
Trimeresurus kraensis, Idiiatullina & Suwannapoom & Tawan & Chanhome & Nguyen & David & Vogel & Poyarkov, 2024
|
publication ID |
https://doi.org/ 10.5852/ejt.2024.930.2489 |
|
publication LSID |
lsid:zoobank.org:pub:C9615291-8DBB-459E-A420-1AAA5DEF625A |
|
DOI |
https://doi.org/10.5281/zenodo.10903323 |
|
persistent identifier |
https://treatment.plazi.org/id/AF38FFC5-F4E0-48CE-A897-8C350982625F |
|
taxon LSID |
lsid:zoobank.org:act:AF38FFC5-F4E0-48CE-A897-8C350982625F |
|
treatment provided by |
Plazi |
|
scientific name |
Trimeresurus kraensis |
| status |
sp. nov. |
Trimeresurus kraensis sp. nov.
urn:lsid:zoobank.org:act:AF38FFC5-F4E0-48CE-A897-8C350982625F
Figs 2–4 View Fig View Fig View Fig , 5C View Fig , 6A–B View Fig ; Tables 3–4 View Table 3 View Table 4
Trimeresurus venustus View in CoL (partim) – Pauwels et al. 2013: 280. — Sumontha et al. 2021: 320.
Trimeresurus cf. venustus View in CoL – Sumontha et al. 2021: 321, fig. 9B.
Trimeresurus cf. venustus View in CoL 1 – Idiiatullina et al. 2023: 699, 704.
Diagnosis
The new species is assigned to the subgenus Trimeresurus based on the following morphological attributes: a long papillose hemipenis and partially fused first supralabial and nasal scales ( Malhotra & Thorpe 2004a; Idiiatullina et al. 2023). The new species Trimeresurus kraensis sp. nov. is distinguished from all other species of the subgenus Trimeresurus by the following combination of morphological characters: (1) a dark olive-green or bottle green dorsum; (2) dorsal pattern consisting of about 60 reddish-brown or purple blotches, transversally elongate but not reaching the lower part of the flanks, two or three series of dark brown spots forming a discontinuous pattern on the 1 st to 3 rd dorsal scale rows and white vertebral spots present in males, located approximately on every two or four dorsal scales; (3) venter creamish-green with some dark brown spots; (4) tips of the ventral plates both cream and dark brown, forming a discontinuous, alternating pale and dark, ventrolateral stripe; (5) males with a reddish-brown postocular stripe; (6) internasals generally in contact behind the posteror tip of the rostral; (7) single large supraocular scale; (8) iris pale copper, (9) tail brown with dark purplish-brown crossbars; (10) dorsal scales in 21–21–15 rows; (11) ventral scales 167 in a single male, 169–171 in females; (12) subcaudal scales 62 in a single male, 52–54 in females, all paired.
Etymology
The species name is the modern Latin adjective ʻ kraensis ʼ in the nominative singular, masculine gender, combining the noun ʻKraʼ, from the name of the Kra Isthmus in Peninsular Thailand where the type locality of the species is located, and the Latin suffix ʻ- ensis ʼ (- is, - e), meaning ʻfromʼ. The species name therefore means, ʻfrom Kraʼ. We suggest the following common names for the new species: งูหางแฮ่มชุมพร (Ngu Hang Ham Chumphorn) (in Thai), and Kra Isthmus Pitviper (English).
Type material
Holotype
THAILAND • ♀ adult; Chumphon Province, Muang District, Banna Subdistrict, Wat [= Temple] Tham Sanook and its cave Tham Sanook ; 10.48089° N, 99.07323° E; 65 m a.s.l.; 27 Jul. 2022; P. Pawangkhanant, N.A. Poyarkov and C. Suwannapoom leg.; AUP-02036 . GoogleMaps
Paratypes
THAILAND • 1 ♂ adult; same collection data as for holotype; 27 Jul. 2022; ZMMU Re-17664 (field label NAP-11581) • 1 ♀ adult; same collection data as for holotype; 27 Jul. 2022; ZMMU Re-17666 (field label NAP-11582) • 1 ♀ subadult; same collection data as for holotype; 15 Jan. 2023; P. Pawangkhanant and S.S. Idiiatullina leg.; ZMMU Re-17665 (field label ISS-004) .
