
Crotaphopeltis hippocrepis (Reinhardt, 1843)
|
publication ID |
https://doi.org/10.5281/zenodo.5394650 |
|
persistent identifier |
https://treatment.plazi.org/id/BB6887E1-FF94-7362-52AE-FD8EFC0CFB65 |
|
treatment provided by |
Marcus |
|
scientific name |
Crotaphopeltis hippocrepis (Reinhardt, 1843) |
| status |
|
Crotaphopeltis hippocrepis (Reinhardt, 1843)
Dipsas hippocrepis Reinhardt, 1843: 26 .
HOLOTYPE. — 1 juvenile female from the Danish settlements on the coast of Guinea, near Accra, Ghana ( ZMUC R63127).
Heterurus hippocrepis – Duméril et al. 1854: 1177.
Leptodeira hippocrepis – Gray 1858: 161.
Leptodira hotamboeia (not Laurenti) – Boulenger 1896: 90 (synonymised D. hippocrepis with L. hotamboeia ).
Crotaphopeltis hotamboeia (not Laurenti) – Chabanaud 1921: 469 (part). – Balletto et al. 1973: 92 (part). – Hughes 1983: 348 (part). – Joger 1990: 95 (part).
Crotaphopeltis hotamboeia hotamboeia (not Laurenti) – Angel 1933: 148 (part). – Monard 1951: 163 (part).
Crotaphopeltis hotemboeia hotemboeia [sic] – Roux- Estève 1969: 114 (part).
Crotaphopeltis acarina Roman, 1974: 6 ; 1980: 53; 1984: 39. – Rasmussen 1979: 99. – Joger 1982: 333. – Welch 1982: 147.
HOLOTYPE. — H-V 4780, 1 adult specimen from Tionkuy, Burkina-Faso.
Crotaphopeltis hippocrepis – Rasmussen 1985: 126. – Böhme & Schneider 1987: 259. – Hughes 1988: 34. – Rödel et al. 1995: 8. – Rasmussen & Hughes 1997: 22.
DIAGNOSIS
A West and Central African savanna species of Crotaphopeltis with the following character combination: 19 scale rows at mid-body, scales in more or less oblique rows anteriorly, smooth all over the body; 12-16 + II + 1 maxillary teeth; 44-58 (male) and 39-54 (female) subcaudals; hemipenis extending to subcaudal scute No. 8-12 and provided with two enlarged spines proximally; dorsum various shades of brown, without white specks; occiput white in juveniles; temples with a dark brown mark extending to posterior jaw, covering the last two to five infralabials with pigment; venter and underside of tail white, cream or yellowish, latter often with median stripe.
DESCRIPTION
Rostral 1.5-2.2 times as broad as deep, usually in broad contact with internasals; frontal 1.2-1.7 time as long as broad ( Table 1), and 1.0-1.6 time as long as its distance from end of snout, and 0.9- 1.5 time as long as suture between parietals; loreal usually one (two on one side in three specimens), normally deeper than long; preocular usually one ( Table 1), in one case in contact with frontal; postoculars usually two (for variation see Table 1); temporals usually 1 + 2 (for variation see Table 1); supralabials usually eight, third to fifth usually entering orbit (for variation see Table 1); infralabials usually ten, first five on each side usually in contact with an anterior chinshield (for various combinations see Table 1); chin-shields and enlarged median gulars in three to six pairs followed by zero to two preventrals.
Dorsal scales often arranged in oblique rows anteriorly, smooth with well-defined single, rarely double, apical pits, the latter being mainly confined to the flanks. Scale row reduction formula (body) of the type as follows:
+p(24) 8+p(106) 3+4(108) 17–––––19––––––17––––––15
+p(24) p+v(106) 3+4(106)
giving the standard count 17-19-15 shared by more than 80% of the specimens (see Table 1 for variation). Scale row reduction formula (tail) as follows:
8 (1-4) - 6 (3-12) - 4 (14-29) - 2 (34-57) male (n = 60)
and
8 (1-4) - 6 (2-8) - 4 (7-22) - 2 (28-48) female (n = 61)
Ventrals smooth 163-181 in male (n = 60), and 166-187 in female (n = 65); anal entire; subcau-
Crotaphopeltis (Serpentes) of the Central African Republic dals paired, 44-58 in male (n = 63), 39-54 in female (n = 64).
