Samoa and Tonga.
Remarks. This tribe contains two genera, a new genus Fibramia described below and Zoramia , corresponding to the clade VIII in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). Both genera have species with elongate second dorsal spine, a single supernumerary dorsal spine, and can be found in tidal fresh and brackish water, near shore waters and in lagoons. Fibramia corresponds to the clade of Ostorhinchus - 4 in the present study, and that of Ostorhinchus I in Mabuchi et al. (2006).
Tribe Lepidamiini new name Fraser & Mabuchi
Type genus Lepidamia Gill 1863
Diagnosis. Members of the Apogoninae : dorsal fin VII(I)-I,9 or VIII-I,9; anal fin II,8; head and body with ctenoid scales; pored lateral-line scales 35–48; preopercle ridge smooth, edges serrate; three supraneurals; supramaxilla absent; basisphenoid present; one pair of reduced uroneurals present; three epurals; five free hypurals; parhypural separate; two autogenous haemal spines; two supernumerary dorsal spines, caudal fin forked.
Other characteristics. first fin ray in second dorsal fin branched and segmented; first anal-fin ray branched and segmented; ctenoid scales on predorsal, cheek, breast, four pelvic scales, and body; ctenoid scales on opercle and onto base of caudal fin; pored lateral-line scales with multiple pores; 15 branched, upper and lower unbranched; developed gill rakers 8–17; unbranched procurrent rays, longest segmented; 10+14 vertebrae; 8 ribs; 9 epipleurals; teeth in multiple rows on premaxilla, dentary, vomer, palatine, all villiform; six infraorbitals, bony shelf on third infraorbital; stomach and intestine pale, peritoneum pale; low crest on PU2.
Distribution. Lepidamia with four species has been collected only from continental marine waters from South Africa to China ( Gon 1995). Members of this tribe have not been collected in Australia or New Guinea. One specimen (USNM 175754, 96 mm SL) was collected in 1909 from Manila Bay, Luzon, Philippines. None have been reported elsewhere in the Philippines or again from Manila Bay. Perhaps the location is erroneous.
Remarks. We did not have tissue from any species. Based on known morphological characters including color patterns of the juveniles and adults, body shape and small body scales, we recognized Lepidamia in its own tribe for the present.
Tribe Ostorhinchini Whitley 1959
Type genus Ostorhinchus Lacepède 1802
Diagnosis. Members of the Apogoninae : dorsal fin VI+I,9 or VII+I,9; anal fin II,8–9; head and body with ctenoid scales; pored lateral-line scales 6–26; preopercle ridge smooth, edges serrate; 2–3 supraneurals; supramaxilla absent; basisphenoid present; one pair of reduced uroneurals present or absent; three epurals; five free hypurals or 1–2 fused; parhypural separate; caudal fin forked; head and/or body with one or more light or dark stripes, bars rarely present, basicaudal spot or broad mark present or absent.
Other characteristics. one or two supernumerary dorsal spines; branched first segmented fin ray in second dorsal-fin; first anal-fin ray branched and segmented; ctenoid scales on predorsal, cheek, breast, two pelvic scales, and body; ctenoid scale on opercle and onto base of caudal fin; pored lateral-line scales with multiple pores; pectoral fin-rays 11–16; three supraneurals; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; unbranched procurrent rays, longest segmented; teeth in one or multiple rows on premaxilla, dentary, vomer, palatine, all villiform or absent on palatine, sometimes enlarged, but not canine on side of dentary; six infraorbitals, bony shelf on third infraorbital; anterior ceratohyal smooth or notched; developed gill rakers 10–26; 10+14 vertebrae; 8 ribs; 8 epipleurals; blackish stomachs and intestines; low crest on PU2.
Distribution. The restricted Ostorhinchus is widespread throughout the Indo-Pacific from the Red Sea, East Africa to Japan, Hawaii, Easter Island and French Polynesia.
Remarks. Monophyly of recent Ostorhinchus (species indicated by blue in Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 ) was significantly rejected by AU test (H 16 in Table 6). This tribe, however, corresponds to one of the five lineages of the recent Ostorhinchus , clade XII. It included most species of the recent Ostorhinchus : many striped species of Ostorhinchus (species of Ostorhinchus -5) and Brephamia parvula Smith & Radcliffe in Radcliffe 1912 ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). Other species of the recent Ostorhinchus were divided into four lineages, Ostorhinchus -1, 2, 3 and 4. They were all placed far from the clade XII. Except for Ostorhinchus -3 ( O. margaritophorus ), monophyly between this tribe (clade XII) and each of the remaining lineages was rejected by AU tests (H07, 08 and 11 in Table 6). Although the bootstrap values supporting the clade XII were not so high (<50%) both in the ML ( Fig. 2 View FIGURE 2 ) and MP ( Figs. 4 View FIGURE 4 and 5 View FIGURE 5 ) trees ( Table 4), and further the clade was divided into two separate clades in the BA tree ( Fig. 3 View FIGURE 3 ), we are treating this group as a single genus, based on the typical color pattern (stripes on body and/or head) shared by most of the included species. This group corresponds to the "striped group" of Ostorhinchus ( Ostorhinchus III ) in Mabuchi et al. (2006). One of the other recent Ostorhinchus lineages, O. margaritophorus , has long stripes, short bars between two long stripes on body and fused hypurals 1+2. Phylogenetic position of this species remains unsettled in the molecular trees ( Table 5), but monophyly between this species and the members of this tribe was not rejected by AU test (H01 in Table 6). Based on its featured color pattern, we tentatively included this species in this tribe. Apogon ( Brephamia) parvula has a very similar species within this tribe. It is Ostorhinchus neotes . Although they were not sister to each other in the molecular trees, their monophyly was not rejected by AU test (H 12 in Table 6). While recognition of Brephamia Jordan in Jordan & Jordan (1922) as a subgenus or genus needs further evaluation, we synonymized it under the restricted Ostorhinchus for the present. Likely synapomorphies may include color pattern groupings, probably at the subgenus level. At the genus level, with the exception of the loss of tiny first dorsal-fin spines, all species have 7 visible first dorsal spines, a serrated preopercular edge and most have blackish stomachs and intestines.
There are two available names ( Gronovichthys Whitley 1929 ; Lovamia Whitley 1930 ) which could be used in the future. There are at least 93 species in the group. We have sampled 31 species. Whitley (1959) first used the present name at family level Ostorhinchidae. The name appeared once. We use the name at the tribal level.
Tribe Pristiapogonini new name Fraser & Mabuchi
Type genus Pristiapogon Klunzinger 1870
Diagnosis. Members of the Apogoninae : dorsal fin VI+I,9 or VII+I,9; anal fin II,8; head and body with ctenoid scales; pored lateral-line scales 23–25; preopercle ridge serrate, edges serrate; infraorbitals serrate; three supraneurals; supramaxilla absent; basisphenoid present; one pair of uroneurals or absent; three epurals; five free hypurals; parhypural separate; one or two supernumerary dorsal spines; caudal fin forked.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal-fin ray branched and segmented; ctenoid scales on predorsal, cheek, breast, two pelvic scales, and body; ctenoid scales on opercle and onto base of caudal fin; pored lateral-line scales with multiple pores; pectoral fin-rays 12–16; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; unbranched procurrent rays, longest segmented; teeth in one or multiple rows on premaxilla, dentary, vomer, palatine, all villiform or absent on palatine; six infraorbitals, bony shelf on third infraorbital; anterior ceratohyal smooth or notched; developed gill rakers 10–19; 10+14 vertebrae; 8 ribs; 8 epipleurals; low crest on PU2.
Distribution. Pristiapogon is wide spread from East Africa, Red Sea, Indian Ocean Islands, Indonesia, Philippines, Pacific islands to Hawaii, Japan, French Polynesia and Australia. One species of Pristicon Fraser 1972 is restricted to the West Pacific. Another species reaches out to some islands on the Pacific Plate. One species of Pristicon has been reported from the west coast of India, with a gap between India and Indonesia ( Suresh & Thomas 2007).
Remarks. This tribe contains two genera, Pristiapogon and Pristicon , corresponding to the clade VI in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). Although bootstrap values supporting the tribe were not so high (62 and 56% BPs in ML and MP analyses, respectively), this tribe is morphologically well-defined: relatively large body with serrations on the preopercle ridge (a likely synapomorphy), edges and infraorbitals (a likely synapomorphy). Species of Pristiapogon usually have a darkish single stripe and/or a variable basicaudal spot while Pristicon have bars or saddles under the dorsal fins and spots at the base of the caudal fin or on the opercle.
Tribe Rhabdamiini new name Fraser & Mabuchi
Type genus Rhabdamia Weber 1909
Diagnosis. Members of the Apogoninae : dorsal fin VI+I,9 or VII+I,10–11; anal fin II,11–13; head and body with weakly ctenoid or cycloid; pored lateral-line scales 23–24; preopercle ridge smooth, edges smooth; two supraneurals; supramaxilla absent; basisphenoid present or absent; anterior pair of uroneurals reduced or absent; three epurals; hypurals 1+2+3+4 fused, the plate fused to terminal centrum; parhypural separate; no autogenous haemal spines; 1–2 supernumerary dorsal spines; caudal fin forked.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal-fin ray branched and segmented; pectoral-fin rays 13–17; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; developed gill rakers 17–31; unbranched procurrent rays, longest segmented; villiform teeth in one or two rows on premaxilla, or with a few canines ( Bentuviaichthys ), one villiform row on dentary and vomer, one villiform row or absent on palatine; six infraorbitals, bony shelf reduce or absent on third infraorbital; stomach and intestine pale with melanophores and silvery peritoneum with melanophores; 10+14 vertebrae; low crest on PU2.
Distribution. No reviews of the species have been made, but members have been reported from East Africa, the Red Sea, islands in the Indian Ocean, the West Pacific and out onto the Pacific Plate.
Remarks. This tribe contains one genus Rhabdamia (including two subgenera Rhabdamia and Bentuviaichthys sensu Fraser 1972 ). There are six names (Appendix A). Three species are streamlined apogonines exploiting the near pelagic zone of coral reefs for food. Likely synapomorphies include smooth preopercle ridge and edges, reduced dentition, fused hypurals, and two supraneurals. All species are mostly translucent. In the present molecular analyses, they formed a robust monophyletic group (clade IX) with Yarica hyalosoma ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4) (see also Fig. 7 View FIGURE 7 ), but it is very unlikely based on morphological evidence (for details, see the remarks of the tribe Glossamiini ). In Fraser (1972), subgenus Rhabdamia was classified under the genus Rhabdamia together with the subgenera Bentuviaichthys and Verulux Fraser 1972 . The last lineage, Verulux (clade V), was placed far from the first lineage, Rhabdamia , in the present molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 ). Although monophyly between the two lineages was not rejected by AU test (H05 in Table 6), we recognized each of them in its own tribe for the present (for Verulux , see the tribe Veruluxini ). The present molecular analyses did not include Bentuviaichthys , but it was included in this tribe based on the following morphological characters: fused hypurals 1+2+3+4, epipleurals absent on last three ribs, single row of teeth on dentary, two supraneurals, slender, mostly translucent body and forked caudal fin. Two individuals of Rhabdamia gracilis from two distantly distributed populations ( Fiji and Ryukyu Islands) were paraphyletic to R. spilota Allen & Kuiter 1994 and genetically distant from each other, which may indicate needs of taxonomical revisions of them.
Tribe Siphamiini Smith 1955
Type genus Siphamia Weber 1909
Diagnosis. Members of the Apogoninae : dorsal fin VI–VII + I,7–11; anal fin II,7–11; pored lateral-line scales 0–24; preopercular ridge smooth, edges smooth to serrate; supraneurals 1–2; supramaxilla absent; basisphenoid absent; uroneurals absent; two epurals; hypurals 1+2 and 3+4 fused into two plates, upper plate fused to terminal centrum; parhypural separate; one or two supernumerary dorsal spines; caudal fin emarginate or forked; bacteria bioluminescent system from hyal region to along body above or past anal-fin base.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal-fin ray branched and segmented; ctenoid, cycloid or spinoid scales; median predorsal scales 0–6; pectoral rays 11–16; segmented principal caudal rays 9+8, 15 branched, upper and lower unbranched; unbranched procurrent rays, longest segmented; villiform teeth in one or multiple rows on premaxilla, dentary, vomer, palatine, or absent on palatine; developed gill rakers 6–18; lateral-line scales usually with a vertical row of free neuromasts; six infraorbitals, bony shelf on third infraorbital; anterior ceratohyal smooth or notched; developed gill rakers 10–26; 10+14 vertebrae; 8 ribs; 8 epipleurals; stomach, intestine and peritoneum generally pale with variously sized melanophores; low crest on PU2.
Distribution. Representatives of the tribe are found from East Africa, Red Sea, islands in the Indian Ocean, throughout the West Pacific to Japan, onto the Pacific Plate to French Polynesia and Australia ( Gon & Allen 2012).
Remarks. This tribe contains one genus Siphamia , corresponding to the clade VII in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). All species of Siphamia have bioluminescent bacteria in a specialized organ (a synapomorphy for the species) unique among apogonines. Smith (1955) proposed placing species of Siphamia in its own subfamily Siphamiinae . Our results based on analysis of five of the twenty-three species suggest that Siphamia roseigaster Ramsay & Ogilby 1887 could be recognized in its own genus Adenapogon McCulloch 1921 , because it was placed relatively far from the remaining species. Gon & Allen's (2012) results based on morphology suggest that two other Australian species belong in Adenapogon and that Fodifoa Whitley 1936 is available for another group of species. We defer to Ofer Gon who is continuing to work on relationships within this tribe ( Gon & Allen 2012).
Tribe Sphaeramiini new name Fraser & Mabuchi
Type genus Sphaeramia Fowler & Bean 1930
Diagnosis. Members of the Apogoninae : dorsal fin VII+I,9–10, VII(I)+I,9–14 or VIII+I,9–10 with eighth spine reduced to a tiny visible spine or a nubbin hidden under skin supported by a free sixth distal radial; anal fin II,8–13; head and body with ctenoid scales; pored lateral-line scales 23–27; preopercle ridge smooth, edges serrate; 2–3 supraneurals; supramaxilla absent; basisphenoid present; one pair of uroneurals; three epurals; five free hypurals or 1+2 fused and 3+4 fused with 3+4 fused to terminal centrum; parhypural separate; two autogenous haemal spines; two supernumerary dorsal spines; caudal fin forked, emarginate or truncate.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal-fin ray branched and segmented; ctenoid scales on predorsal, cheek, breast, two pelvic scales, and body; ctenoid scales on opercle and onto base of caudal fin; 1–3 predorsal scales cycloid or ctenoid; pored lateral-line scales with multiple pores; pectoral-fin rays 13–17; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; developed gill rakers 7–38; unbranched procurrent rays, longest segmented or spinous; third or fourth dorsal spine longest; 10+14 or 10+15 vertebrae; 8 ribs; 8–9 epipleurals; teeth in one or multiple rows on premaxilla, dentary, vomer, palatine, all villiform; six infraorbitals, bony shelf on third infraorbital; stomach and intestine pale or black with pale peritoneum; low crest on PU2.
