
Orobdella tsushimensis Nakano, 2011
|
publication ID |
https://doi.org/ 10.12782/sd.17.2.235 |
|
persistent identifier |
https://treatment.plazi.org/id/B71D685D-FFE0-FFA8-FE9F-F9B5FA930C52 |
|
treatment provided by |
Felipe |
|
scientific name |
Orobdella tsushimensis Nakano, 2011 |
| status |
|
Orobdella tsushimensis Nakano, 2011
urn:lsid:zoobank.org:act:4294992A-9445-4C67-BEBE-213D8570E75C ( Figs 2–4 View Fig View Fig View Fig )
Orobdella tsushimensis Nakano, 2011a: 41–46 , figs 2–4.
Diagnosis (amended from Nakano 2011a). In life, dorsal surface brownish yellow or reddish, ventral surface whitish yellow. Somites III and IV both uniannulate, somites VIII–XXV quadrannulate, somite XXVI triannulate. Clitellum in XI b5 to XIII a2. Pharynx reaching to XIV. Gastropore conspicuous at XIII a1/a2. Gastroporal duct bottle-shaped, slightly winding at junction with gastropore. Male gonopore at XI b6 (slightly anterior to middle of annulus), female gonopore inconspicuous in XIII a1/a2, behind gastropore, gonopores separated by 1/2+5 annuli. Paired epididymides in XVII–XIX (i.e. occupying approximately two somites). Atrial cornua developed, coniform or ovate.
Material examined. Three specimens collected from along mountain trails on Mt. Doiksilsan (alt. 639 m, 34°04′35″N, 125°06′27″E), Gageodo Island , Heuksan-myeon , Sinan-gun, Jeollanam-do, the Republic of Korea, by Hong-Yul Seo: KUZ Z213 View Materials , dissected, on 26 July 2011; KUZ Z214 View Materials on 23 August 2011; and KUZ Z215 View Materials , dissected, on 25 July 2011 GoogleMaps .
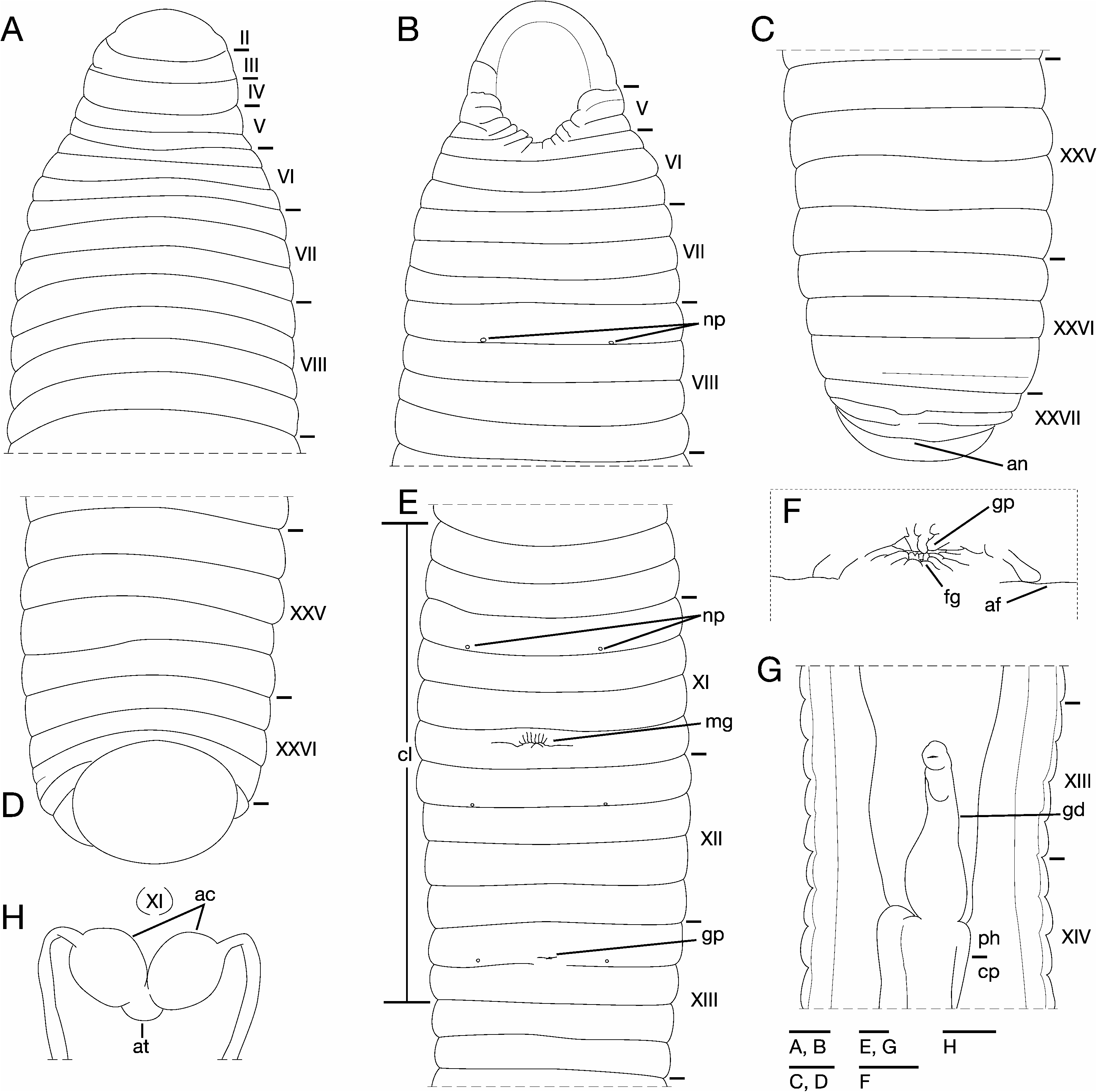
Description of Gageodo Island specimens. Body firm and muscular, elongate, gaining regularly in width in caudal direction in cephalic part, dorso-ventrally depressed, sides nearly parallel from midlength to point just anterior of caudal sucker, maximum BL 112.5 mm, BW 9.38 mm (KUZ Z213) ( Fig. 2A, B