Description (holotype)
Adult female specimen ( Fig. 3 View Fig ) in good state of preservation; measurements of holotype presented in Table 4 View Table 4 . Body cylindrical, long and thin ( SVL 459 mm, TaL 83 mm, TL 542 mm, TaL/TL 0.153) ( Fig. 3 View Fig ). Head triangular in dorsal view ( Fig. 3E View Fig ), elongate, clearly distinct from neck (HL 21.7 mm, HW 17.3 mm, HW/HL 0.80). Snout elongate, flattened, and rounded in dorsal view ( Fig. 3E View Fig ), rather rectangular in lateral view ( Fig. 3D View Fig ), with very distinct and sharp canthus rostralis (SnL/HL 0.31, SnL/ED 2.31). Rostral barely visible in dorsal aspect, triangular ( Fig. 3E View Fig ). One pair of enlarged internasals, in contact with each other behind posteror tip of rostral ( Fig. 3E View Fig ). Pupil vertically elliptical, loreal pit present, and triangular in shape ( Fig. 3D View Fig ). Nostril completely enclosed in entire nasal scale, nasal scale partially fused with first supralabial ( Fig. 3D View Fig ). Shield bordering anterior edge of loreal pit fused with second supralabial ( Fig. 3D View Fig ). One subocular long, thin, crescent-like, separated from 4 th and 5 th supralabials by one row of scales; single posterior subocular small ( Fig. 3D View Fig ). Three preoculars on each side of head; two upper preoculars located above loreal pit, elongated, in contact with single scale formed by fused second supralabial and loreal, which separates them from nasal; lower preoculars elongated, rectangular, forming lower margin of loreal pit, lower preocular in contact with third supralabial ( Fig. 3D View Fig ). Supralabials 12/12, fourth one largest ( Fig. 3D View Fig ); 12/12 infralabials, those of first pair in contact with each other behind mental ( Fig. 3F View Fig ); first three pairs of infralabials in contact with single pair of chin shields ( Fig. 3F View Fig ). Seven pairs of gulars aligned between chin shields and single preventral ( Fig. 3F View Fig ). One supraocular on each side, enlarged, slightly indented on its inner margin by upper head scales ( Fig. 3E View Fig ). Scales on snout and in interorbital region smooth, irregular, subimbricate; temporal and occipital scales moderately keeled ( Fig. 3D–E View Fig ). Dorsal scales in 21–21–15 rows. Dorsal scales all keeled, except first row, which is smooth. Single preventral + 171 ventrals. Cloacal plate single; 54 subcaudals, all divided.
Colouration of holotype in life ( Fig. 3 View Fig )
The background dorsal colour is bright olive-green above, slightly paler bluish-green on the lower part of each side; many dorsal scales heavily stippled with minute reddish-brown dots, especially on the lower part of the sides; a dorsal series of about 60 dorsal blotches, dark reddish-brown, transversally elongate, rather short, wider than long, not extending downwards on the flanks beyond the 5 th dorsal scale row and not forming true crossbands; these dorsal blotches are much irregular in shape, often constricted in their middle or partially divided into two lateral blotches, united or alternating with one another along the vertebral row; white, elongate narrow spots or dashes aligned on the vertebral row, located approximately every two or four dorsal scales; one or two series of dark brown spots, about one-scale long, scattered on the 1 st to 3 rd dorsal scale rows, irregularly spaced, usually forming pair obliquely arranged; some scales of the 1 st dorsal row cream as the tips of dorsals. The tail is coloured and patterned like the dorsum, with about 15 dorsal blotches, turning to dark brown on a pink background on the posterior quarter of the tail. The head is bright olive-green like the body above, largely covered with dark reddish-brown areas, of the same colour as the dorsal blotches, so extensively that the head appears reddish-brown above with narrow olive-green lines; rostral olive-green on its lower part, dark reddish-brown on its upper part; internasals and the canthal scales more extensively green than reddish-brown; side of the head pale green; paler than the upper head surface; nasal, loreal and preoculars with dark reddish-brown spots or blotches; supralabials dotted with pale blue; two broad vertical streaks, dark reddish brown covering the parts of the supralabials located below the loreal pit and the eye, respectively; a broad, dark reddish-brown postocular streak, edged above with a narrow area of bright olive-green, extends from the eye to the corner of the mouth. The infralabials, chin and throat are cream with dark brown spots, more numerous on the mental scale and the supralabials.