Dentition
Maxillary teeth 12-16 + II + 1 (x = 14.6, s = 0.8, n = 51) in male, 13-16 + II + 1 (x = 14.2, s = 0.8, n = 52) in female, i.e. a small but significant (p <0.05) sexual dimorphism in number of solid maxillary teeth as in Dipsadoboa unicolor Günther, 1858 (Rasmussen 1996) . Palatine teeth 8-11 (x = 10.3, s = 0.7, n = 49), no sexual dimorphism.
Dimensions
Total length of type (female) 24 cm. Largest male 75 cm; largest female 70 cm; smallest specimen (with umbilical scars) 16 cm.
Hemipenes
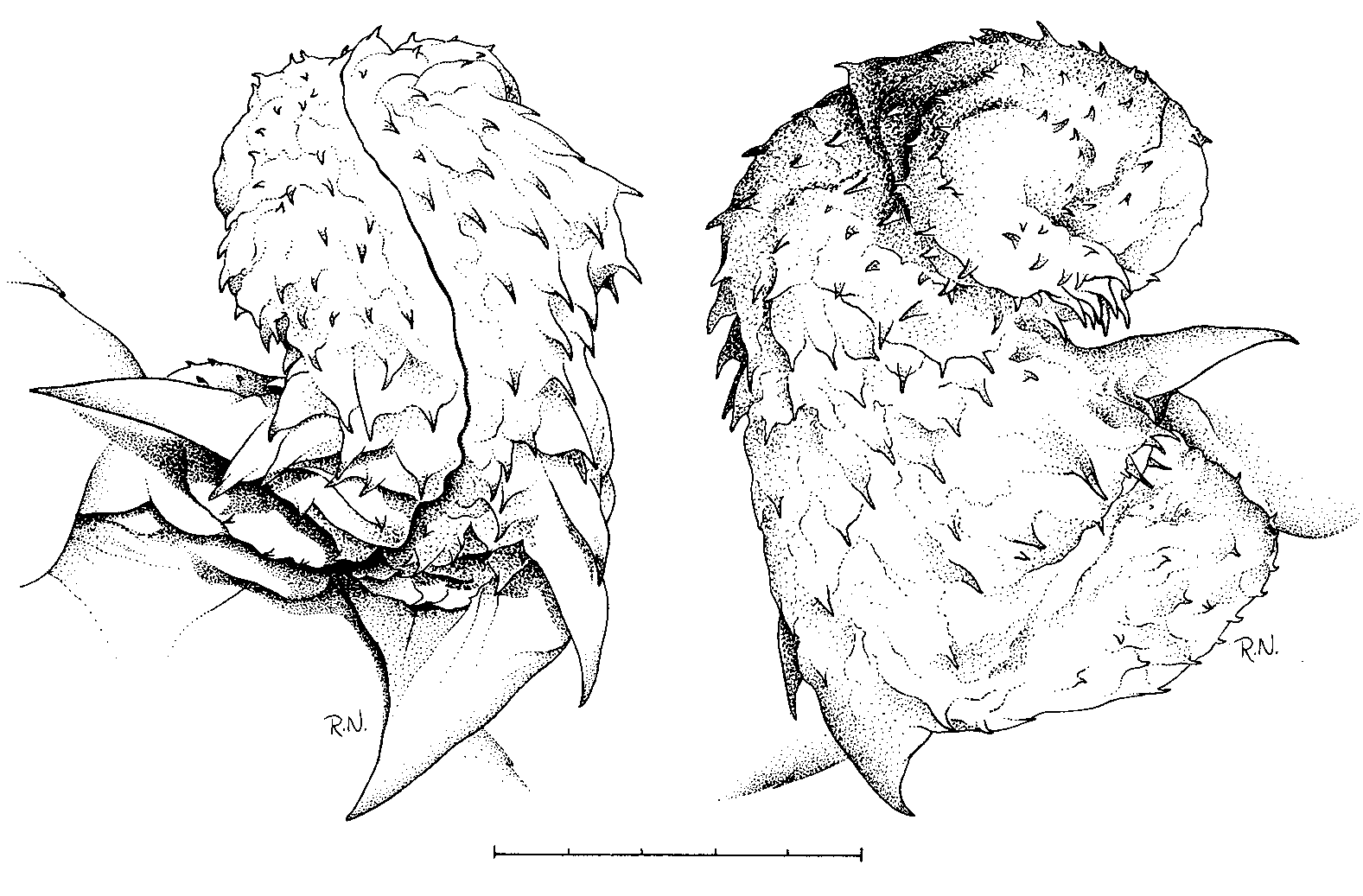
In situ the organs extend to subcaudal scute No. 8-12, (x = 10.1, s = 1.1, n = 38). In everted condition the hemipenes are characterised by two enlarged spines at the lower truncus, each with a row of accessory spines decreasing in size towards tip ( Fig. 3 View FIG ).
Internal morphology
Tongue sheath extending to ventral scute (VS) No. 15-21 in male (n = 38), 15-22 in female (n = 41), giving relative position 8.5-12.4% VS in male and 8.2-12.4% VS in female.
Tip of heart extending to ventral scute No. 34-42 in male (n = 44), 35-40 in female (n = 47), giving relative position 19.9-24.0% VS in male and 19.4-23.1% VS in female. Trachea extends to heart tip or up to 2 VS posterior to heart. Left lung 1-2 VS long.
Anterior end of liver situated at VS No. 45-51 in male (n = 41) and 42-51 in female (n = 45), giving relative position 25.3-29.6% VS in male and 23.1-28.7% VS in female. Absolute distance heart tip to anterior end of liver 7-13 VS in male and 6-13 VS in female.
Right kidney longer than left one; absolute length (right/left) 24-33/21-29 VS in male (n = 35) and 21-31/17-27 VS in female (n = 36), giving relative lengths 13.8-19.4/11.8-16.8% VS in male and 11.8-17.9/9.3-15.6% VS in female. Anterior end of kidneys situated at VS No. 130- 146/ 136-151 in male, 134-152/ 140-159 in female, giving relative position 76.5-82.0/79.8- 84.9% VS in male and 78.1-84.0/80.9-86.9% VS in female. Posterior ends of kidneys situated at VS No. 161-173/ 162-174 in male and 160- 180/ 161-181 in female, giving relative position 95.3-97.7/95.9-97.8% VS in male and 94.9- 98.9/95.4-97.7% VS in female.
Anal glands extending to subcaudal scute No. 3-5 (x = 3.6, s = 0.6, n = 25) in male and to 3-5, (x = 4.4, s = 0.6, n = 36) in female.