Distribution. Species of Apogonichthyoides are found throughout the Indo-West Pacific from East Africa, Red Sea, India, some island of the Indian Ocean, throughout Indonesia, Philippines to Japan, New Guinea, Australia and the Solomon Islands. Some species have restricted distribution and some are rare in collections. Species of Jaydia Smith 1961 have a more continental distribution, mostly caught in trawls, and are widespread from Africa to Japan, New Guinea, Australia and larger islands in the Coral Sea. Representatives of Nectamia are found on reef habitats from East Africa, Red Sea, islands in the Indian Ocean, throughout the West Pacific to Japan, onto the Pacific Plate out to the Line Islands and French Polynesia. Sphaeramia , represented by two species, one found in mangrove habitats and one on coral reefs, also have wide distribution in the Indo-West Pacific. Quinca , represented by a single species, is known from northwestern coast of Australia.
Remarks. This tribe contains six genera, Apogonichthyoides , Jaydia , Nectamia , Pterapogon Koumans 1933 , Quinca and Sphaeramia , almost corresponding to the clade IV in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). First three of the six genera were formerly classified under Ostorhinchus , corresponding to "barred group" of Ostorhinchus ( Ostorhinchus II ) in Mabuchi et al. (2006). Two of the remaining three genera ( Pterapogon and Sphaeramia ) have distinct bar(s) on body. In addition to the six genera, corresponding molecular clade included three species of Ostorhinchus -2 ( O. hoevenii Bleeker 1854b , O. ishigakiensis Ida & Moyer 1974 , and O. rueppellii Günther 1859 ). But there are no morphological characters that support this association. We defer taxonomical action on this possible relationship for the present. This clade further included another species of the former Ostorhinchus ( Ostorhinchus -1). For this species, the latest authors ( Allen & Erdmann 2012) provisionally used Apogonichthyoides as its genus name, and we followed it here, but such application of the name made this genus paraphyletic in our molecular trees. Based on its morphological features, we tentatively included it [ Apogonichthyoides (?) melas ] in this tribe, but further study including more Sphaeramiini species will be needed to clarify its taxonomic status. In the present molecular analyses, phylogenetic position of Pterapogon remained unsettled ( Table 5). It was sister to the clade VII ( Siphamiini ) in ML tree ( Fig. 2 View FIGURE 2 ), sister to Ostorhinchus margaritophorus in MP tree-A ( Fig. 4 View FIGURE 4 ), and nested within the clade XII ( Ostorhinchini ) in the MP tree-B ( Fig. 5 View FIGURE 5 ). It formed a polytomy with O. margaritophorus , clade VII, clade VIII, and a large clade including clades IX–XII in BA tree ( Fig. 3 View FIGURE 3 ). Pterapogon has, however, a synapomorphy (spinous procurrent caudal rays) uniquely held with Sphaeramia as well as similarities in color patterns and body shape, but differs with fused hypurals 1+2 and 3+4. Based on these morphological features, we tentatively included this species in the tribe Sphaeramiini . Monophyly of them was not rejected by AU test (H02 in Table 6), although its inclusion within Ostorhinchini was also not rejected (H03 in Table 6). We had no good DNA extracts from tissue of Quinca . This monotypic genus has morphological characteristics and color pattern suggestive of a relationship with Apogonichthyoides , and was provisionally place in the Sphaeramiini .
Tribe Veruluxini new name Fraser & Mabuchi
Type genus Verulux Fraser 1972
Diagnosis. Members of the Apogoninae : dorsal fin VI–I,9; anal fin II,9; head and body with weakly ctenoid or cycloid scales; pored lateral-line scales 24; preopercle ridge smooth, edges smooth; one supraneural; supramaxilla absent; basisphenoid present; uroneurals absent; two epurals; fused hypurals 1+2+3+4, fused to terminal centrum; no autogenous haemal spines; one supernumerary dorsal spine; bioluminescent organ under cleithrum; caudal fin forked.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal fin-ray branched and segmented; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; developed gill rakers 12–15; unbranched procurrent rays, longest segmented; pectoral fin-rays 14–16; 10+14 vertebrae; 8 ribs; 5–6 epipleurals; teeth in one row on premaxilla, dentary, vomer, palatine, all villiform, or absent on palatine; six infraorbitals, bony shelf absent on third infraorbital; stomach and intestine black with silvery peritoneum; low crest on PU2.
Distribution. The single described species is known from East Africa, Red Sea, islands in the Indian Ocean, throughout the West Pacific to Japan, onto the Pacific Plate to Marshall Islands, Vanuatu, New Caledonia and Australia.
Remarks. This tribe contains only one species, Verulux cypselurus , corresponding to the clade V in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). Representatives of Verulux (two individuals from Ryukyu Islands and Seychelles) were monophyletic separated by a moderate genetic distance, which may indicate presence of cryptic species. This lineage was recovered far from Rhabdamia (tribe Rhabdamiini ), although their monophyly (genus Rhabdamia sensu Fraser 1972 ) was not rejected by AU test (H05 in Table 6). The single described species has a bioluminescent organ under cleithrum unlike Rhabdamia .
Tribe Zoramiini new name Fraser & Mabuchi
Type genus Zoramia Jordan 1917
Diagnosis. Members of the Apogoninae : dorsal fin VI–I,9; anal fin II,8–9; head and body with ctenoid scales; pored lateral-line scales 24; preopercle ridge smooth, edges serrate; three supraneurals; supramaxilla absent; basisphenoid present; anterior pair of uroneurals; three epurals; five free hypurals; parhypural separate; two autogenous haemal spines; one supernumerary dorsal spine; caudal fin forked.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal-fin ray branched and segmented; second dorsal spine longest, very long and filamentous in some species of both genera; ctenoid scales on predorsal, cheek, breast, two pelvic scales, and body; ctenoid scales on opercle and onto base of caudal fin; cycloid or ctenoid predorsal scales1–3; pored lateral-line scales simple with one pore above and one below midline; pectoral-fin rays 13–17; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; developed gill rakers 7–38; unbranched procurrent rays, longest segmented; 10+14 vertebrae; 8 ribs; 7–8 epipleurals; teeth in one, two or multiple rows on premaxilla, dentary, vomer, palatine, all villiform; six infraorbitals, bony shelf absent or on third infraorbital; stomach and intestine black with silvery peritoneum; low crest on PU2.
Distribution. Species of Zoramia are known from East Africa, Red Sea, islands in the Indian Ocean, throughout the West Pacific to Japan, onto the Pacific Plate to Marshall Islands, Samoa, Tonga, Solomon Islands, New Caledonia and Australia. Species of Fibramia are known from East Africa, some islands in the Indian Ocean, Sri Lanka, throughout the West Pacific to Philippines, Solomon Islands, Santa Cruz Islands, Fiji, Vanuatu, New Caledonia, Australia, Samoa and Tonga.
Remarks. This tribe contains two genera, a new genus Fibramia described below and Zoramia , corresponding to the clade VIII in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). Both genera have species with elongate second dorsal spine, a single supernumerary dorsal spine, and can be found in tidal fresh and brackish water, near shore waters and in lagoons. Fibramia corresponds to the clade of Ostorhinchus - 4 in the present study, and that of Ostorhinchus I in Mabuchi et al. (2006).
Tribe Ostorhinchini Whitley 1959
Type genus Ostorhinchus Lacepède 1802
Diagnosis. Members of the Apogoninae : dorsal fin VI+I,9 or VII+I,9; anal fin II,8–9; head and body with ctenoid scales; pored lateral-line scales 6–26; preopercle ridge smooth, edges serrate; 2–3 supraneurals; supramaxilla absent; basisphenoid present; one pair of reduced uroneurals present or absent; three epurals; five free hypurals or 1–2 fused; parhypural separate; caudal fin forked; head and/or body with one or more light or dark stripes, bars rarely present, basicaudal spot or broad mark present or absent.
Other characteristics. one or two supernumerary dorsal spines; branched first segmented fin ray in second dorsal-fin; first anal-fin ray branched and segmented; ctenoid scales on predorsal, cheek, breast, two pelvic scales, and body; ctenoid scale on opercle and onto base of caudal fin; pored lateral-line scales with multiple pores; pectoral fin-rays 11–16; three supraneurals; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; unbranched procurrent rays, longest segmented; teeth in one or multiple rows on premaxilla, dentary, vomer, palatine, all villiform or absent on palatine, sometimes enlarged, but not canine on side of dentary; six infraorbitals, bony shelf on third infraorbital; anterior ceratohyal smooth or notched; developed gill rakers 10–26; 10+14 vertebrae; 8 ribs; 8 epipleurals; blackish stomachs and intestines; low crest on PU2.
Distribution. The restricted Ostorhinchus is widespread throughout the Indo-Pacific from the Red Sea, East Africa to Japan, Hawaii, Easter Island and French Polynesia.
Remarks. Monophyly of recent Ostorhinchus (species indicated by blue in Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 ) was significantly rejected by AU test (H 16 in Table 6). This tribe, however, corresponds to one of the five lineages of the recent Ostorhinchus , clade XII. It included most species of the recent Ostorhinchus : many striped species of Ostorhinchus (species of Ostorhinchus -5) and Brephamia parvula Smith & Radcliffe in Radcliffe 1912 ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). Other species of the recent Ostorhinchus were divided into four lineages, Ostorhinchus -1, 2, 3 and 4. They were all placed far from the clade XII. Except for Ostorhinchus -3 ( O. margaritophorus ), monophyly between this tribe (clade XII) and each of the remaining lineages was rejected by AU tests (H07, 08 and 11 in Table 6). Although the bootstrap values supporting the clade XII were not so high (<50%) both in the ML ( Fig. 2 View FIGURE 2 ) and MP ( Figs. 4 View FIGURE 4 and 5 View FIGURE 5 ) trees ( Table 4), and further the clade was divided into two separate clades in the BA tree ( Fig. 3 View FIGURE 3 ), we are treating this group as a single genus, based on the typical color pattern (stripes on body and/or head) shared by most of the included species. This group corresponds to the "striped group" of Ostorhinchus ( Ostorhinchus III ) in Mabuchi et al. (2006). One of the other recent Ostorhinchus lineages, O. margaritophorus , has long stripes, short bars between two long stripes on body and fused hypurals 1+2. Phylogenetic position of this species remains unsettled in the molecular trees ( Table 5), but monophyly between this species and the members of this tribe was not rejected by AU test (H01 in Table 6). Based on its featured color pattern, we tentatively included this species in this tribe. Apogon ( Brephamia) parvula has a very similar species within this tribe. It is Ostorhinchus neotes . Although they were not sister to each other in the molecular trees, their monophyly was not rejected by AU test (H 12 in Table 6). While recognition of Brephamia Jordan in Jordan & Jordan (1922) as a subgenus or genus needs further evaluation, we synonymized it under the restricted Ostorhinchus for the present. Likely synapomorphies may include color pattern groupings, probably at the subgenus level. At the genus level, with the exception of the loss of tiny first dorsal-fin spines, all species have 7 visible first dorsal spines, a serrated preopercular edge and most have blackish stomachs and intestines.
There are two available names ( Gronovichthys Whitley 1929 ; Lovamia Whitley 1930 ) which could be used in the future. There are at least 93 species in the group. We have sampled 31 species. Whitley (1959) first used the present name at family level Ostorhinchidae. The name appeared once. We use the name at the tribal level.
Tribe Pristiapogonini new name Fraser & Mabuchi
Type genus Pristiapogon Klunzinger 1870
Diagnosis. Members of the Apogoninae : dorsal fin VI+I,9 or VII+I,9; anal fin II,8; head and body with ctenoid scales; pored lateral-line scales 23–25; preopercle ridge serrate, edges serrate; infraorbitals serrate; three supraneurals; supramaxilla absent; basisphenoid present; one pair of uroneurals or absent; three epurals; five free hypurals; parhypural separate; one or two supernumerary dorsal spines; caudal fin forked.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal-fin ray branched and segmented; ctenoid scales on predorsal, cheek, breast, two pelvic scales, and body; ctenoid scales on opercle and onto base of caudal fin; pored lateral-line scales with multiple pores; pectoral fin-rays 12–16; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; unbranched procurrent rays, longest segmented; teeth in one or multiple rows on premaxilla, dentary, vomer, palatine, all villiform or absent on palatine; six infraorbitals, bony shelf on third infraorbital; anterior ceratohyal smooth or notched; developed gill rakers 10–19; 10+14 vertebrae; 8 ribs; 8 epipleurals; low crest on PU2.
Distribution. Pristiapogon is wide spread from East Africa, Red Sea, Indian Ocean Islands, Indonesia, Philippines, Pacific islands to Hawaii, Japan, French Polynesia and Australia. One species of Pristicon Fraser 1972 is restricted to the West Pacific. Another species reaches out to some islands on the Pacific Plate. One species of Pristicon has been reported from the west coast of India, with a gap between India and Indonesia ( Suresh & Thomas 2007).
Remarks. This tribe contains two genera, Pristiapogon and Pristicon , corresponding to the clade VI in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). Although bootstrap values supporting the tribe were not so high (62 and 56% BPs in ML and MP analyses, respectively), this tribe is morphologically well-defined: relatively large body with serrations on the preopercle ridge (a likely synapomorphy), edges and infraorbitals (a likely synapomorphy). Species of Pristiapogon usually have a darkish single stripe and/or a variable basicaudal spot while Pristicon have bars or saddles under the dorsal fins and spots at the base of the caudal fin or on the opercle.
Tribe Rhabdamiini new name Fraser & Mabuchi
Type genus Rhabdamia Weber 1909
Diagnosis. Members of the Apogoninae : dorsal fin VI+I,9 or VII+I,10–11; anal fin II,11–13; head and body with weakly ctenoid or cycloid; pored lateral-line scales 23–24; preopercle ridge smooth, edges smooth; two supraneurals; supramaxilla absent; basisphenoid present or absent; anterior pair of uroneurals reduced or absent; three epurals; hypurals 1+2+3+4 fused, the plate fused to terminal centrum; parhypural separate; no autogenous haemal spines; 1–2 supernumerary dorsal spines; caudal fin forked.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal-fin ray branched and segmented; pectoral-fin rays 13–17; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; developed gill rakers 17–31; unbranched procurrent rays, longest segmented; villiform teeth in one or two rows on premaxilla, or with a few canines ( Bentuviaichthys ), one villiform row on dentary and vomer, one villiform row or absent on palatine; six infraorbitals, bony shelf reduce or absent on third infraorbital; stomach and intestine pale with melanophores and silvery peritoneum with melanophores; 10+14 vertebrae; low crest on PU2.