The venter is pale creamish-green, mostly uniform but with some scattered dark brown spots; tips and outer parts of most ventral plates both cream and dark brown, forming a discontinuous, pale and dark, ventrolateral stripe extending from the neck to the vent. The ventral surface of the tail is cream heavily blotched with dark brown, becoming entirely dark brown posteriorly.
Colouration of holotype in preservative
In preservative, the background dorsal colour fades to dark grey, but the pattern remains generally unchanged.
Variation
The main meristic and morphometric characters of the type series of Trimeresurus kraensis sp. nov. are summarized in Table 4 View Table 4 ; photographs of paratype female ZMMU Re-17665 and an uncollected specimen are presented in Fig. 4C View Fig and Fig. 4B View Fig , respectively. The holotype AUP-02036 represents the longest known specimen and is 542 mm long. The longest known male is 397 mm long (SVL 328 mm, TaL 69 mm, ZMMU Re-17664). Ratio TaL/TL: 0.138 –0.174 (male: 0.174, females: 0.138 –0.153). Below, we will summarize the variation among the sexes in several main morphological characters.
Body scalation
21–21–15 DSR; 167–171 VEN (male: 167, females: 169–171); 52–62 SC (male: 62, females: 52–53); total number of VEN + SC: 222–229 (male: 229, females: 222–225).
Head scalation
The internasals in contact in all examined specimens but in the male paratype ZMMU Re-17664 they are separated by a small scale; SL: 9–12; IL: 10–13.
Main characters of colouration pattern
Reddish-brown dorsal blotches not forming true crossbands, not extending downwards below the 5 th dorsal scale rows; postocular streak always present, dark reddish-brown, broad and conspicuous in both males and females; white ventrolateral stripe present in both sexes.
Distribution and natural history
Currently, Trimeresurus kraensis sp. nov. is known only from two small karst formations, namely the Sanook Cave (environs of Wat Tham Sanook Monastery ; Fig. 4A View Fig ) and Chang Phueak Cave (environs of Wat Tham Chang Phueak Monastery), both in Banna Subdistrict, Muang District, Chumphon Province, Thailand (approximate coordinates: 10.446216° N, 99.035114° E; 87 m a.s.l.; Sumontha et al. 2021; our data). The two localities are separated by a distance of 5.8 km from each other and are located on the opposite sides of a low limestone hilly area spanning from north-east to south-west for ca 22 km in the central part of the Chumphon Province of Thailand. It is highly likely that Trimeresurus kraensis sp. nov. can be found at additional localities along this karst area; nevertheless, the estimated distribution range of the new species seems to be restricted to this limestone landscape.
The new species appears to be a locally abundant species of snake. Adult individuals were recorded after dusk (18:30–22:00) while foraging on large karst rocks and walls ( Fig. 4A View Fig ). Young snakes were generally found while perching on small bushes growing among the limestone rocks; individuals in pre-shedding phase were also recorded while perching on bushes. Trimeresurus kraensis sp. nov. is quite aggressive when handled. Toxicological status of the new species remains unknown. Nothing is known about the diet of the new species in the wild either, but it is most likely a generalist predator, as captive individuals ate geckos, small frogs and mice, similar to T. kuiburi as described by Sumontha et al. (2021). Reproduction of the new species was not observed.
At the type locality (the Sanook Cave ), the habitat of the new species represents tropical secondary forest with numerous lianas and bushes covering the limestone formations ( Fig. 4A View Fig ). The new species was found in syntopy with snake species Ahaetulla prasina Boie, 1827 and Elaphe taeniura Cope, 1861 ( Colubridae Oppel, 1811 ), and Trimeresurus cf. albolabris Gray, 1842 ( Viperidae ), as well as with gecko species Cyrtodactylus sanook Pauwels, Sumontha, Latinne & Grismer, 2013 , C. cf. oldhami (Theobald, 1876) , Gehyra lacerata (Taylor, 1962) , Dixonius siamensis (Boulenger, 1899) , and Gekko gecko (Linnaeus, 1758) ( Gekkonidae Oppel, 1811 ) (see Pauwels et al. 2013; our data). The syntopic species of amphibians included: Microhyla tetrix Poyarkov, Pawangkhanant, Gorin, Juthong & Suwannapoom, 2020 , M. butleri Boulenger, 1900 , M. heymonsi Vogt, 1911 , M. mukhlesuri Hasan, Islam, Kuramoto, Kurabayashi & Sumida, 2014 , Micryletta cf. erythropoda (Tarkhnishvili, 1994) , and Glyphoglossus guttulatus (Blyth, 1856) ( Microhylidae Günther, 1858 (1843)); Kurixalus sp. ( Rhacophoridae Hoffman, 1932 (1858)); Occidozyga martensii (Peters, 1867) and Limnonectes hascheanus (Stoliczka, 1870) ( Dicroglossidae Anderson, 1871 ); and Ingerophrynus parvus (Boulenger, 1887) ( Bufonidae Gray, 1825 ).