View Fig ), minimum BL 40.2 mm, BW 2.84 mm (KUZ Z214). Caudal sucker ventral, oval, its greater diameter smaller than BW ( Fig. 4B View Fig ). In life, dorsal surfaces black or brown, dorso-lateral surface reddish ( Fig. 3 View Fig ), ventral surface whitish yellow. Color faded in preservative, without any dark lines ( Fig. 2A View Fig ).
Somite I completely merged with prostomium ( Fig. 4A View Fig ). Somites II–IV uniannulate; slight dorsal furrow present in III (KUZ Z215); similar furrow also present in IV (KUZ Z213, 215). Somite V biannulate; (a1+a2)>a3 ( Fig. 4A, B View Fig ); (a1+a2) with slight ventral furrow reaching to lateral sides of body (KUZ Z215); a3 forming posterior margin of oral sucker ( Fig. 4B View Fig ). Somites VI–VII triannulate ( Fig. 3A, B View Fig ). Somites VIII–XXV quadrannulate; a1=a2=b5=b6 ( Fig. 4A–E View Fig ); b5 of X and a2 of XIII respectively being first and last annuli of clitellum ( Fig. 4E View Fig ). Somite XXVI triannulate; a1=a2<a3; a3 with slight dorsal furrow (KUZ Z213) ( Fig. 4C View Fig ). Somite XXVII with a few furrows; anus behind it with no post-anal annulus ( Fig. 4C View Fig ).
Eyes in three pairs, first pair dorsally on posterior margin of II, second and third pairs dorsolaterally on posterior margin of V (a1+a2) or undetectable (KUZ Z213). Nephridiopores in 17 pairs in VIII–XXIV, situated ventrally at posterior margin of a1 of each somite ( Fig. 4B, E View Fig ). Papillae numerous, minute, hardly visible, one row on every annulus.
Pharynx agnathous, euthylaematous, reaching to XIV a2– b5. Crop tubular, reaching to XIX b6. Gastropore conspicuous, ventral, in furrow of XIII a1/a2 ( Fig. 4E, F View Fig ). Gastroporal duct bottle-shaped, slightly winding at junction with gastropore, joining with crop in XIV a2/b5 ( Fig. 4G View Fig ). Intestine tubular and acecate, reaching to XXIV a2–XXIV/XXV. Rectum tubular, thin-walled.