Distribution. No reviews of the species have been made, but members have been reported from East Africa, the Red Sea, islands in the Indian Ocean, the West Pacific and out onto the Pacific Plate.
Remarks. This tribe contains one genus Rhabdamia (including two subgenera Rhabdamia and Bentuviaichthys sensu Fraser 1972 ). There are six names (Appendix A). Three species are streamlined apogonines exploiting the near pelagic zone of coral reefs for food. Likely synapomorphies include smooth preopercle ridge and edges, reduced dentition, fused hypurals, and two supraneurals. All species are mostly translucent. In the present molecular analyses, they formed a robust monophyletic group (clade IX) with Yarica hyalosoma ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4) (see also Fig. 7 View FIGURE 7 ), but it is very unlikely based on morphological evidence (for details, see the remarks of the tribe Glossamiini ). In Fraser (1972), subgenus Rhabdamia was classified under the genus Rhabdamia together with the subgenera Bentuviaichthys and Verulux Fraser 1972 . The last lineage, Verulux (clade V), was placed far from the first lineage, Rhabdamia , in the present molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 ). Although monophyly between the two lineages was not rejected by AU test (H05 in Table 6), we recognized each of them in its own tribe for the present (for Verulux , see the tribe Veruluxini ). The present molecular analyses did not include Bentuviaichthys , but it was included in this tribe based on the following morphological characters: fused hypurals 1+2+3+4, epipleurals absent on last three ribs, single row of teeth on dentary, two supraneurals, slender, mostly translucent body and forked caudal fin. Two individuals of Rhabdamia gracilis from two distantly distributed populations ( Fiji and Ryukyu Islands) were paraphyletic to R. spilota Allen & Kuiter 1994 and genetically distant from each other, which may indicate needs of taxonomical revisions of them.
Tribe Siphamiini Smith 1955
Type genus Siphamia Weber 1909
Diagnosis. Members of the Apogoninae : dorsal fin VI–VII + I,7–11; anal fin II,7–11; pored lateral-line scales 0–24; preopercular ridge smooth, edges smooth to serrate; supraneurals 1–2; supramaxilla absent; basisphenoid absent; uroneurals absent; two epurals; hypurals 1+2 and 3+4 fused into two plates, upper plate fused to terminal centrum; parhypural separate; one or two supernumerary dorsal spines; caudal fin emarginate or forked; bacteria bioluminescent system from hyal region to along body above or past anal-fin base.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal-fin ray branched and segmented; ctenoid, cycloid or spinoid scales; median predorsal scales 0–6; pectoral rays 11–16; segmented principal caudal rays 9+8, 15 branched, upper and lower unbranched; unbranched procurrent rays, longest segmented; villiform teeth in one or multiple rows on premaxilla, dentary, vomer, palatine, or absent on palatine; developed gill rakers 6–18; lateral-line scales usually with a vertical row of free neuromasts; six infraorbitals, bony shelf on third infraorbital; anterior ceratohyal smooth or notched; developed gill rakers 10–26; 10+14 vertebrae; 8 ribs; 8 epipleurals; stomach, intestine and peritoneum generally pale with variously sized melanophores; low crest on PU2.
Distribution. Representatives of the tribe are found from East Africa, Red Sea, islands in the Indian Ocean, throughout the West Pacific to Japan, onto the Pacific Plate to French Polynesia and Australia ( Gon & Allen 2012).
Remarks. This tribe contains one genus Siphamia , corresponding to the clade VII in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). All species of Siphamia have bioluminescent bacteria in a specialized organ (a synapomorphy for the species) unique among apogonines. Smith (1955) proposed placing species of Siphamia in its own subfamily Siphamiinae . Our results based on analysis of five of the twenty-three species suggest that Siphamia roseigaster Ramsay & Ogilby 1887 could be recognized in its own genus Adenapogon McCulloch 1921 , because it was placed relatively far from the remaining species. Gon & Allen's (2012) results based on morphology suggest that two other Australian species belong in Adenapogon and that Fodifoa Whitley 1936 is available for another group of species. We defer to Ofer Gon who is continuing to work on relationships within this tribe ( Gon & Allen 2012).
Tribe Sphaeramiini new name Fraser & Mabuchi
Type genus Sphaeramia Fowler & Bean 1930
Diagnosis. Members of the Apogoninae : dorsal fin VII+I,9–10, VII(I)+I,9–14 or VIII+I,9–10 with eighth spine reduced to a tiny visible spine or a nubbin hidden under skin supported by a free sixth distal radial; anal fin II,8–13; head and body with ctenoid scales; pored lateral-line scales 23–27; preopercle ridge smooth, edges serrate; 2–3 supraneurals; supramaxilla absent; basisphenoid present; one pair of uroneurals; three epurals; five free hypurals or 1+2 fused and 3+4 fused with 3+4 fused to terminal centrum; parhypural separate; two autogenous haemal spines; two supernumerary dorsal spines; caudal fin forked, emarginate or truncate.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal-fin ray branched and segmented; ctenoid scales on predorsal, cheek, breast, two pelvic scales, and body; ctenoid scales on opercle and onto base of caudal fin; 1–3 predorsal scales cycloid or ctenoid; pored lateral-line scales with multiple pores; pectoral-fin rays 13–17; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; developed gill rakers 7–38; unbranched procurrent rays, longest segmented or spinous; third or fourth dorsal spine longest; 10+14 or 10+15 vertebrae; 8 ribs; 8–9 epipleurals; teeth in one or multiple rows on premaxilla, dentary, vomer, palatine, all villiform; six infraorbitals, bony shelf on third infraorbital; stomach and intestine pale or black with pale peritoneum; low crest on PU2.
Distribution. Species of Apogonichthyoides are found throughout the Indo-West Pacific from East Africa, Red Sea, India, some island of the Indian Ocean, throughout Indonesia, Philippines to Japan, New Guinea, Australia and the Solomon Islands. Some species have restricted distribution and some are rare in collections. Species of Jaydia Smith 1961 have a more continental distribution, mostly caught in trawls, and are widespread from Africa to Japan, New Guinea, Australia and larger islands in the Coral Sea. Representatives of Nectamia are found on reef habitats from East Africa, Red Sea, islands in the Indian Ocean, throughout the West Pacific to Japan, onto the Pacific Plate out to the Line Islands and French Polynesia. Sphaeramia , represented by two species, one found in mangrove habitats and one on coral reefs, also have wide distribution in the Indo-West Pacific. Quinca , represented by a single species, is known from northwestern coast of Australia.
Remarks. This tribe contains six genera, Apogonichthyoides , Jaydia , Nectamia , Pterapogon Koumans 1933 , Quinca and Sphaeramia , almost corresponding to the clade IV in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). First three of the six genera were formerly classified under Ostorhinchus , corresponding to "barred group" of Ostorhinchus ( Ostorhinchus II ) in Mabuchi et al. (2006). Two of the remaining three genera ( Pterapogon and Sphaeramia ) have distinct bar(s) on body. In addition to the six genera, corresponding molecular clade included three species of Ostorhinchus -2 ( O. hoevenii Bleeker 1854b , O. ishigakiensis Ida & Moyer 1974 , and O. rueppellii Günther 1859 ). But there are no morphological characters that support this association. We defer taxonomical action on this possible relationship for the present. This clade further included another species of the former Ostorhinchus ( Ostorhinchus -1). For this species, the latest authors ( Allen & Erdmann 2012) provisionally used Apogonichthyoides as its genus name, and we followed it here, but such application of the name made this genus paraphyletic in our molecular trees. Based on its morphological features, we tentatively included it [ Apogonichthyoides (?) melas ] in this tribe, but further study including more Sphaeramiini species will be needed to clarify its taxonomic status. In the present molecular analyses, phylogenetic position of Pterapogon remained unsettled ( Table 5). It was sister to the clade VII ( Siphamiini ) in ML tree ( Fig. 2 View FIGURE 2 ), sister to Ostorhinchus margaritophorus in MP tree-A ( Fig. 4 View FIGURE 4 ), and nested within the clade XII ( Ostorhinchini ) in the MP tree-B ( Fig. 5 View FIGURE 5 ). It formed a polytomy with O. margaritophorus , clade VII, clade VIII, and a large clade including clades IX–XII in BA tree ( Fig. 3 View FIGURE 3 ). Pterapogon has, however, a synapomorphy (spinous procurrent caudal rays) uniquely held with Sphaeramia as well as similarities in color patterns and body shape, but differs with fused hypurals 1+2 and 3+4. Based on these morphological features, we tentatively included this species in the tribe Sphaeramiini . Monophyly of them was not rejected by AU test (H02 in Table 6), although its inclusion within Ostorhinchini was also not rejected (H03 in Table 6). We had no good DNA extracts from tissue of Quinca . This monotypic genus has morphological characteristics and color pattern suggestive of a relationship with Apogonichthyoides , and was provisionally place in the Sphaeramiini .
Tribe Veruluxini new name Fraser & Mabuchi
Type genus Verulux Fraser 1972
Diagnosis. Members of the Apogoninae : dorsal fin VI–I,9; anal fin II,9; head and body with weakly ctenoid or cycloid scales; pored lateral-line scales 24; preopercle ridge smooth, edges smooth; one supraneural; supramaxilla absent; basisphenoid present; uroneurals absent; two epurals; fused hypurals 1+2+3+4, fused to terminal centrum; no autogenous haemal spines; one supernumerary dorsal spine; bioluminescent organ under cleithrum; caudal fin forked.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal fin-ray branched and segmented; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; developed gill rakers 12–15; unbranched procurrent rays, longest segmented; pectoral fin-rays 14–16; 10+14 vertebrae; 8 ribs; 5–6 epipleurals; teeth in one row on premaxilla, dentary, vomer, palatine, all villiform, or absent on palatine; six infraorbitals, bony shelf absent on third infraorbital; stomach and intestine black with silvery peritoneum; low crest on PU2.
Distribution. The single described species is known from East Africa, Red Sea, islands in the Indian Ocean, throughout the West Pacific to Japan, onto the Pacific Plate to Marshall Islands, Vanuatu, New Caledonia and Australia.
Remarks. This tribe contains only one species, Verulux cypselurus , corresponding to the clade V in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). Representatives of Verulux (two individuals from Ryukyu Islands and Seychelles) were monophyletic separated by a moderate genetic distance, which may indicate presence of cryptic species. This lineage was recovered far from Rhabdamia (tribe Rhabdamiini ), although their monophyly (genus Rhabdamia sensu Fraser 1972 ) was not rejected by AU test (H05 in Table 6). The single described species has a bioluminescent organ under cleithrum unlike Rhabdamia .
Tribe Zoramiini new name Fraser & Mabuchi
Type genus Zoramia Jordan 1917
Diagnosis. Members of the Apogoninae : dorsal fin VI–I,9; anal fin II,8–9; head and body with ctenoid scales; pored lateral-line scales 24; preopercle ridge smooth, edges serrate; three supraneurals; supramaxilla absent; basisphenoid present; anterior pair of uroneurals; three epurals; five free hypurals; parhypural separate; two autogenous haemal spines; one supernumerary dorsal spine; caudal fin forked.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal-fin ray branched and segmented; second dorsal spine longest, very long and filamentous in some species of both genera; ctenoid scales on predorsal, cheek, breast, two pelvic scales, and body; ctenoid scales on opercle and onto base of caudal fin; cycloid or ctenoid predorsal scales1–3; pored lateral-line scales simple with one pore above and one below midline; pectoral-fin rays 13–17; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; developed gill rakers 7–38; unbranched procurrent rays, longest segmented; 10+14 vertebrae; 8 ribs; 7–8 epipleurals; teeth in one, two or multiple rows on premaxilla, dentary, vomer, palatine, all villiform; six infraorbitals, bony shelf absent or on third infraorbital; stomach and intestine black with silvery peritoneum; low crest on PU2.
Distribution. Species of Zoramia are known from East Africa, Red Sea, islands in the Indian Ocean, throughout the West Pacific to Japan, onto the Pacific Plate to Marshall Islands, Samoa, Tonga, Solomon Islands, New Caledonia and Australia. Species of Fibramia are known from East Africa, some islands in the Indian Ocean, Sri Lanka, throughout the West Pacific to Philippines, Solomon Islands, Santa Cruz Islands, Fiji, Vanuatu, New Caledonia, Australia, Samoa and Tonga.
Remarks. This tribe contains two genera, a new genus Fibramia described below and Zoramia , corresponding to the clade VIII in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). Both genera have species with elongate second dorsal spine, a single supernumerary dorsal spine, and can be found in tidal fresh and brackish water, near shore waters and in lagoons. Fibramia corresponds to the clade of Ostorhinchus - 4 in the present study, and that of Ostorhinchus I in Mabuchi et al. (2006).
Tribe Pristiapogonini new name Fraser & Mabuchi
Type genus Pristiapogon Klunzinger 1870
Diagnosis. Members of the Apogoninae : dorsal fin VI+I,9 or VII+I,9; anal fin II,8; head and body with ctenoid scales; pored lateral-line scales 23–25; preopercle ridge serrate, edges serrate; infraorbitals serrate; three supraneurals; supramaxilla absent; basisphenoid present; one pair of uroneurals or absent; three epurals; five free hypurals; parhypural separate; one or two supernumerary dorsal spines; caudal fin forked.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal-fin ray branched and segmented; ctenoid scales on predorsal, cheek, breast, two pelvic scales, and body; ctenoid scales on opercle and onto base of caudal fin; pored lateral-line scales with multiple pores; pectoral fin-rays 12–16; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; unbranched procurrent rays, longest segmented; teeth in one or multiple rows on premaxilla, dentary, vomer, palatine, all villiform or absent on palatine; six infraorbitals, bony shelf on third infraorbital; anterior ceratohyal smooth or notched; developed gill rakers 10–19; 10+14 vertebrae; 8 ribs; 8 epipleurals; low crest on PU2.
Distribution. Pristiapogon is wide spread from East Africa, Red Sea, Indian Ocean Islands, Indonesia, Philippines, Pacific islands to Hawaii, Japan, French Polynesia and Australia. One species of Pristicon Fraser 1972 is restricted to the West Pacific. Another species reaches out to some islands on the Pacific Plate. One species of Pristicon has been reported from the west coast of India, with a gap between India and Indonesia ( Suresh & Thomas 2007).