Comparisons
The new species is morphologically and phylogenetically placed within the subgenus Trimeresurus ( Malhotra & Thorpe 2004a; David et al. 2011) and is morphologically overall most similar to other limestone-dwelling species of pitvipers belonging to the T. kanburiensis complex, including: T. ciliaris , T. kanburiensis , T. kuiburi , and T. venustus s. str.; therefore, the comparisons of the new species with these four congeners appear to be the most pertinent. The main diagnostic characters separating Trimeresurus kraensis sp. nov. from these four species are summarized in Table 3 View Table 3 , additional morphological data are presented in Appendix 1. The comparison of body colouration and head scalation of these species is presented in Figs 5 View Fig and 6 View Fig , respectively.
Trimeresurus kraensis sp. nov. differs from T. ciliaris by having lower total length in males (max TL 397 mm vs 432 mm); lower number of ventrals in males (VEN 167 vs 172–175 [avg. 173]); slightly lower number of ventrals plus subcaudals in males (VEN+SC 229 vs 232–238 [avg. 234.0]); higher number of anterior dorsal scale rows (ASR 21 vs 17); higher number of midbody scale rows (MSR 21 vs 17); postocular stripe straight (vs concave); small scale between nasal and 2 nd supralabial absent (vs present); single large supraocular scale (vs three or four small supraocular scales); white vertebral dots absent in females (vs present, every 2–3 scales); ventral surface pale creamish-green (vs creamy white); pale and dark ventrolateral stripe always present, discontinuous, reddish-brown (vs absent); and iris pale copper (vs olive-green with faded brown horizontal stripe).
The new species differs from T. kanburiensis by having lower total length in both sexes (max TL 397 mm in male, 542 mm in female vs 500 mm in male, 667 mm in female, respectively); slightly lower number of ventrals in males (VEN 167 vs 172); slightly higher number of subcaudals in females (SC 52–54 [avg. 53.0] vs 46–51 [avg. 48.3]); slightly lower anterior number of dorsal scale rows (ASR 21 vs 21–23 [avg. 22.2]); higher number of midbody scale rows (MSR 21 vs 19); higher number of interorbital scales (IOS 11–13 vs 7–9); internasals generally in contact (vs always separated); ventral surface pale creamish-green (vs creamy white).
Trimeresurus kraensis sp. nov. further differs from T. kuiburi by having lower total length in males (max TL 397 mm vs 465 mm); higher number of midbody scale rows (MSR 21 vs 19); postocular streak straight (vs concave); white vertebral dots present on every 2–4 scales in males (vs every 5–6 scales); and pale and dark lateral stripe on ventrals always present, discontinuous, cream, and reddish-brown (vs absent).
Finally, Trimeresurus kraensis sp. nov. can be differentiated from T. venustus s. str. by having lower total length in both sexes (max TL 397 mm in male, 542 mm in female vs 580 mm in male, 530 mm in female, respectively); slightly higher number of ventrals in females (VEN 169–171 [avg. 170.3] vs 173–181 [avg. 177.3]); slightly lower number of ventrals plus subcaudals in both sexes (VEN+SC 229 in male, 222–225 [avg. 223.3] in females vs 229–248 [avg. 238.7] in males, 224–236 [avg. 231.5] in females); internasals always in contact (vs always separated); white vertebral dots present every 2–4 scales in males (vs absent); dorsal pattern consisting of transversally elongate blotches, not extending downwards beyond the 5 th dorsal scale row (vs true crossbands, narrow and reaching the 1 st dorsal scale row).