Coloration (in life)
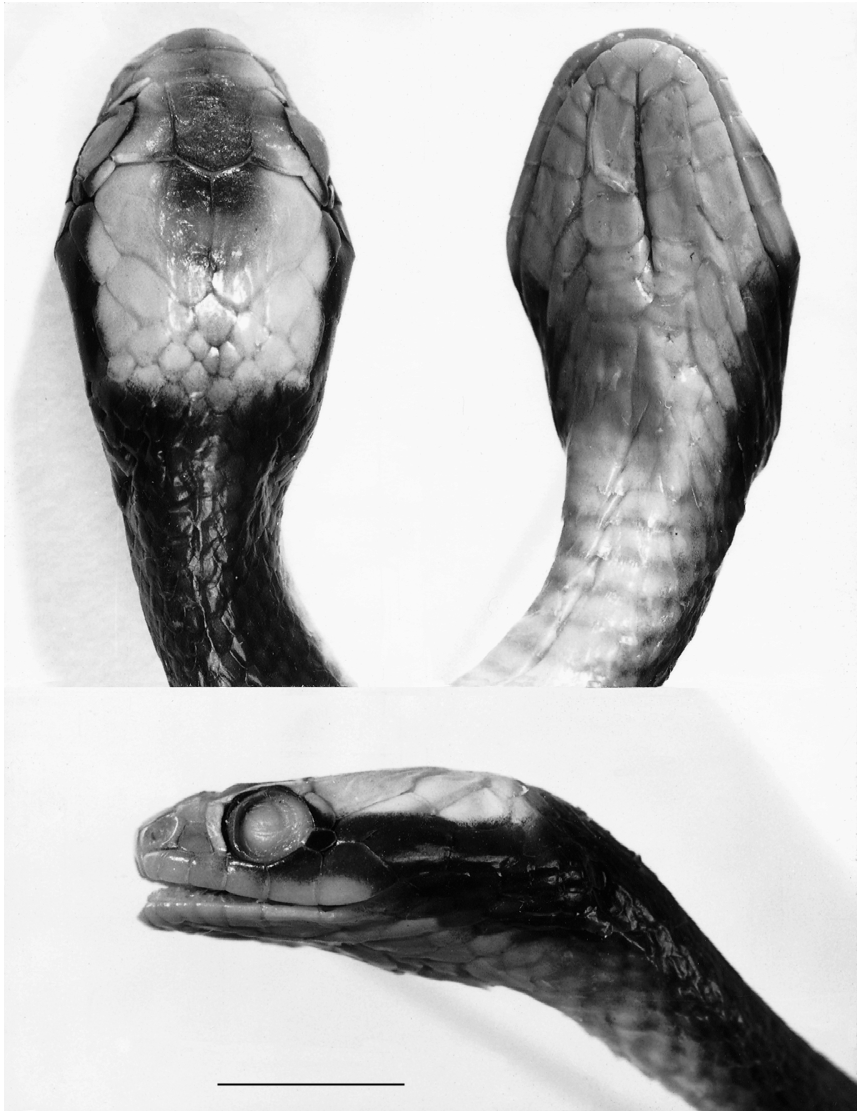
Dorsum dull chestnut, growing a little paler ventrally. Central part of each scale slightly paler than edge. Belly uniform whitish. Each side of head with dark chestnut band extending to posterior infralabials (Roman 1974).
Coloration (in preservative)
Dorsum various shades of brown, often chestnut, without white specks. Venter and underside of tail white, cream or yellowish, tail often with a pigmented median stripe. Occiput in juveniles either covered by two white parallel bars or by a horseshoe mark ( Fig. 4 View FIG ) which is overlaid by pigment in adults. Temples with a dark brown to black mark extending to lower jaw, covering most posterior two to five infralabials (Roman 1980: 52, upper figure). Upper labials pale in juveniles, gradually being overlaid by pigment with increasing size of specimens.
Biology
Crotaphopeltis hippocrepis has eyes of moderate to large size (greatest horizontal diameter of the eye 0.7-1.1 [x = 0.9, s = 0.1, n = 79] time the length of the snout and vertical pupils thus indicating a nocturnal way of life as in its congeners.
Crotaphopeltis hippocrepis occurs in moist (Guinean) savannas with more than 900 mm precipitation annually (Roman 1980); however, it also lives in drier (Sudanian) savanna in the Central African Republic and also in coastal thickets in Ghana (Hughes 1988). Apparently it mainly preys upon anurans, as only such food items are found in their intestines (n = 3), apart from a single juvenile specimen with some ants (which primarily may have been eaten by an anuran)!