Remarks. This tribe contains two genera, Pristiapogon and Pristicon , corresponding to the clade VI in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). Although bootstrap values supporting the tribe were not so high (62 and 56% BPs in ML and MP analyses, respectively), this tribe is morphologically well-defined: relatively large body with serrations on the preopercle ridge (a likely synapomorphy), edges and infraorbitals (a likely synapomorphy). Species of Pristiapogon usually have a darkish single stripe and/or a variable basicaudal spot while Pristicon have bars or saddles under the dorsal fins and spots at the base of the caudal fin or on the opercle.
Tribe Rhabdamiini new name Fraser & Mabuchi
Type genus Rhabdamia Weber 1909
Diagnosis. Members of the Apogoninae : dorsal fin VI+I,9 or VII+I,10–11; anal fin II,11–13; head and body with weakly ctenoid or cycloid; pored lateral-line scales 23–24; preopercle ridge smooth, edges smooth; two supraneurals; supramaxilla absent; basisphenoid present or absent; anterior pair of uroneurals reduced or absent; three epurals; hypurals 1+2+3+4 fused, the plate fused to terminal centrum; parhypural separate; no autogenous haemal spines; 1–2 supernumerary dorsal spines; caudal fin forked.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal-fin ray branched and segmented; pectoral-fin rays 13–17; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; developed gill rakers 17–31; unbranched procurrent rays, longest segmented; villiform teeth in one or two rows on premaxilla, or with a few canines ( Bentuviaichthys ), one villiform row on dentary and vomer, one villiform row or absent on palatine; six infraorbitals, bony shelf reduce or absent on third infraorbital; stomach and intestine pale with melanophores and silvery peritoneum with melanophores; 10+14 vertebrae; low crest on PU2.
Distribution. No reviews of the species have been made, but members have been reported from East Africa, the Red Sea, islands in the Indian Ocean, the West Pacific and out onto the Pacific Plate.
Remarks. This tribe contains one genus Rhabdamia (including two subgenera Rhabdamia and Bentuviaichthys sensu Fraser 1972 ). There are six names (Appendix A). Three species are streamlined apogonines exploiting the near pelagic zone of coral reefs for food. Likely synapomorphies include smooth preopercle ridge and edges, reduced dentition, fused hypurals, and two supraneurals. All species are mostly translucent. In the present molecular analyses, they formed a robust monophyletic group (clade IX) with Yarica hyalosoma ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4) (see also Fig. 7 View FIGURE 7 ), but it is very unlikely based on morphological evidence (for details, see the remarks of the tribe Glossamiini ). In Fraser (1972), subgenus Rhabdamia was classified under the genus Rhabdamia together with the subgenera Bentuviaichthys and Verulux Fraser 1972 . The last lineage, Verulux (clade V), was placed far from the first lineage, Rhabdamia , in the present molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 ). Although monophyly between the two lineages was not rejected by AU test (H05 in Table 6), we recognized each of them in its own tribe for the present (for Verulux , see the tribe Veruluxini ). The present molecular analyses did not include Bentuviaichthys , but it was included in this tribe based on the following morphological characters: fused hypurals 1+2+3+4, epipleurals absent on last three ribs, single row of teeth on dentary, two supraneurals, slender, mostly translucent body and forked caudal fin. Two individuals of Rhabdamia gracilis from two distantly distributed populations ( Fiji and Ryukyu Islands) were paraphyletic to R. spilota Allen & Kuiter 1994 and genetically distant from each other, which may indicate needs of taxonomical revisions of them.
Tribe Siphamiini Smith 1955
Type genus Siphamia Weber 1909
Diagnosis. Members of the Apogoninae : dorsal fin VI–VII + I,7–11; anal fin II,7–11; pored lateral-line scales 0–24; preopercular ridge smooth, edges smooth to serrate; supraneurals 1–2; supramaxilla absent; basisphenoid absent; uroneurals absent; two epurals; hypurals 1+2 and 3+4 fused into two plates, upper plate fused to terminal centrum; parhypural separate; one or two supernumerary dorsal spines; caudal fin emarginate or forked; bacteria bioluminescent system from hyal region to along body above or past anal-fin base.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal-fin ray branched and segmented; ctenoid, cycloid or spinoid scales; median predorsal scales 0–6; pectoral rays 11–16; segmented principal caudal rays 9+8, 15 branched, upper and lower unbranched; unbranched procurrent rays, longest segmented; villiform teeth in one or multiple rows on premaxilla, dentary, vomer, palatine, or absent on palatine; developed gill rakers 6–18; lateral-line scales usually with a vertical row of free neuromasts; six infraorbitals, bony shelf on third infraorbital; anterior ceratohyal smooth or notched; developed gill rakers 10–26; 10+14 vertebrae; 8 ribs; 8 epipleurals; stomach, intestine and peritoneum generally pale with variously sized melanophores; low crest on PU2.
Distribution. Representatives of the tribe are found from East Africa, Red Sea, islands in the Indian Ocean, throughout the West Pacific to Japan, onto the Pacific Plate to French Polynesia and Australia ( Gon & Allen 2012).
Remarks. This tribe contains one genus Siphamia , corresponding to the clade VII in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). All species of Siphamia have bioluminescent bacteria in a specialized organ (a synapomorphy for the species) unique among apogonines. Smith (1955) proposed placing species of Siphamia in its own subfamily Siphamiinae . Our results based on analysis of five of the twenty-three species suggest that Siphamia roseigaster Ramsay & Ogilby 1887 could be recognized in its own genus Adenapogon McCulloch 1921 , because it was placed relatively far from the remaining species. Gon & Allen's (2012) results based on morphology suggest that two other Australian species belong in Adenapogon and that Fodifoa Whitley 1936 is available for another group of species. We defer to Ofer Gon who is continuing to work on relationships within this tribe ( Gon & Allen 2012).
Tribe Sphaeramiini new name Fraser & Mabuchi
Type genus Sphaeramia Fowler & Bean 1930
Diagnosis. Members of the Apogoninae : dorsal fin VII+I,9–10, VII(I)+I,9–14 or VIII+I,9–10 with eighth spine reduced to a tiny visible spine or a nubbin hidden under skin supported by a free sixth distal radial; anal fin II,8–13; head and body with ctenoid scales; pored lateral-line scales 23–27; preopercle ridge smooth, edges serrate; 2–3 supraneurals; supramaxilla absent; basisphenoid present; one pair of uroneurals; three epurals; five free hypurals or 1+2 fused and 3+4 fused with 3+4 fused to terminal centrum; parhypural separate; two autogenous haemal spines; two supernumerary dorsal spines; caudal fin forked, emarginate or truncate.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal-fin ray branched and segmented; ctenoid scales on predorsal, cheek, breast, two pelvic scales, and body; ctenoid scales on opercle and onto base of caudal fin; 1–3 predorsal scales cycloid or ctenoid; pored lateral-line scales with multiple pores; pectoral-fin rays 13–17; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; developed gill rakers 7–38; unbranched procurrent rays, longest segmented or spinous; third or fourth dorsal spine longest; 10+14 or 10+15 vertebrae; 8 ribs; 8–9 epipleurals; teeth in one or multiple rows on premaxilla, dentary, vomer, palatine, all villiform; six infraorbitals, bony shelf on third infraorbital; stomach and intestine pale or black with pale peritoneum; low crest on PU2.
Distribution. Species of Apogonichthyoides are found throughout the Indo-West Pacific from East Africa, Red Sea, India, some island of the Indian Ocean, throughout Indonesia, Philippines to Japan, New Guinea, Australia and the Solomon Islands. Some species have restricted distribution and some are rare in collections. Species of Jaydia Smith 1961 have a more continental distribution, mostly caught in trawls, and are widespread from Africa to Japan, New Guinea, Australia and larger islands in the Coral Sea. Representatives of Nectamia are found on reef habitats from East Africa, Red Sea, islands in the Indian Ocean, throughout the West Pacific to Japan, onto the Pacific Plate out to the Line Islands and French Polynesia. Sphaeramia , represented by two species, one found in mangrove habitats and one on coral reefs, also have wide distribution in the Indo-West Pacific. Quinca , represented by a single species, is known from northwestern coast of Australia.
Remarks. This tribe contains six genera, Apogonichthyoides , Jaydia , Nectamia , Pterapogon Koumans 1933 , Quinca and Sphaeramia , almost corresponding to the clade IV in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). First three of the six genera were formerly classified under Ostorhinchus , corresponding to "barred group" of Ostorhinchus ( Ostorhinchus II ) in Mabuchi et al. (2006). Two of the remaining three genera ( Pterapogon and Sphaeramia ) have distinct bar(s) on body. In addition to the six genera, corresponding molecular clade included three species of Ostorhinchus -2 ( O. hoevenii Bleeker 1854b , O. ishigakiensis Ida & Moyer 1974 , and O. rueppellii Günther 1859 ). But there are no morphological characters that support this association. We defer taxonomical action on this possible relationship for the present. This clade further included another species of the former Ostorhinchus ( Ostorhinchus -1). For this species, the latest authors ( Allen & Erdmann 2012) provisionally used Apogonichthyoides as its genus name, and we followed it here, but such application of the name made this genus paraphyletic in our molecular trees. Based on its morphological features, we tentatively included it [ Apogonichthyoides (?) melas ] in this tribe, but further study including more Sphaeramiini species will be needed to clarify its taxonomic status. In the present molecular analyses, phylogenetic position of Pterapogon remained unsettled ( Table 5). It was sister to the clade VII ( Siphamiini ) in ML tree ( Fig. 2 View FIGURE 2 ), sister to Ostorhinchus margaritophorus in MP tree-A ( Fig. 4 View FIGURE 4 ), and nested within the clade XII ( Ostorhinchini ) in the MP tree-B ( Fig. 5 View FIGURE 5 ). It formed a polytomy with O. margaritophorus , clade VII, clade VIII, and a large clade including clades IX–XII in BA tree ( Fig. 3 View FIGURE 3 ). Pterapogon has, however, a synapomorphy (spinous procurrent caudal rays) uniquely held with Sphaeramia as well as similarities in color patterns and body shape, but differs with fused hypurals 1+2 and 3+4. Based on these morphological features, we tentatively included this species in the tribe Sphaeramiini . Monophyly of them was not rejected by AU test (H02 in Table 6), although its inclusion within Ostorhinchini was also not rejected (H03 in Table 6). We had no good DNA extracts from tissue of Quinca . This monotypic genus has morphological characteristics and color pattern suggestive of a relationship with Apogonichthyoides , and was provisionally place in the Sphaeramiini .
Tribe Veruluxini new name Fraser & Mabuchi
Type genus Verulux Fraser 1972
Diagnosis. Members of the Apogoninae : dorsal fin VI–I,9; anal fin II,9; head and body with weakly ctenoid or cycloid scales; pored lateral-line scales 24; preopercle ridge smooth, edges smooth; one supraneural; supramaxilla absent; basisphenoid present; uroneurals absent; two epurals; fused hypurals 1+2+3+4, fused to terminal centrum; no autogenous haemal spines; one supernumerary dorsal spine; bioluminescent organ under cleithrum; caudal fin forked.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal fin-ray branched and segmented; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; developed gill rakers 12–15; unbranched procurrent rays, longest segmented; pectoral fin-rays 14–16; 10+14 vertebrae; 8 ribs; 5–6 epipleurals; teeth in one row on premaxilla, dentary, vomer, palatine, all villiform, or absent on palatine; six infraorbitals, bony shelf absent on third infraorbital; stomach and intestine black with silvery peritoneum; low crest on PU2.
Distribution. The single described species is known from East Africa, Red Sea, islands in the Indian Ocean, throughout the West Pacific to Japan, onto the Pacific Plate to Marshall Islands, Vanuatu, New Caledonia and Australia.
Remarks. This tribe contains only one species, Verulux cypselurus , corresponding to the clade V in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). Representatives of Verulux (two individuals from Ryukyu Islands and Seychelles) were monophyletic separated by a moderate genetic distance, which may indicate presence of cryptic species. This lineage was recovered far from Rhabdamia (tribe Rhabdamiini ), although their monophyly (genus Rhabdamia sensu Fraser 1972 ) was not rejected by AU test (H05 in Table 6). The single described species has a bioluminescent organ under cleithrum unlike Rhabdamia .
Tribe Zoramiini new name Fraser & Mabuchi
Type genus Zoramia Jordan 1917
Diagnosis. Members of the Apogoninae : dorsal fin VI–I,9; anal fin II,8–9; head and body with ctenoid scales; pored lateral-line scales 24; preopercle ridge smooth, edges serrate; three supraneurals; supramaxilla absent; basisphenoid present; anterior pair of uroneurals; three epurals; five free hypurals; parhypural separate; two autogenous haemal spines; one supernumerary dorsal spine; caudal fin forked.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal-fin ray branched and segmented; second dorsal spine longest, very long and filamentous in some species of both genera; ctenoid scales on predorsal, cheek, breast, two pelvic scales, and body; ctenoid scales on opercle and onto base of caudal fin; cycloid or ctenoid predorsal scales1–3; pored lateral-line scales simple with one pore above and one below midline; pectoral-fin rays 13–17; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; developed gill rakers 7–38; unbranched procurrent rays, longest segmented; 10+14 vertebrae; 8 ribs; 7–8 epipleurals; teeth in one, two or multiple rows on premaxilla, dentary, vomer, palatine, all villiform; six infraorbitals, bony shelf absent or on third infraorbital; stomach and intestine black with silvery peritoneum; low crest on PU2.
Distribution. Species of Zoramia are known from East Africa, Red Sea, islands in the Indian Ocean, throughout the West Pacific to Japan, onto the Pacific Plate to Marshall Islands, Samoa, Tonga, Solomon Islands, New Caledonia and Australia. Species of Fibramia are known from East Africa, some islands in the Indian Ocean, Sri Lanka, throughout the West Pacific to Philippines, Solomon Islands, Santa Cruz Islands, Fiji, Vanuatu, New Caledonia, Australia, Samoa and Tonga.
Remarks. This tribe contains two genera, a new genus Fibramia described below and Zoramia , corresponding to the clade VIII in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). Both genera have species with elongate second dorsal spine, a single supernumerary dorsal spine, and can be found in tidal fresh and brackish water, near shore waters and in lagoons. Fibramia corresponds to the clade of Ostorhinchus - 4 in the present study, and that of Ostorhinchus I in Mabuchi et al. (2006).