Among the other species of the subgenus Trimeresurus , the new species can be readily distinguished from T. albolabris Gray, 1842 , T. andersonii Theobald, 1868 , T. cantori (Blyth, 1846) , T. cardamomensis (Malhotra, Thorpe, Mrinalini & Stuart, 2011) , T. caudornatus Chen, Ding, Vogel & Shi, 2020 , T. davidi Chandramouli, Campbell & Vogel, 2020 , T. erythrurus (Cantor, 1839) , T. fasciatus (Boulenger, 1896) , T. guoi Chen, Shi, Vogel & Ding, 2021 , T. honsonensis ( Grismer, Ngo & Grismer, 2008) , T. insularis Kramer, 1977 , T. labialis (Fitzinger in Steindachner, 1867), T. macrops Kramer, 1977 , T. mutabilis Stoliczka, 1870 , T. purpureomaculatus (Gray, 1832) , T. rubeus (Malhotra, Thorpe, Mrinalini & Stuart, 2011) , T. salazar Mirza, Bhosale, Phansalkar, Sawant, Gowande & Patel, 2020 , and T. septentrionalis Kramer, 1977 by dorsal colouration and pattern (reddish-brown or purple dorsal blotches on olive-green or bottle-green background in the new species vs uniform green or green colouration with no pattern or pattern consisting of small brownish spots or speckles in other species, or straw-yellow background with irregular, dark-brown transverse body bands in T. honsonensis ). Moreover, by having 21 MSR, the new species can be further separated from T. cantori (27, 29 or 31 MSR), T. erythrurus (23 rarely 21, 25 MSR), and T. purpureomaculatus (25 rarely 27, 29 MSR) (see Gumprecht et al. 2004; Grismer et al. 2008; Malhotra et al. 2011; Chandramouli et al. 2020; Chen et al. 2020, 2021; Mirza et al. 2020; our data).
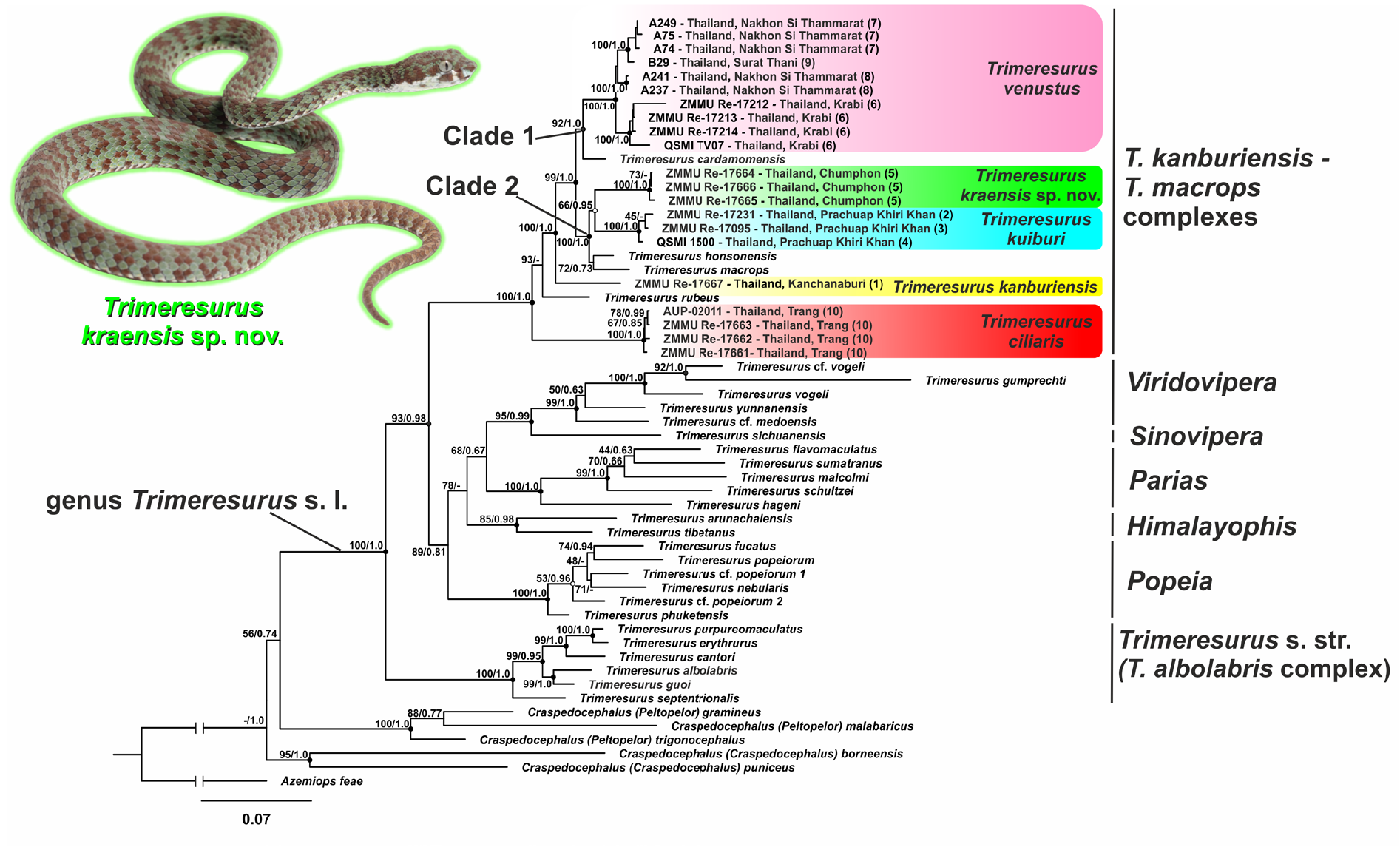
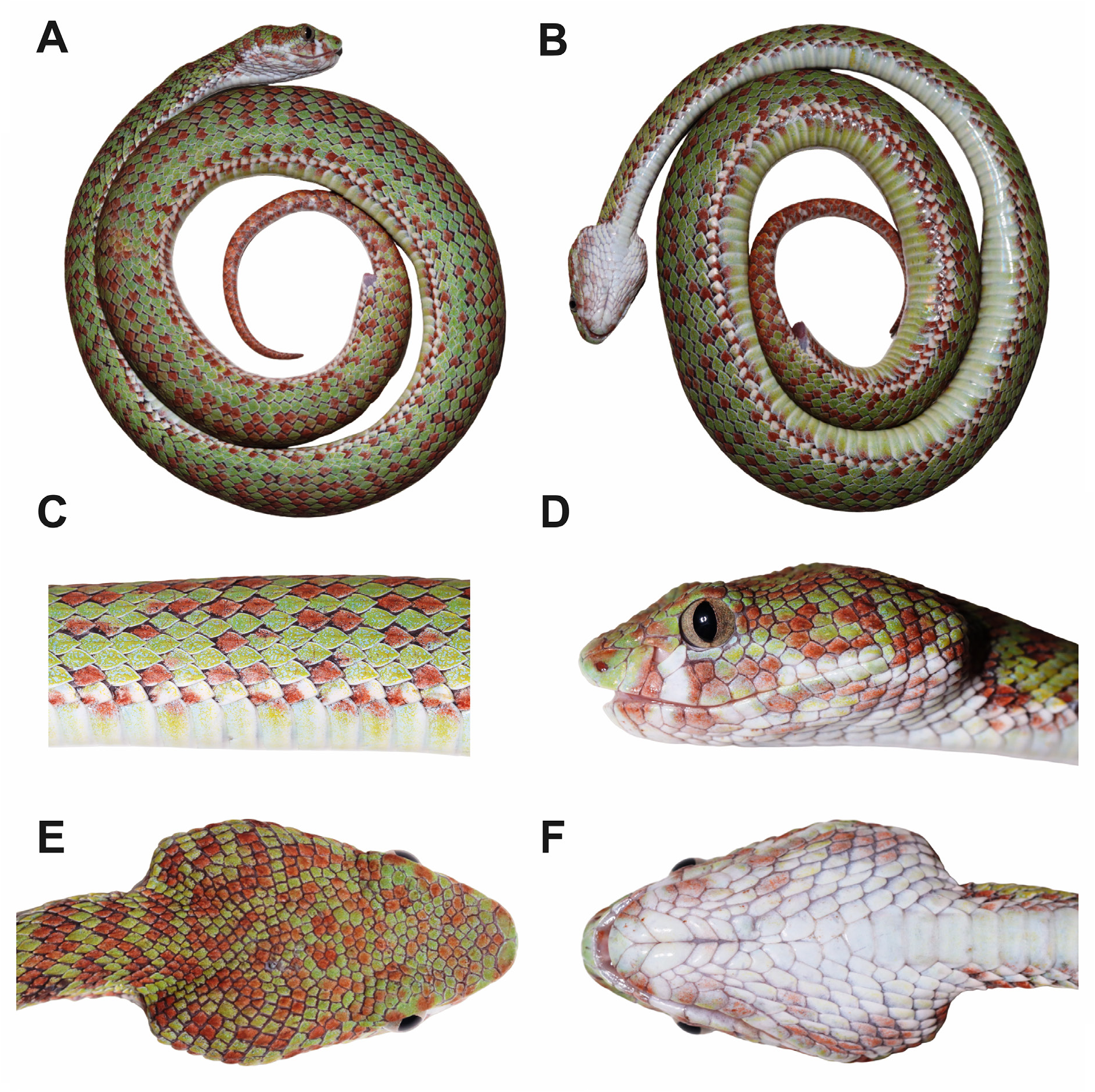



Table 4. Morphological data on the type series of Trimeresurus kraensis sp. nov. For abbreviations see Material and methods section.