Male gonopore slightly anterior to middle of XI b6 ( Fig. 4E View Fig ). Female gonopore in furrow of XIII a2/b5, inconspicuous, located posterior to gastropore ( Fig. 4F View Fig ). Gonopores separated by 1/2+5 annuli ( Fig. 4E View Fig ). Testisacs multiple, in XIX b5–b6 to XXV b6. Paired epididymides in XVII a1/a2– a2/b5 to XIX a2/b5–b5/b6. Ejaculatory bulbs absent. Ejaculatory ducts in XI b5 to XVII a1/a2–a2/b5, loosely coiled, each widening from respective junction with epididymis, narrowing at junction with atrial cornua, then turning gradually inward toward atrial cornua without pre-atrial loop ( Fig. 4H View Fig ). Pair of ovate, muscular atrial cornua in XI b5 and b6 ( Fig. 4E View Fig ). Atrium short, muscular, globular, in XI b5 and b6. Penis sheath and penis absent. One pair of ovisacs, thinwalled, globular, in XIII a2 and b5. Oviducts thin-walled, right oviduct crossing ventrally beneath nerve cord, both oviducts converging into common oviduct in XIII a2. Common oviduct thin-walled, short, directly ascending to female gonopore.
Distribution. Known from mountainous regions of Tsushima Island, Japan ( Nakano 2011a) and Gageodo Island, Korea ( Fig. 1 View Fig ).
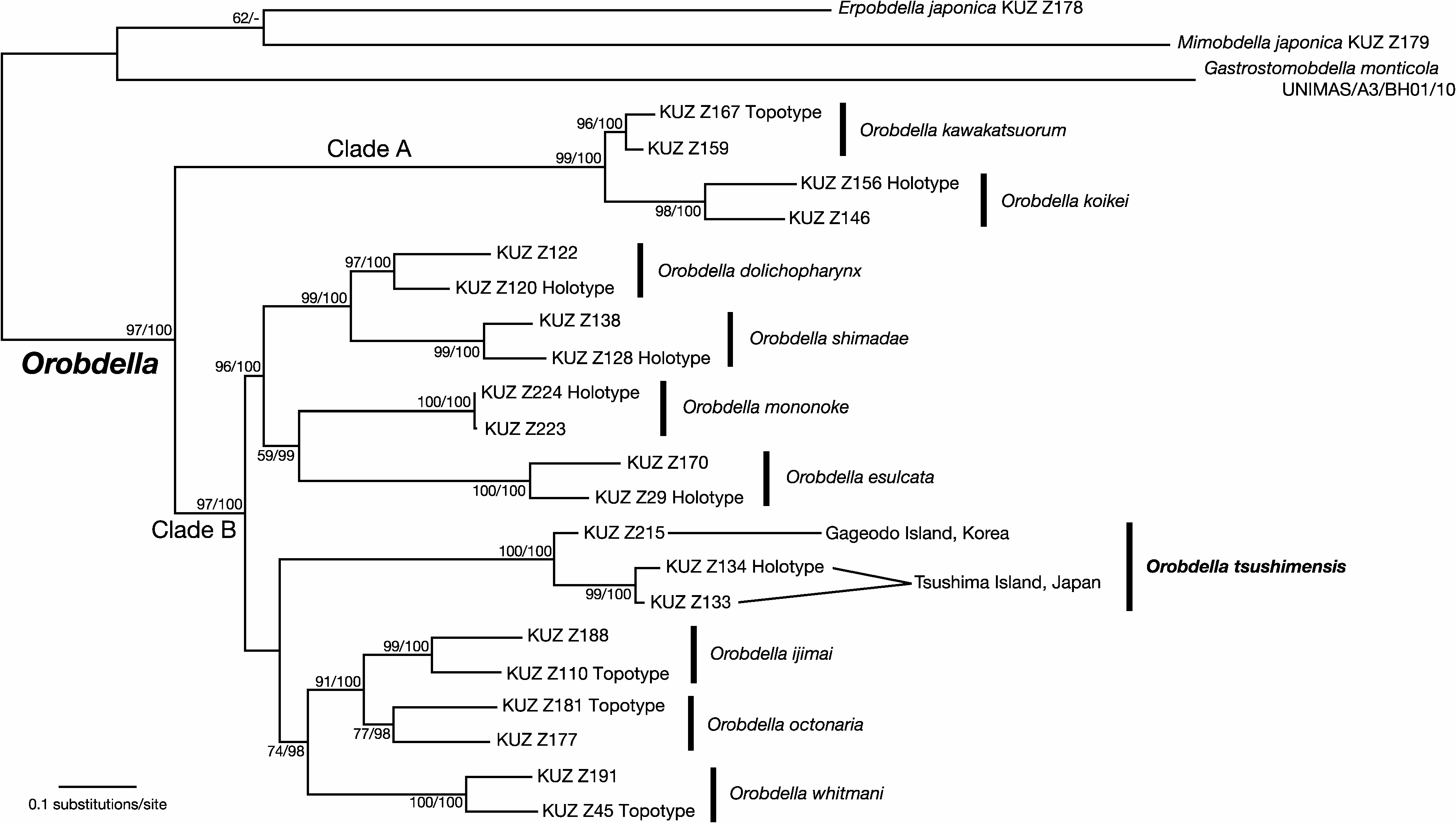
Phylogenetic position. The ML tree with ln L = −13384.71 ( Fig. 5 View Fig ) was nearly identical to the obtained BI tree (not shown but see below). Monophyly of the genus Orobdella was confirmed (BS=97%, BPP=100%). Orobdella was divided into two subclades: Clade A consisting of two species from Hokkaido, Japan, O. koikei Nakano, 2012 and O. kawakatsuorum Richardson, 1975 ; and Clade B composed of the other seven species distributed in areas south of Hokkaido. In the BI tree, however, Clade B was divided into a polytomy of three clades: O. tsushimensis ; (( O. dolichopharynx Nakano, 2011 + O. shimadae Nakano, 2011 )+ ( O. esulcata Nakano, 2010 + O. mononoke Nakano, 2012 )); and (( O. octonaria Oka, 1895 + O. ijimai Oka, 1895 )+ O. whitmani ). Monophyly of O. tsushimensis from Gageodo Island and Tsushima Island was well supported (BS=100%, BPP=100%). In the ML tree, O. tsushimensis and the (( O. octonaria + O. ijimai )+ O. whitmani ) clade formed a monophyletic clade, but with low support (BS=25%). The COI K2P distance between the specimen in Gageodo Island and those in Tsushima Island was between 5.2% and 5.5%.