Tribe Rhabdamiini new name Fraser & Mabuchi
Type genus Rhabdamia Weber 1909
Diagnosis. Members of the Apogoninae : dorsal fin VI+I,9 or VII+I,10–11; anal fin II,11–13; head and body with weakly ctenoid or cycloid; pored lateral-line scales 23–24; preopercle ridge smooth, edges smooth; two supraneurals; supramaxilla absent; basisphenoid present or absent; anterior pair of uroneurals reduced or absent; three epurals; hypurals 1+2+3+4 fused, the plate fused to terminal centrum; parhypural separate; no autogenous haemal spines; 1–2 supernumerary dorsal spines; caudal fin forked.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal-fin ray branched and segmented; pectoral-fin rays 13–17; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; developed gill rakers 17–31; unbranched procurrent rays, longest segmented; villiform teeth in one or two rows on premaxilla, or with a few canines ( Bentuviaichthys ), one villiform row on dentary and vomer, one villiform row or absent on palatine; six infraorbitals, bony shelf reduce or absent on third infraorbital; stomach and intestine pale with melanophores and silvery peritoneum with melanophores; 10+14 vertebrae; low crest on PU2.
Distribution. No reviews of the species have been made, but members have been reported from East Africa, the Red Sea, islands in the Indian Ocean, the West Pacific and out onto the Pacific Plate.
Remarks. This tribe contains one genus Rhabdamia (including two subgenera Rhabdamia and Bentuviaichthys sensu Fraser 1972 ). There are six names (Appendix A). Three species are streamlined apogonines exploiting the near pelagic zone of coral reefs for food. Likely synapomorphies include smooth preopercle ridge and edges, reduced dentition, fused hypurals, and two supraneurals. All species are mostly translucent. In the present molecular analyses, they formed a robust monophyletic group (clade IX) with Yarica hyalosoma ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4) (see also Fig. 7 View FIGURE 7 ), but it is very unlikely based on morphological evidence (for details, see the remarks of the tribe Glossamiini ). In Fraser (1972), subgenus Rhabdamia was classified under the genus Rhabdamia together with the subgenera Bentuviaichthys and Verulux Fraser 1972 . The last lineage, Verulux (clade V), was placed far from the first lineage, Rhabdamia , in the present molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 ). Although monophyly between the two lineages was not rejected by AU test (H05 in Table 6), we recognized each of them in its own tribe for the present (for Verulux , see the tribe Veruluxini ). The present molecular analyses did not include Bentuviaichthys , but it was included in this tribe based on the following morphological characters: fused hypurals 1+2+3+4, epipleurals absent on last three ribs, single row of teeth on dentary, two supraneurals, slender, mostly translucent body and forked caudal fin. Two individuals of Rhabdamia gracilis from two distantly distributed populations ( Fiji and Ryukyu Islands) were paraphyletic to R. spilota Allen & Kuiter 1994 and genetically distant from each other, which may indicate needs of taxonomical revisions of them.
Tribe Siphamiini Smith 1955
Type genus Siphamia Weber 1909
Diagnosis. Members of the Apogoninae : dorsal fin VI–VII + I,7–11; anal fin II,7–11; pored lateral-line scales 0–24; preopercular ridge smooth, edges smooth to serrate; supraneurals 1–2; supramaxilla absent; basisphenoid absent; uroneurals absent; two epurals; hypurals 1+2 and 3+4 fused into two plates, upper plate fused to terminal centrum; parhypural separate; one or two supernumerary dorsal spines; caudal fin emarginate or forked; bacteria bioluminescent system from hyal region to along body above or past anal-fin base.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal-fin ray branched and segmented; ctenoid, cycloid or spinoid scales; median predorsal scales 0–6; pectoral rays 11–16; segmented principal caudal rays 9+8, 15 branched, upper and lower unbranched; unbranched procurrent rays, longest segmented; villiform teeth in one or multiple rows on premaxilla, dentary, vomer, palatine, or absent on palatine; developed gill rakers 6–18; lateral-line scales usually with a vertical row of free neuromasts; six infraorbitals, bony shelf on third infraorbital; anterior ceratohyal smooth or notched; developed gill rakers 10–26; 10+14 vertebrae; 8 ribs; 8 epipleurals; stomach, intestine and peritoneum generally pale with variously sized melanophores; low crest on PU2.
Distribution. Representatives of the tribe are found from East Africa, Red Sea, islands in the Indian Ocean, throughout the West Pacific to Japan, onto the Pacific Plate to French Polynesia and Australia ( Gon & Allen 2012).
Remarks. This tribe contains one genus Siphamia , corresponding to the clade VII in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). All species of Siphamia have bioluminescent bacteria in a specialized organ (a synapomorphy for the species) unique among apogonines. Smith (1955) proposed placing species of Siphamia in its own subfamily Siphamiinae . Our results based on analysis of five of the twenty-three species suggest that Siphamia roseigaster Ramsay & Ogilby 1887 could be recognized in its own genus Adenapogon McCulloch 1921 , because it was placed relatively far from the remaining species. Gon & Allen's (2012) results based on morphology suggest that two other Australian species belong in Adenapogon and that Fodifoa Whitley 1936 is available for another group of species. We defer to Ofer Gon who is continuing to work on relationships within this tribe ( Gon & Allen 2012).
Tribe Sphaeramiini new name Fraser & Mabuchi
Type genus Sphaeramia Fowler & Bean 1930
Diagnosis. Members of the Apogoninae : dorsal fin VII+I,9–10, VII(I)+I,9–14 or VIII+I,9–10 with eighth spine reduced to a tiny visible spine or a nubbin hidden under skin supported by a free sixth distal radial; anal fin II,8–13; head and body with ctenoid scales; pored lateral-line scales 23–27; preopercle ridge smooth, edges serrate; 2–3 supraneurals; supramaxilla absent; basisphenoid present; one pair of uroneurals; three epurals; five free hypurals or 1+2 fused and 3+4 fused with 3+4 fused to terminal centrum; parhypural separate; two autogenous haemal spines; two supernumerary dorsal spines; caudal fin forked, emarginate or truncate.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal-fin ray branched and segmented; ctenoid scales on predorsal, cheek, breast, two pelvic scales, and body; ctenoid scales on opercle and onto base of caudal fin; 1–3 predorsal scales cycloid or ctenoid; pored lateral-line scales with multiple pores; pectoral-fin rays 13–17; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; developed gill rakers 7–38; unbranched procurrent rays, longest segmented or spinous; third or fourth dorsal spine longest; 10+14 or 10+15 vertebrae; 8 ribs; 8–9 epipleurals; teeth in one or multiple rows on premaxilla, dentary, vomer, palatine, all villiform; six infraorbitals, bony shelf on third infraorbital; stomach and intestine pale or black with pale peritoneum; low crest on PU2.
Distribution. Species of Apogonichthyoides are found throughout the Indo-West Pacific from East Africa, Red Sea, India, some island of the Indian Ocean, throughout Indonesia, Philippines to Japan, New Guinea, Australia and the Solomon Islands. Some species have restricted distribution and some are rare in collections. Species of Jaydia Smith 1961 have a more continental distribution, mostly caught in trawls, and are widespread from Africa to Japan, New Guinea, Australia and larger islands in the Coral Sea. Representatives of Nectamia are found on reef habitats from East Africa, Red Sea, islands in the Indian Ocean, throughout the West Pacific to Japan, onto the Pacific Plate out to the Line Islands and French Polynesia. Sphaeramia , represented by two species, one found in mangrove habitats and one on coral reefs, also have wide distribution in the Indo-West Pacific. Quinca , represented by a single species, is known from northwestern coast of Australia.
Remarks. This tribe contains six genera, Apogonichthyoides , Jaydia , Nectamia , Pterapogon Koumans 1933 , Quinca and Sphaeramia , almost corresponding to the clade IV in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). First three of the six genera were formerly classified under Ostorhinchus , corresponding to "barred group" of Ostorhinchus ( Ostorhinchus II ) in Mabuchi et al. (2006). Two of the remaining three genera ( Pterapogon and Sphaeramia ) have distinct bar(s) on body. In addition to the six genera, corresponding molecular clade included three species of Ostorhinchus -2 ( O. hoevenii Bleeker 1854b , O. ishigakiensis Ida & Moyer 1974 , and O. rueppellii Günther 1859 ). But there are no morphological characters that support this association. We defer taxonomical action on this possible relationship for the present. This clade further included another species of the former Ostorhinchus ( Ostorhinchus -1). For this species, the latest authors ( Allen & Erdmann 2012) provisionally used Apogonichthyoides as its genus name, and we followed it here, but such application of the name made this genus paraphyletic in our molecular trees. Based on its morphological features, we tentatively included it [ Apogonichthyoides (?) melas ] in this tribe, but further study including more Sphaeramiini species will be needed to clarify its taxonomic status. In the present molecular analyses, phylogenetic position of Pterapogon remained unsettled ( Table 5). It was sister to the clade VII ( Siphamiini ) in ML tree ( Fig. 2 View FIGURE 2 ), sister to Ostorhinchus margaritophorus in MP tree-A ( Fig. 4 View FIGURE 4 ), and nested within the clade XII ( Ostorhinchini ) in the MP tree-B ( Fig. 5 View FIGURE 5 ). It formed a polytomy with O. margaritophorus , clade VII, clade VIII, and a large clade including clades IX–XII in BA tree ( Fig. 3 View FIGURE 3 ). Pterapogon has, however, a synapomorphy (spinous procurrent caudal rays) uniquely held with Sphaeramia as well as similarities in color patterns and body shape, but differs with fused hypurals 1+2 and 3+4. Based on these morphological features, we tentatively included this species in the tribe Sphaeramiini . Monophyly of them was not rejected by AU test (H02 in Table 6), although its inclusion within Ostorhinchini was also not rejected (H03 in Table 6). We had no good DNA extracts from tissue of Quinca . This monotypic genus has morphological characteristics and color pattern suggestive of a relationship with Apogonichthyoides , and was provisionally place in the Sphaeramiini .
Tribe Veruluxini new name Fraser & Mabuchi
Type genus Verulux Fraser 1972
Diagnosis. Members of the Apogoninae : dorsal fin VI–I,9; anal fin II,9; head and body with weakly ctenoid or cycloid scales; pored lateral-line scales 24; preopercle ridge smooth, edges smooth; one supraneural; supramaxilla absent; basisphenoid present; uroneurals absent; two epurals; fused hypurals 1+2+3+4, fused to terminal centrum; no autogenous haemal spines; one supernumerary dorsal spine; bioluminescent organ under cleithrum; caudal fin forked.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal fin-ray branched and segmented; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; developed gill rakers 12–15; unbranched procurrent rays, longest segmented; pectoral fin-rays 14–16; 10+14 vertebrae; 8 ribs; 5–6 epipleurals; teeth in one row on premaxilla, dentary, vomer, palatine, all villiform, or absent on palatine; six infraorbitals, bony shelf absent on third infraorbital; stomach and intestine black with silvery peritoneum; low crest on PU2.
Distribution. The single described species is known from East Africa, Red Sea, islands in the Indian Ocean, throughout the West Pacific to Japan, onto the Pacific Plate to Marshall Islands, Vanuatu, New Caledonia and Australia.
Remarks. This tribe contains only one species, Verulux cypselurus , corresponding to the clade V in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). Representatives of Verulux (two individuals from Ryukyu Islands and Seychelles) were monophyletic separated by a moderate genetic distance, which may indicate presence of cryptic species. This lineage was recovered far from Rhabdamia (tribe Rhabdamiini ), although their monophyly (genus Rhabdamia sensu Fraser 1972 ) was not rejected by AU test (H05 in Table 6). The single described species has a bioluminescent organ under cleithrum unlike Rhabdamia .
Tribe Zoramiini new name Fraser & Mabuchi
Type genus Zoramia Jordan 1917
Diagnosis. Members of the Apogoninae : dorsal fin VI–I,9; anal fin II,8–9; head and body with ctenoid scales; pored lateral-line scales 24; preopercle ridge smooth, edges serrate; three supraneurals; supramaxilla absent; basisphenoid present; anterior pair of uroneurals; three epurals; five free hypurals; parhypural separate; two autogenous haemal spines; one supernumerary dorsal spine; caudal fin forked.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal-fin ray branched and segmented; second dorsal spine longest, very long and filamentous in some species of both genera; ctenoid scales on predorsal, cheek, breast, two pelvic scales, and body; ctenoid scales on opercle and onto base of caudal fin; cycloid or ctenoid predorsal scales1–3; pored lateral-line scales simple with one pore above and one below midline; pectoral-fin rays 13–17; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; developed gill rakers 7–38; unbranched procurrent rays, longest segmented; 10+14 vertebrae; 8 ribs; 7–8 epipleurals; teeth in one, two or multiple rows on premaxilla, dentary, vomer, palatine, all villiform; six infraorbitals, bony shelf absent or on third infraorbital; stomach and intestine black with silvery peritoneum; low crest on PU2.
Distribution. Species of Zoramia are known from East Africa, Red Sea, islands in the Indian Ocean, throughout the West Pacific to Japan, onto the Pacific Plate to Marshall Islands, Samoa, Tonga, Solomon Islands, New Caledonia and Australia. Species of Fibramia are known from East Africa, some islands in the Indian Ocean, Sri Lanka, throughout the West Pacific to Philippines, Solomon Islands, Santa Cruz Islands, Fiji, Vanuatu, New Caledonia, Australia, Samoa and Tonga.
Remarks. This tribe contains two genera, a new genus Fibramia described below and Zoramia , corresponding to the clade VIII in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). Both genera have species with elongate second dorsal spine, a single supernumerary dorsal spine, and can be found in tidal fresh and brackish water, near shore waters and in lagoons. Fibramia corresponds to the clade of Ostorhinchus - 4 in the present study, and that of Ostorhinchus I in Mabuchi et al. (2006).
Tribe Siphamiini Smith 1955
Type genus Siphamia Weber 1909
Diagnosis. Members of the Apogoninae : dorsal fin VI–VII + I,7–11; anal fin II,7–11; pored lateral-line scales 0–24; preopercular ridge smooth, edges smooth to serrate; supraneurals 1–2; supramaxilla absent; basisphenoid absent; uroneurals absent; two epurals; hypurals 1+2 and 3+4 fused into two plates, upper plate fused to terminal centrum; parhypural separate; one or two supernumerary dorsal spines; caudal fin emarginate or forked; bacteria bioluminescent system from hyal region to along body above or past anal-fin base.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal-fin ray branched and segmented; ctenoid, cycloid or spinoid scales; median predorsal scales 0–6; pectoral rays 11–16; segmented principal caudal rays 9+8, 15 branched, upper and lower unbranched; unbranched procurrent rays, longest segmented; villiform teeth in one or multiple rows on premaxilla, dentary, vomer, palatine, or absent on palatine; developed gill rakers 6–18; lateral-line scales usually with a vertical row of free neuromasts; six infraorbitals, bony shelf on third infraorbital; anterior ceratohyal smooth or notched; developed gill rakers 10–26; 10+14 vertebrae; 8 ribs; 8 epipleurals; stomach, intestine and peritoneum generally pale with variously sized melanophores; low crest on PU2.