By palpating bellies of gravid females (n = 3), four to ten eggs have been registered.
Distribution
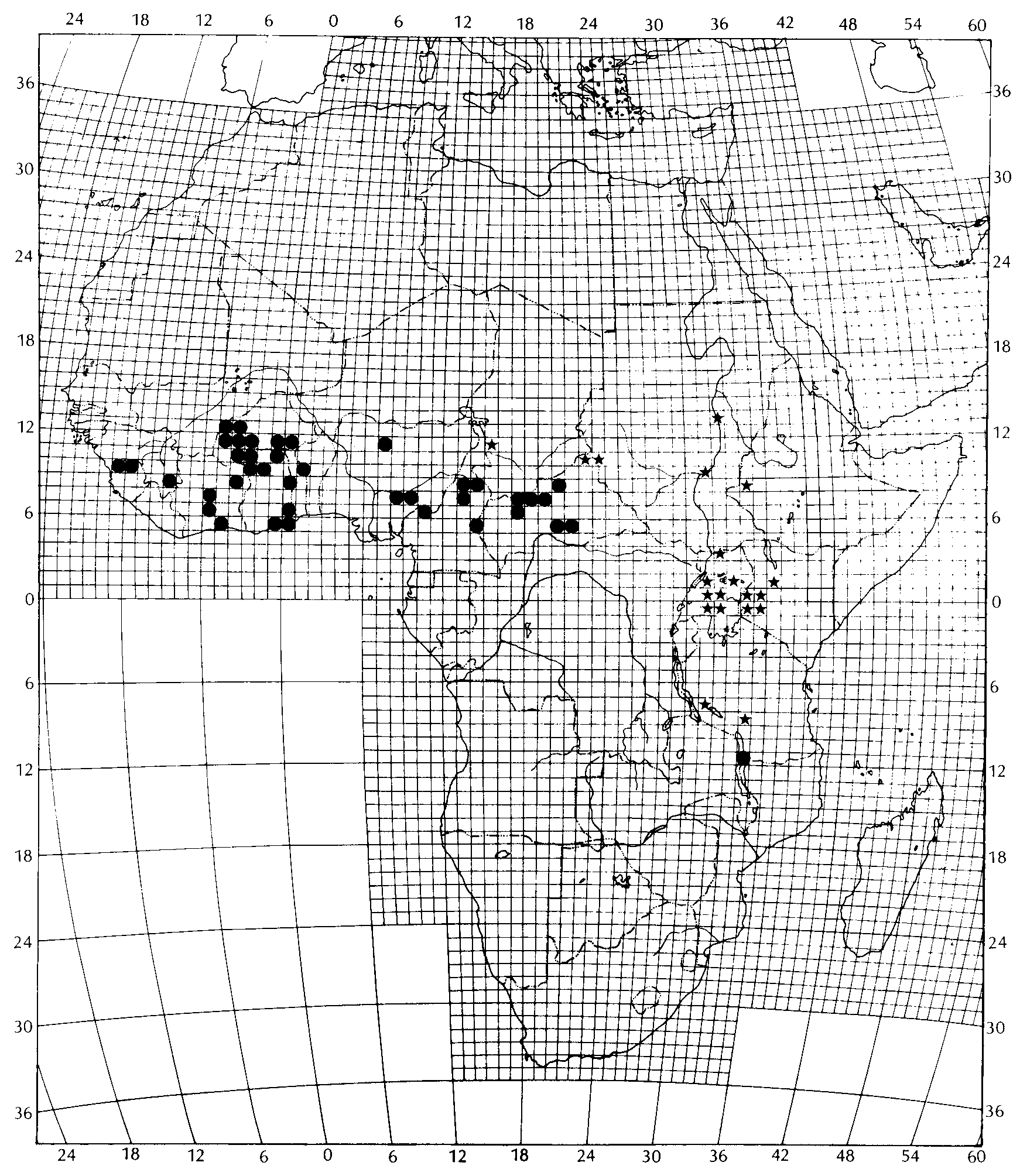
Crotaphopeltis hippocrepis is known from Guinea, Sierre Leone, Ivory Coast, Burkina-Faso, Ghana, Togo, Benin, Nigeria, Cameroon and the Central African Republic ( Fig. 2 View FIG ). Two records from East Africa, i.e. Victoria Nyansa (MCZ 6258) and Ugano in Matengo Hochland, Tanzania (NMW 25542:7), respectively, need confirmation (see Remarks).