| Specimen ID | AUP-02036 | ZMMU Re-17664 | ZMMU Re-17665 | ZMMU Re-17666 |
|---|---|---|---|---|
| Type status | Holotype | Paratype | Paratype | Paratype |
| Sex | F | M | F | F |
| SVL (mm) | 459 | 328 | 270 | 263 |
| TaL (mm) | 83 | 69 | 47 | 42 |
| TL (mm) | 542 | 397 | 317 | 305 |
| HL (mm) | 21.7 | 16.2 | 15.8 | 14.8 |
| HW (mm) | 17.3 | 11.5 | 11 | 10.5 |
| HD (mm) | 10.4 | 7.4 | 8.1 | 6.7 |
| SnL (mm) | 6.7 | 4.4 | 4.3 | 4 |
| ED (mm) | 2.9 | 3.1 | 2.9 | 2.9 |
| VED (mm) | 2.4 | 2.2 | 2.1 | 2.1 |
| SOL (mm) | 3.5 | 2.9 | 3.5 | 2.9 |
| SOW (mm) | 1.3 | 1.1 | 1.2 | 1.1 |
| Internasals in contact | yes | no | yes | yes |
| Contact nasal–1 st SL | partially fused | partially fused | partially fused | partially fused |
| IOS | 11 | 11 | 13 | 11 |
| PosOc | 2/2 | 2/2 | 2/2 | 2/2 |
| SL | 12/12 | 9/9 | 10/9 | 9/9 |
| IL | 12/12 | 11/10 | 12/13 | 11/11 |
| ASR | 21 | 21 | 21 | 21 |
| MSR | 21 | 21 | 21 | 21 |
| PSR | 15 | 15 | 15 | 15 |
| SRR 21 to 19 (V) | 112 | 112 | 111 | 108 |
| SRR 19 to 17 (V) | 122 | 122 | 121–122 | 117 |
| SRR 17 to 15 (V) | 131 | 131-132 | 131 | 125 |
| PreV | 1 | 2 | 2 | 1 |
| VEN | 171 | 167 | 169 | 171 |
| CP | 1 | 1 | 1 | 1 |
| SC | 54 | 62 | 53 | 52 |
| ZMMU |
Zoological Museum, Moscow Lomonosov State University |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SubOrder |
Serpentes |
|
Family |
|
|
SubFamily |
Crotalinae |
|
Genus |
Trimeresurus kraensis
| Idiiatullina, Sabira S., Pawangkhanant, Parinya, Suwannapoom, Chatmongkon, Tawan, Tanapong, Chanhome, Lawan, Nguyen, Tan Van, David, Patrick, Vogel, Gernot & Poyarkov, Nikolay A. 2024 |
Trimeresurus cf. venustus
| Idiiatullina S. & Pawangkhanant P. & Tawan T. & Worranuch T. & Dechochai B. & Suwannapoom C. & Nguyen T. V. & Chanhome K. & Poyarkov N. A. 2023: 699 |
Trimeresurus cf. venustus
| Sumontha et al. 2021: 321 |
Trimeresurus venustus
| Sumontha et al. 2021: 320 |
| Pauwels O. S. G. & Sumontha M. & Latinne A. & Grismer L. L. 2013: 280 |