Distribution. Representatives of the tribe are found from East Africa, Red Sea, islands in the Indian Ocean, throughout the West Pacific to Japan, onto the Pacific Plate to French Polynesia and Australia ( Gon & Allen 2012).
Remarks. This tribe contains one genus Siphamia , corresponding to the clade VII in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). All species of Siphamia have bioluminescent bacteria in a specialized organ (a synapomorphy for the species) unique among apogonines. Smith (1955) proposed placing species of Siphamia in its own subfamily Siphamiinae . Our results based on analysis of five of the twenty-three species suggest that Siphamia roseigaster Ramsay & Ogilby 1887 could be recognized in its own genus Adenapogon McCulloch 1921 , because it was placed relatively far from the remaining species. Gon & Allen's (2012) results based on morphology suggest that two other Australian species belong in Adenapogon and that Fodifoa Whitley 1936 is available for another group of species. We defer to Ofer Gon who is continuing to work on relationships within this tribe ( Gon & Allen 2012).
Tribe Sphaeramiini new name Fraser & Mabuchi
Type genus Sphaeramia Fowler & Bean 1930
Diagnosis. Members of the Apogoninae : dorsal fin VII+I,9–10, VII(I)+I,9–14 or VIII+I,9–10 with eighth spine reduced to a tiny visible spine or a nubbin hidden under skin supported by a free sixth distal radial; anal fin II,8–13; head and body with ctenoid scales; pored lateral-line scales 23–27; preopercle ridge smooth, edges serrate; 2–3 supraneurals; supramaxilla absent; basisphenoid present; one pair of uroneurals; three epurals; five free hypurals or 1+2 fused and 3+4 fused with 3+4 fused to terminal centrum; parhypural separate; two autogenous haemal spines; two supernumerary dorsal spines; caudal fin forked, emarginate or truncate.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal-fin ray branched and segmented; ctenoid scales on predorsal, cheek, breast, two pelvic scales, and body; ctenoid scales on opercle and onto base of caudal fin; 1–3 predorsal scales cycloid or ctenoid; pored lateral-line scales with multiple pores; pectoral-fin rays 13–17; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; developed gill rakers 7–38; unbranched procurrent rays, longest segmented or spinous; third or fourth dorsal spine longest; 10+14 or 10+15 vertebrae; 8 ribs; 8–9 epipleurals; teeth in one or multiple rows on premaxilla, dentary, vomer, palatine, all villiform; six infraorbitals, bony shelf on third infraorbital; stomach and intestine pale or black with pale peritoneum; low crest on PU2.
Distribution. Species of Apogonichthyoides are found throughout the Indo-West Pacific from East Africa, Red Sea, India, some island of the Indian Ocean, throughout Indonesia, Philippines to Japan, New Guinea, Australia and the Solomon Islands. Some species have restricted distribution and some are rare in collections. Species of Jaydia Smith 1961 have a more continental distribution, mostly caught in trawls, and are widespread from Africa to Japan, New Guinea, Australia and larger islands in the Coral Sea. Representatives of Nectamia are found on reef habitats from East Africa, Red Sea, islands in the Indian Ocean, throughout the West Pacific to Japan, onto the Pacific Plate out to the Line Islands and French Polynesia. Sphaeramia , represented by two species, one found in mangrove habitats and one on coral reefs, also have wide distribution in the Indo-West Pacific. Quinca , represented by a single species, is known from northwestern coast of Australia.
Remarks. This tribe contains six genera, Apogonichthyoides , Jaydia , Nectamia , Pterapogon Koumans 1933 , Quinca and Sphaeramia , almost corresponding to the clade IV in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). First three of the six genera were formerly classified under Ostorhinchus , corresponding to "barred group" of Ostorhinchus ( Ostorhinchus II ) in Mabuchi et al. (2006). Two of the remaining three genera ( Pterapogon and Sphaeramia ) have distinct bar(s) on body. In addition to the six genera, corresponding molecular clade included three species of Ostorhinchus -2 ( O. hoevenii Bleeker 1854b , O. ishigakiensis Ida & Moyer 1974 , and O. rueppellii Günther 1859 ). But there are no morphological characters that support this association. We defer taxonomical action on this possible relationship for the present. This clade further included another species of the former Ostorhinchus ( Ostorhinchus -1). For this species, the latest authors ( Allen & Erdmann 2012) provisionally used Apogonichthyoides as its genus name, and we followed it here, but such application of the name made this genus paraphyletic in our molecular trees. Based on its morphological features, we tentatively included it [ Apogonichthyoides (?) melas ] in this tribe, but further study including more Sphaeramiini species will be needed to clarify its taxonomic status. In the present molecular analyses, phylogenetic position of Pterapogon remained unsettled ( Table 5). It was sister to the clade VII ( Siphamiini ) in ML tree ( Fig. 2 View FIGURE 2 ), sister to Ostorhinchus margaritophorus in MP tree-A ( Fig. 4 View FIGURE 4 ), and nested within the clade XII ( Ostorhinchini ) in the MP tree-B ( Fig. 5 View FIGURE 5 ). It formed a polytomy with O. margaritophorus , clade VII, clade VIII, and a large clade including clades IX–XII in BA tree ( Fig. 3 View FIGURE 3 ). Pterapogon has, however, a synapomorphy (spinous procurrent caudal rays) uniquely held with Sphaeramia as well as similarities in color patterns and body shape, but differs with fused hypurals 1+2 and 3+4. Based on these morphological features, we tentatively included this species in the tribe Sphaeramiini . Monophyly of them was not rejected by AU test (H02 in Table 6), although its inclusion within Ostorhinchini was also not rejected (H03 in Table 6). We had no good DNA extracts from tissue of Quinca . This monotypic genus has morphological characteristics and color pattern suggestive of a relationship with Apogonichthyoides , and was provisionally place in the Sphaeramiini .
Tribe Veruluxini new name Fraser & Mabuchi
Type genus Verulux Fraser 1972
Diagnosis. Members of the Apogoninae : dorsal fin VI–I,9; anal fin II,9; head and body with weakly ctenoid or cycloid scales; pored lateral-line scales 24; preopercle ridge smooth, edges smooth; one supraneural; supramaxilla absent; basisphenoid present; uroneurals absent; two epurals; fused hypurals 1+2+3+4, fused to terminal centrum; no autogenous haemal spines; one supernumerary dorsal spine; bioluminescent organ under cleithrum; caudal fin forked.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal fin-ray branched and segmented; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; developed gill rakers 12–15; unbranched procurrent rays, longest segmented; pectoral fin-rays 14–16; 10+14 vertebrae; 8 ribs; 5–6 epipleurals; teeth in one row on premaxilla, dentary, vomer, palatine, all villiform, or absent on palatine; six infraorbitals, bony shelf absent on third infraorbital; stomach and intestine black with silvery peritoneum; low crest on PU2.
Distribution. The single described species is known from East Africa, Red Sea, islands in the Indian Ocean, throughout the West Pacific to Japan, onto the Pacific Plate to Marshall Islands, Vanuatu, New Caledonia and Australia.
Remarks. This tribe contains only one species, Verulux cypselurus , corresponding to the clade V in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). Representatives of Verulux (two individuals from Ryukyu Islands and Seychelles) were monophyletic separated by a moderate genetic distance, which may indicate presence of cryptic species. This lineage was recovered far from Rhabdamia (tribe Rhabdamiini ), although their monophyly (genus Rhabdamia sensu Fraser 1972 ) was not rejected by AU test (H05 in Table 6). The single described species has a bioluminescent organ under cleithrum unlike Rhabdamia .
Tribe Zoramiini new name Fraser & Mabuchi
Type genus Zoramia Jordan 1917
Diagnosis. Members of the Apogoninae : dorsal fin VI–I,9; anal fin II,8–9; head and body with ctenoid scales; pored lateral-line scales 24; preopercle ridge smooth, edges serrate; three supraneurals; supramaxilla absent; basisphenoid present; anterior pair of uroneurals; three epurals; five free hypurals; parhypural separate; two autogenous haemal spines; one supernumerary dorsal spine; caudal fin forked.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal-fin ray branched and segmented; second dorsal spine longest, very long and filamentous in some species of both genera; ctenoid scales on predorsal, cheek, breast, two pelvic scales, and body; ctenoid scales on opercle and onto base of caudal fin; cycloid or ctenoid predorsal scales1–3; pored lateral-line scales simple with one pore above and one below midline; pectoral-fin rays 13–17; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; developed gill rakers 7–38; unbranched procurrent rays, longest segmented; 10+14 vertebrae; 8 ribs; 7–8 epipleurals; teeth in one, two or multiple rows on premaxilla, dentary, vomer, palatine, all villiform; six infraorbitals, bony shelf absent or on third infraorbital; stomach and intestine black with silvery peritoneum; low crest on PU2.
Distribution. Species of Zoramia are known from East Africa, Red Sea, islands in the Indian Ocean, throughout the West Pacific to Japan, onto the Pacific Plate to Marshall Islands, Samoa, Tonga, Solomon Islands, New Caledonia and Australia. Species of Fibramia are known from East Africa, some islands in the Indian Ocean, Sri Lanka, throughout the West Pacific to Philippines, Solomon Islands, Santa Cruz Islands, Fiji, Vanuatu, New Caledonia, Australia, Samoa and Tonga.
Remarks. This tribe contains two genera, a new genus Fibramia described below and Zoramia , corresponding to the clade VIII in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). Both genera have species with elongate second dorsal spine, a single supernumerary dorsal spine, and can be found in tidal fresh and brackish water, near shore waters and in lagoons. Fibramia corresponds to the clade of Ostorhinchus - 4 in the present study, and that of Ostorhinchus I in Mabuchi et al. (2006).
Tribe Sphaeramiini new name Fraser & Mabuchi
Type genus Sphaeramia Fowler & Bean 1930
Diagnosis. Members of the Apogoninae : dorsal fin VII+I,9–10, VII(I)+I,9–14 or VIII+I,9–10 with eighth spine reduced to a tiny visible spine or a nubbin hidden under skin supported by a free sixth distal radial; anal fin II,8–13; head and body with ctenoid scales; pored lateral-line scales 23–27; preopercle ridge smooth, edges serrate; 2–3 supraneurals; supramaxilla absent; basisphenoid present; one pair of uroneurals; three epurals; five free hypurals or 1+2 fused and 3+4 fused with 3+4 fused to terminal centrum; parhypural separate; two autogenous haemal spines; two supernumerary dorsal spines; caudal fin forked, emarginate or truncate.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal-fin ray branched and segmented; ctenoid scales on predorsal, cheek, breast, two pelvic scales, and body; ctenoid scales on opercle and onto base of caudal fin; 1–3 predorsal scales cycloid or ctenoid; pored lateral-line scales with multiple pores; pectoral-fin rays 13–17; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; developed gill rakers 7–38; unbranched procurrent rays, longest segmented or spinous; third or fourth dorsal spine longest; 10+14 or 10+15 vertebrae; 8 ribs; 8–9 epipleurals; teeth in one or multiple rows on premaxilla, dentary, vomer, palatine, all villiform; six infraorbitals, bony shelf on third infraorbital; stomach and intestine pale or black with pale peritoneum; low crest on PU2.
Distribution. Species of Apogonichthyoides are found throughout the Indo-West Pacific from East Africa, Red Sea, India, some island of the Indian Ocean, throughout Indonesia, Philippines to Japan, New Guinea, Australia and the Solomon Islands. Some species have restricted distribution and some are rare in collections. Species of Jaydia Smith 1961 have a more continental distribution, mostly caught in trawls, and are widespread from Africa to Japan, New Guinea, Australia and larger islands in the Coral Sea. Representatives of Nectamia are found on reef habitats from East Africa, Red Sea, islands in the Indian Ocean, throughout the West Pacific to Japan, onto the Pacific Plate out to the Line Islands and French Polynesia. Sphaeramia , represented by two species, one found in mangrove habitats and one on coral reefs, also have wide distribution in the Indo-West Pacific. Quinca , represented by a single species, is known from northwestern coast of Australia.
Remarks. This tribe contains six genera, Apogonichthyoides , Jaydia , Nectamia , Pterapogon Koumans 1933 , Quinca and Sphaeramia , almost corresponding to the clade IV in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). First three of the six genera were formerly classified under Ostorhinchus , corresponding to "barred group" of Ostorhinchus ( Ostorhinchus II ) in Mabuchi et al. (2006). Two of the remaining three genera ( Pterapogon and Sphaeramia ) have distinct bar(s) on body. In addition to the six genera, corresponding molecular clade included three species of Ostorhinchus -2 ( O. hoevenii Bleeker 1854b , O. ishigakiensis Ida & Moyer 1974 , and O. rueppellii Günther 1859 ). But there are no morphological characters that support this association. We defer taxonomical action on this possible relationship for the present. This clade further included another species of the former Ostorhinchus ( Ostorhinchus -1). For this species, the latest authors ( Allen & Erdmann 2012) provisionally used Apogonichthyoides as its genus name, and we followed it here, but such application of the name made this genus paraphyletic in our molecular trees. Based on its morphological features, we tentatively included it [ Apogonichthyoides (?) melas ] in this tribe, but further study including more Sphaeramiini species will be needed to clarify its taxonomic status. In the present molecular analyses, phylogenetic position of Pterapogon remained unsettled ( Table 5). It was sister to the clade VII ( Siphamiini ) in ML tree ( Fig. 2 View FIGURE 2 ), sister to Ostorhinchus margaritophorus in MP tree-A ( Fig. 4 View FIGURE 4 ), and nested within the clade XII ( Ostorhinchini ) in the MP tree-B ( Fig. 5 View FIGURE 5 ). It formed a polytomy with O. margaritophorus , clade VII, clade VIII, and a large clade including clades IX–XII in BA tree ( Fig. 3 View FIGURE 3 ). Pterapogon has, however, a synapomorphy (spinous procurrent caudal rays) uniquely held with Sphaeramia as well as similarities in color patterns and body shape, but differs with fused hypurals 1+2 and 3+4. Based on these morphological features, we tentatively included this species in the tribe Sphaeramiini . Monophyly of them was not rejected by AU test (H02 in Table 6), although its inclusion within Ostorhinchini was also not rejected (H03 in Table 6). We had no good DNA extracts from tissue of Quinca . This monotypic genus has morphological characteristics and color pattern suggestive of a relationship with Apogonichthyoides , and was provisionally place in the Sphaeramiini .