Localities and material examined
Guinea. No specific locality, MHNG 722.95. — Beyla (Chabanaud 1921), MNHN 1921.462-463.
Sierre Leone. Saniya, USNM 248844. — Musaia, BMNH 1963.1056.
Ivory Coast. Adiopodume, MHNG 1565.9. — Bowake, FMNH 154659; MNHN 1987.1802-03. — Kotiessou, MNHN 1962.546. — Lamto (Roux- Estève 1969), MHNG 1464-65; MNHN 1977.383- 84, 1977.386-87, 1977.389, 1977.393, 1977.395-97, 1977.399-400, 1977.402-03, 1977.406, 1977.415- 16, 1977.418, 1992.4323, 1993.1771, 1993.3421, 1993.3442, 1993.7297, 1993.7299, 1994.7297, 1994.7299. — Nassian (Balletto et al. 1973), MSNG 44151. — Sakassou (near Toumodi), MHNG 1565.7.
Burkina Faso. Balave (Roman 1974). — Bomborokuy (Roman 1974). — Boromo (Roman 1974). — Dano (Roman 1974). — Farakoba, MHNG 1579.14. — Kampti (Roman 1974). — Nouna (Roman 1974). — Ouargaye (Roman 1974). — Ouarkoyo (Roman 1974). — Safane (Roman 1974). — Satiri (Roman 1974). — Sibi (Roman 1974). — Tionkuy (Roman 1974). — Zakuy (Roman 1974).
Ghana. No specific locality, ZFMK 63801, 63882- 83. — Accra SMNS 2864; ZMUC R63127, R63276. — Accra (5 mls E), CAS 125475. — Achimote, BMNH 1932.9.1.3; CAS 97515; FMNH 74847-49; MCZ 53709-10; ZFMK 63774-75. — Afienya (6 mls SW), CAS 125472. — Akosombo, ZFMK 63828. — Bombi Forest Reserve (near Damongo), ZFMK 63794. — Kpong ZFMK 63763. — Kwabenyan, BH C7H 165; ZFMK 63830. — Legon Hill, MCZ 53307; ZFMK 63751- 58, 63764-65, 63773, 63880; ZMUC R631238. — Mole National Park, ZFMK 63831. — Nalerigu, ZFMK 63829. — Nungua Farm, ZFMK 63881. — Shishe, USNM 223930. — Wa, BH C7H 188; BMNH 1982.439; ZFMK 63875-79, 63884-85; ZMUC R631239. — Zawse-Bawku, ZFMK 63793.
Togo. Bismarcksburg, ZMB 21346. — Pagala, USNM 223867.
Benin. Soubroukou, USNM 223778-79.
Nigeria. Northern part, BMNH 1975.623. — Makurdi, BMNH 1937.12.4.52-56. — Samaru, BMNH 1975.621. — Wukari, BMNH 1938.3.1.122, 1938.3.1.124-33.
Cameroon. Bafout (Joger 1982; Böhme & Schneider 1987). — Boli (near Garoua), MCZ 44107. — Garoua-Boulai, MHNG 1461.63. — Banguve, 15 km from Ngaouyanga (Monard 1951). — Ngaouyanga (Monard 1951). — Tchollire (Joger 1982; Böhme & Schneider 1987).