Remarks. The specimens of Orobdella from Gageodo Island were identified as O. tsushimensis based on their possession of the following characteristics: IV uniannulate; male gonopore in the middle of XI b6, female gonopore in the furrow of XIII a1/a2, 1/2+5 annuli between the gonopores; a bottle-shaped gastroporal duct; and epididymides located in approximately eight successive annuli in XVII–XIX. Although the specimens from Tsushima Island possess conical atrial cornua ( Nakano 2011a), the specimens from Gageodo Island have ovate atrial cornua ( Fig. 4H View Fig ). In addition, the specimens from Gageodo Island have a reddish dorsal surface in contrast to a brownish yellow dorsal surface of the specimens from Tsushima Island. The diagnosis of O. tsushimensis in Nakano (2011a) was amended above to take these differences into account.
In our molecular phylogenetic analyses, the specimens of O. tsushimensis from Gageodo Island and Tsushima Island together form a well-supported monophyletic clade (BS= 100%, BPP=100%) ( Fig. 5 View Fig ) and the COI sequence divergence between them is only 5.2–5.5%. For comparison, the mean intraspecific variation of COI sequences of O. koikei is 7.1% ( Nakano 2012a). Our molecular phylogenetic analyses and distance data thus provide addition confirmation that the Orobdella leeches from Gageodo Island are O. tsushimensis .
This is the first record from Korea of both the genus Orobdella and the monotypic family Orobdellidae , which was delineated by a recent molecular phylogenetic study ( Nakano et al. 2012). Orobdella tsushimensis has been reported only from Tsushima Island and Gageodo Island, which are both located offshore of the Korean Peninsula ( Fig. 1 View Fig ). There are no records of Orobdella from the Kore- an Peninsula itself. Gilyarov et al. (1969) reported a find of Orobdella species from the continent (Primorsky Krai, Russia), although they appear to have misidentified the species. Thus, it is highly likely that Orobdella leeches are also present in the Korean Peninsula. Further faunal surveys are needed to confirm this, and to reveal more fully the evolutionary and biogeographical history of this genus.



| KUZ |
Zoological Collection of the Kyoto University |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Orobdella tsushimensis Nakano, 2011
| Nakano, Takafumi & Seo, Hong-Yul 2012 |
Orobdella tsushimensis
| Nakano, T. 2011: 46 |