Tribe Veruluxini new name Fraser & Mabuchi
Type genus Verulux Fraser 1972
Diagnosis. Members of the Apogoninae : dorsal fin VI–I,9; anal fin II,9; head and body with weakly ctenoid or cycloid scales; pored lateral-line scales 24; preopercle ridge smooth, edges smooth; one supraneural; supramaxilla absent; basisphenoid present; uroneurals absent; two epurals; fused hypurals 1+2+3+4, fused to terminal centrum; no autogenous haemal spines; one supernumerary dorsal spine; bioluminescent organ under cleithrum; caudal fin forked.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal fin-ray branched and segmented; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; developed gill rakers 12–15; unbranched procurrent rays, longest segmented; pectoral fin-rays 14–16; 10+14 vertebrae; 8 ribs; 5–6 epipleurals; teeth in one row on premaxilla, dentary, vomer, palatine, all villiform, or absent on palatine; six infraorbitals, bony shelf absent on third infraorbital; stomach and intestine black with silvery peritoneum; low crest on PU2.
Distribution. The single described species is known from East Africa, Red Sea, islands in the Indian Ocean, throughout the West Pacific to Japan, onto the Pacific Plate to Marshall Islands, Vanuatu, New Caledonia and Australia.
Remarks. This tribe contains only one species, Verulux cypselurus , corresponding to the clade V in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). Representatives of Verulux (two individuals from Ryukyu Islands and Seychelles) were monophyletic separated by a moderate genetic distance, which may indicate presence of cryptic species. This lineage was recovered far from Rhabdamia (tribe Rhabdamiini ), although their monophyly (genus Rhabdamia sensu Fraser 1972 ) was not rejected by AU test (H05 in Table 6). The single described species has a bioluminescent organ under cleithrum unlike Rhabdamia .
Tribe Zoramiini new name Fraser & Mabuchi
Type genus Zoramia Jordan 1917
Diagnosis. Members of the Apogoninae : dorsal fin VI–I,9; anal fin II,8–9; head and body with ctenoid scales; pored lateral-line scales 24; preopercle ridge smooth, edges serrate; three supraneurals; supramaxilla absent; basisphenoid present; anterior pair of uroneurals; three epurals; five free hypurals; parhypural separate; two autogenous haemal spines; one supernumerary dorsal spine; caudal fin forked.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal-fin ray branched and segmented; second dorsal spine longest, very long and filamentous in some species of both genera; ctenoid scales on predorsal, cheek, breast, two pelvic scales, and body; ctenoid scales on opercle and onto base of caudal fin; cycloid or ctenoid predorsal scales1–3; pored lateral-line scales simple with one pore above and one below midline; pectoral-fin rays 13–17; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; developed gill rakers 7–38; unbranched procurrent rays, longest segmented; 10+14 vertebrae; 8 ribs; 7–8 epipleurals; teeth in one, two or multiple rows on premaxilla, dentary, vomer, palatine, all villiform; six infraorbitals, bony shelf absent or on third infraorbital; stomach and intestine black with silvery peritoneum; low crest on PU2.
Distribution. Species of Zoramia are known from East Africa, Red Sea, islands in the Indian Ocean, throughout the West Pacific to Japan, onto the Pacific Plate to Marshall Islands, Samoa, Tonga, Solomon Islands, New Caledonia and Australia. Species of Fibramia are known from East Africa, some islands in the Indian Ocean, Sri Lanka, throughout the West Pacific to Philippines, Solomon Islands, Santa Cruz Islands, Fiji, Vanuatu, New Caledonia, Australia, Samoa and Tonga.
Remarks. This tribe contains two genera, a new genus Fibramia described below and Zoramia , corresponding to the clade VIII in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). Both genera have species with elongate second dorsal spine, a single supernumerary dorsal spine, and can be found in tidal fresh and brackish water, near shore waters and in lagoons. Fibramia corresponds to the clade of Ostorhinchus - 4 in the present study, and that of Ostorhinchus I in Mabuchi et al. (2006).
Tribe Veruluxini new name Fraser & Mabuchi
Type genus Verulux Fraser 1972
Diagnosis. Members of the Apogoninae : dorsal fin VI–I,9; anal fin II,9; head and body with weakly ctenoid or cycloid scales; pored lateral-line scales 24; preopercle ridge smooth, edges smooth; one supraneural; supramaxilla absent; basisphenoid present; uroneurals absent; two epurals; fused hypurals 1+2+3+4, fused to terminal centrum; no autogenous haemal spines; one supernumerary dorsal spine; bioluminescent organ under cleithrum; caudal fin forked.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal fin-ray branched and segmented; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; developed gill rakers 12–15; unbranched procurrent rays, longest segmented; pectoral fin-rays 14–16; 10+14 vertebrae; 8 ribs; 5–6 epipleurals; teeth in one row on premaxilla, dentary, vomer, palatine, all villiform, or absent on palatine; six infraorbitals, bony shelf absent on third infraorbital; stomach and intestine black with silvery peritoneum; low crest on PU2.
Distribution. The single described species is known from East Africa, Red Sea, islands in the Indian Ocean, throughout the West Pacific to Japan, onto the Pacific Plate to Marshall Islands, Vanuatu, New Caledonia and Australia.