Central African Republic. Bangbali, MNHN 1997.3522. — Bossangoa, MNHN 1994.7392, 1994.7401. — Gribingui River (Mocquard 1896; Joger 1990), MNHN 1921.19. — Kaga Bandoro (Fort Crampel), MNHN 1998.251. — Kouki, MNHN 1994.3273-76, 1994.3278-83, 1994.7410-11, 1994.7419, 1994.7423-24, 1994.7427, 1994.7437- 38, 1994.7442. — Seko, MNHN 1996.6531-32. Sogesca (South of Bambari), MNHN 1992.4618.
Victoria Nyansa. MCZ 6258.
Tanzania. Ugano (Matengo Hochland), NMW 25542: 7.
Geographical variation
Populations from only four countries are reasonably well-represented. With respect to number of ventrals, the males exhibit a smooth east-west cline with decreasing numbers of ventrals ( Table 2). The females, however, have an increasing number of ventrals until Ghana and then low counts for the Ivory Coast ( Table 2).
With respect to subcaudals, the highest numbers in both sexes are found in Nigeria from where they declines eastwards and westwards ( Table 3). With respect to number of solid maxillary teeth, the highest numbers in males are found in Nigeria as well, whereas the females exhibit a smooth east-west cline with decreasing numbers of maxillary teeth ( Table 4).
As to the relative position of the heart, both sexes have highest values in Nigeria, i.e. the heart is more posteriorly positioned ( Table 5).
In opposition to two other species within the genus ( C. degeni and C. tornieri ), the distribution of C. hippocrepis seems to be reasonably continuous and the variation only of a small scale, making C. hippocrepis a homogeneous entity.
REMARKS
The two East African records of this species seem dubious. José P. Rosado ( in litt.) kindly informs the senior author that the specimen from Victoria Nyansa was “purchased from the Linnean Natural History Institute in 1889 [...] and that many of the catalogue entries during that period have been rejected (discarded) for bad locality information”. (The Linnean Natural History Institute is probably the same as the one Gans [1966: 232] mentions, i.e. a commercial trading concern which purchased collections, arranged for them to be identified by specialists, and then sold their components to various museums).
Loveridge (1942) examined the series of which the specimen from Ugano, Matengo Hochland, is supposed to be a part. According to Loveridge, seven specimens were examined, one of which was exchanged with Harvard, leaving six specimens in Vienna. Nonetheless the catalogue in Vienna today reads that the series comprises eight specimens!
Albeit Loveridge was not especially keen in his use of numerals, it is fairly evident that the present specimen (, SC 48/48 + 1, total length 66 cm) was not included in his counts. It is thus possible that the present specimen has not been a part of the original series, but rather of another one.
On the basis of number of ventrals, subcaudals, and maxillary teeth plus the position of the heart, it is possible to assign these two specimens to any of the various populations.
| ZMUC |
Zoological Museum, University of Copenhagen |
| MHNG |
Museum d'Histoire Naturelle |
| MNHN |
Museum National d'Histoire Naturelle |
| FMNH |
Field Museum of Natural History |
| MSNG |
Museo Civico di Storia Naturale di Genova 'Giacomo Doria' |
| ZFMK |
Zoologisches Forschungsmuseum Alexander Koenig |
| SMNS |
Staatliches Museum fuer Naturkund Stuttgart |
| CAS |
California Academy of Sciences |
| MCZ |
Museum of Comparative Zoology |
| ZMB |
Museum für Naturkunde Berlin (Zoological Collections) |
| NMW |
Naturhistorisches Museum, Wien |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Crotaphopeltis hippocrepis (Reinhardt, 1843)
| Rasmussen, Jens Bødtker, Chirio, Laurent & Ineich, Ivan 2000 |
Crotaphopeltis acarina
| Roman 1974: 6 |
Dipsas hippocrepis
| Reinhardt 1843: 26 |