Remarks. This tribe contains only one species, Verulux cypselurus , corresponding to the clade V in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). Representatives of Verulux (two individuals from Ryukyu Islands and Seychelles) were monophyletic separated by a moderate genetic distance, which may indicate presence of cryptic species. This lineage was recovered far from Rhabdamia (tribe Rhabdamiini ), although their monophyly (genus Rhabdamia sensu Fraser 1972 ) was not rejected by AU test (H05 in Table 6). The single described species has a bioluminescent organ under cleithrum unlike Rhabdamia .
Tribe Zoramiini new name Fraser & Mabuchi
Type genus Zoramia Jordan 1917
Diagnosis. Members of the Apogoninae : dorsal fin VI–I,9; anal fin II,8–9; head and body with ctenoid scales; pored lateral-line scales 24; preopercle ridge smooth, edges serrate; three supraneurals; supramaxilla absent; basisphenoid present; anterior pair of uroneurals; three epurals; five free hypurals; parhypural separate; two autogenous haemal spines; one supernumerary dorsal spine; caudal fin forked.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal-fin ray branched and segmented; second dorsal spine longest, very long and filamentous in some species of both genera; ctenoid scales on predorsal, cheek, breast, two pelvic scales, and body; ctenoid scales on opercle and onto base of caudal fin; cycloid or ctenoid predorsal scales1–3; pored lateral-line scales simple with one pore above and one below midline; pectoral-fin rays 13–17; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; developed gill rakers 7–38; unbranched procurrent rays, longest segmented; 10+14 vertebrae; 8 ribs; 7–8 epipleurals; teeth in one, two or multiple rows on premaxilla, dentary, vomer, palatine, all villiform; six infraorbitals, bony shelf absent or on third infraorbital; stomach and intestine black with silvery peritoneum; low crest on PU2.
Distribution. Species of Zoramia are known from East Africa, Red Sea, islands in the Indian Ocean, throughout the West Pacific to Japan, onto the Pacific Plate to Marshall Islands, Samoa, Tonga, Solomon Islands, New Caledonia and Australia. Species of Fibramia are known from East Africa, some islands in the Indian Ocean, Sri Lanka, throughout the West Pacific to Philippines, Solomon Islands, Santa Cruz Islands, Fiji, Vanuatu, New Caledonia, Australia, Samoa and Tonga.
Remarks. This tribe contains two genera, a new genus Fibramia described below and Zoramia , corresponding to the clade VIII in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). Both genera have species with elongate second dorsal spine, a single supernumerary dorsal spine, and can be found in tidal fresh and brackish water, near shore waters and in lagoons. Fibramia corresponds to the clade of Ostorhinchus - 4 in the present study, and that of Ostorhinchus I in Mabuchi et al. (2006).
Type genus Zoramia Jordan 1917
Diagnosis. Members of the Apogoninae : dorsal fin VI–I,9; anal fin II,8–9; head and body with ctenoid scales; pored lateral-line scales 24; preopercle ridge smooth, edges serrate; three supraneurals; supramaxilla absent; basisphenoid present; anterior pair of uroneurals; three epurals; five free hypurals; parhypural separate; two autogenous haemal spines; one supernumerary dorsal spine; caudal fin forked.
Other characteristics. first segmented fin-ray in second dorsal-fin branched, first anal-fin ray branched and segmented; second dorsal spine longest, very long and filamentous in some species of both genera; ctenoid scales on predorsal, cheek, breast, two pelvic scales, and body; ctenoid scales on opercle and onto base of caudal fin; cycloid or ctenoid predorsal scales1–3; pored lateral-line scales simple with one pore above and one below midline; pectoral-fin rays 13–17; 9+8 segmented principal caudal rays, 15 branched, upper and lower unbranched; developed gill rakers 7–38; unbranched procurrent rays, longest segmented; 10+14 vertebrae; 8 ribs; 7–8 epipleurals; teeth in one, two or multiple rows on premaxilla, dentary, vomer, palatine, all villiform; six infraorbitals, bony shelf absent or on third infraorbital; stomach and intestine black with silvery peritoneum; low crest on PU2.
Distribution. Species of Zoramia are known from East Africa, Red Sea, islands in the Indian Ocean, throughout the West Pacific to Japan, onto the Pacific Plate to Marshall Islands, Samoa, Tonga, Solomon Islands, New Caledonia and Australia. Species of Fibramia are known from East Africa, some islands in the Indian Ocean, Sri Lanka, throughout the West Pacific to Philippines, Solomon Islands, Santa Cruz Islands, Fiji, Vanuatu, New Caledonia, Australia, Samoa and Tonga.
Remarks. This tribe contains two genera, a new genus Fibramia described below and Zoramia , corresponding to the clade VIII in the molecular trees ( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 4). Both genera have species with elongate second dorsal spine, a single supernumerary dorsal spine, and can be found in tidal fresh and brackish water, near shore waters and in lagoons. Fibramia corresponds to the clade of Ostorhinchus - 4 in the present study, and that of Ostorhinchus I in Mabuchi et al. (2006).
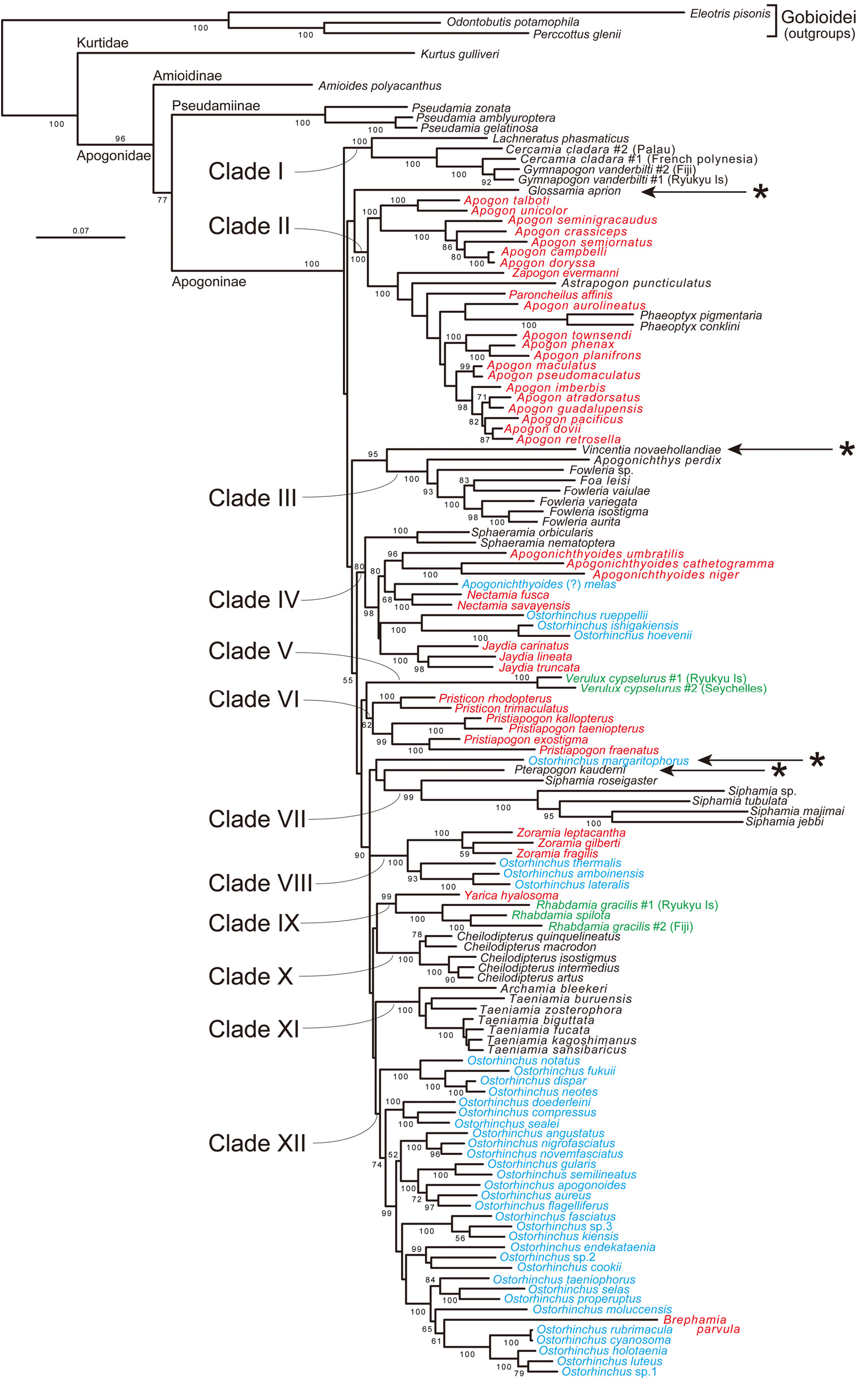
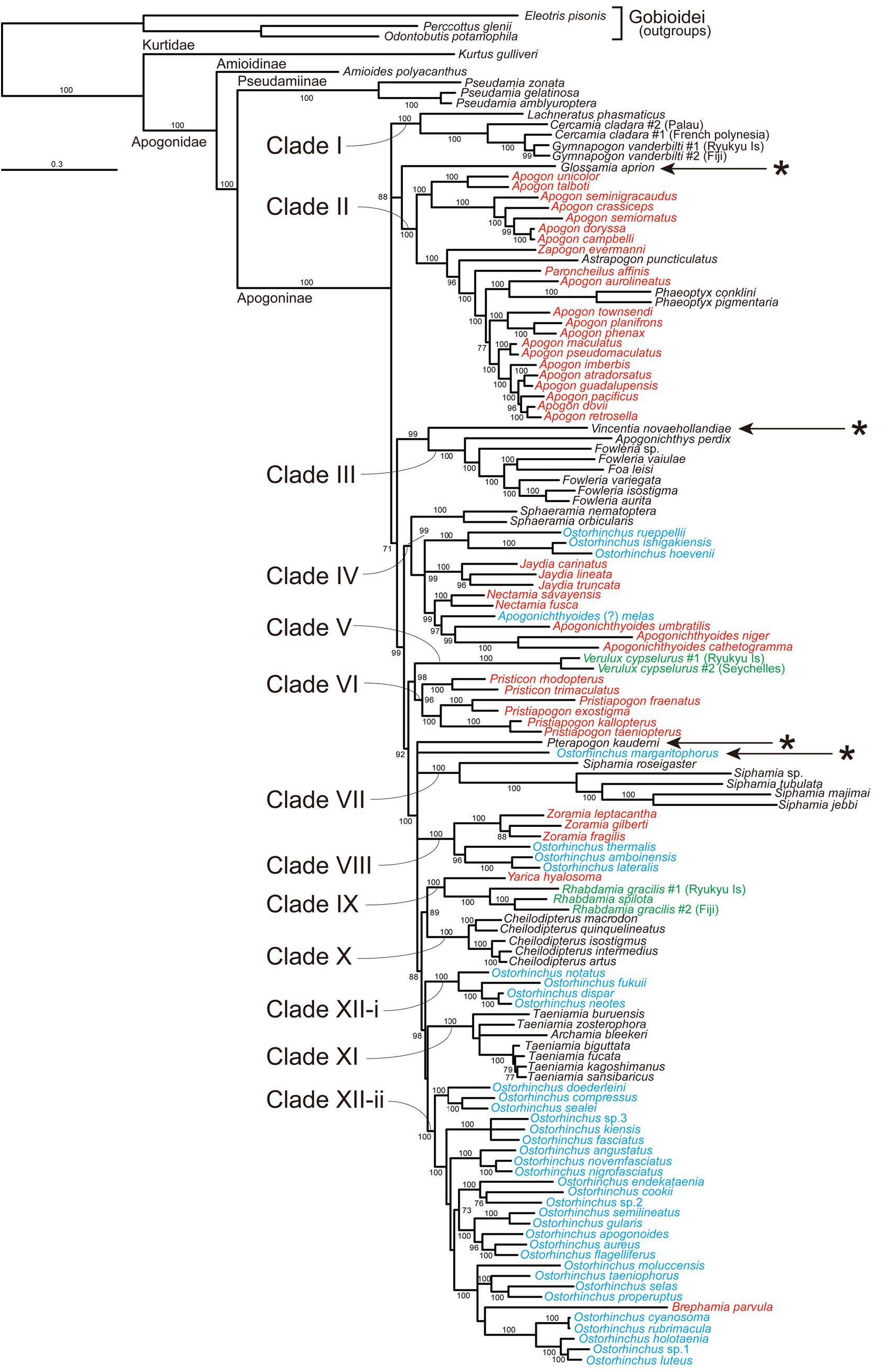
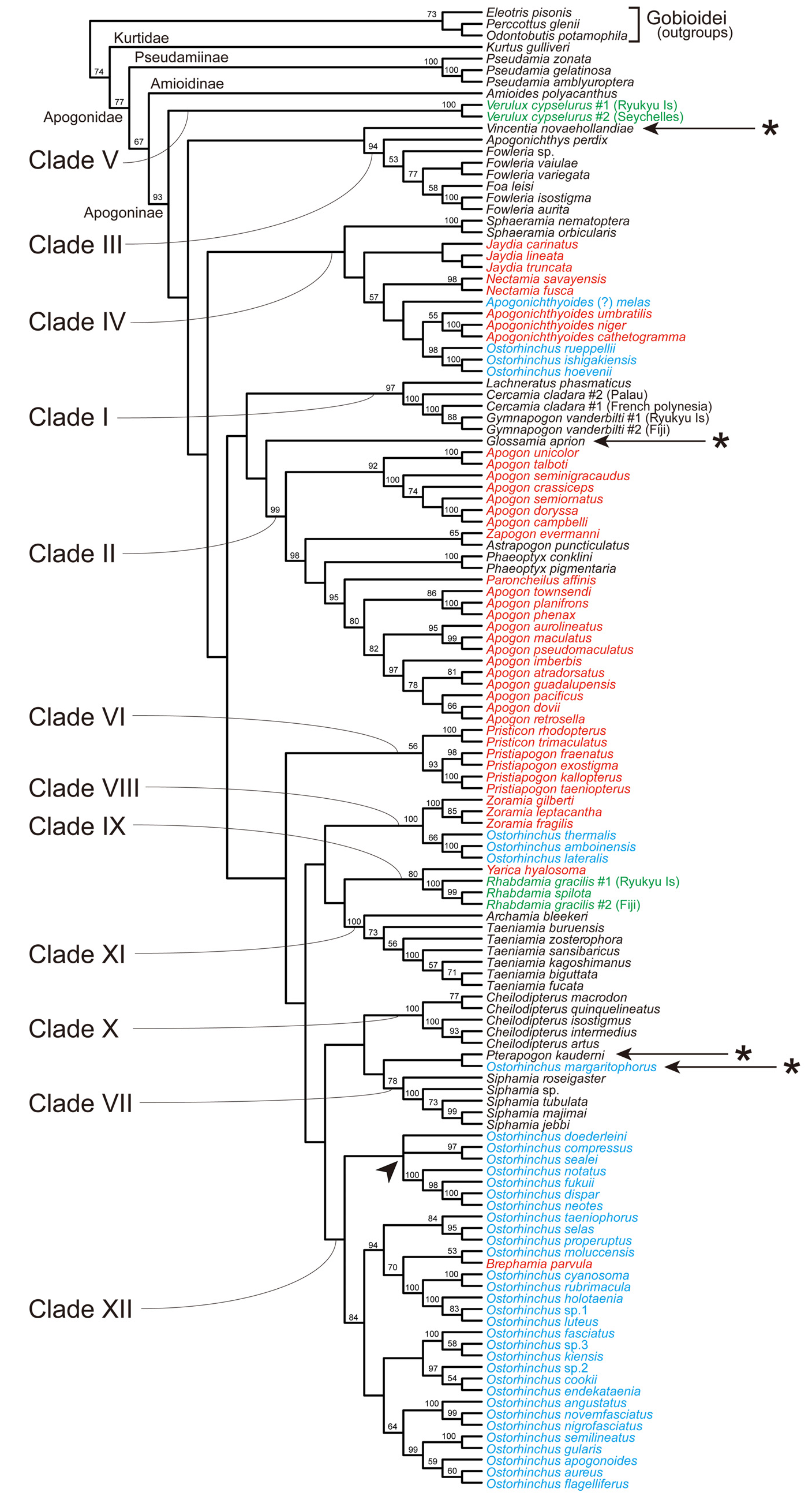
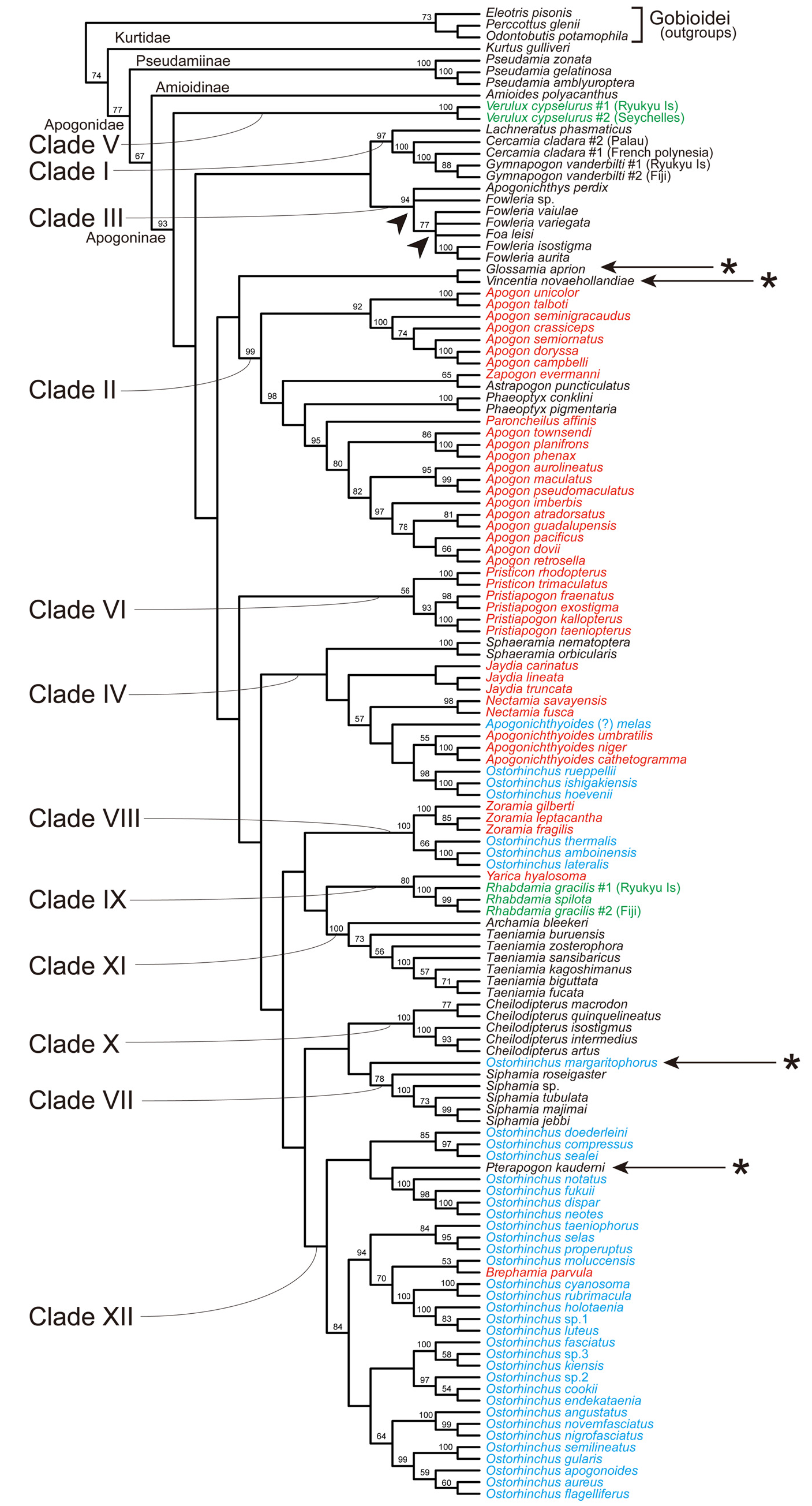
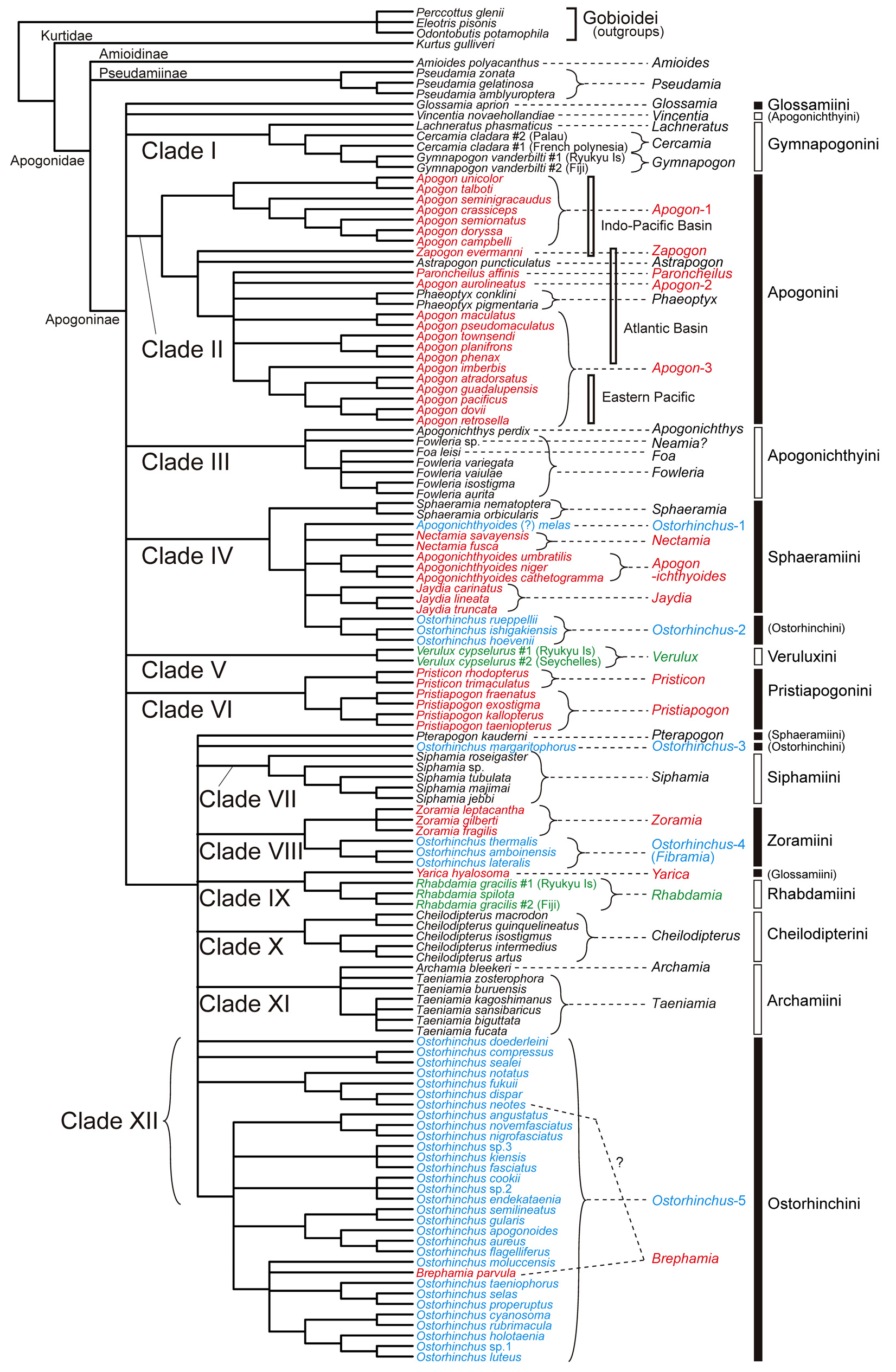

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.