
Swima fulgida, Osborn & Haddock & Rouse, 2011
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2011.00727.x |
|
DOI |
https://doi.org/10.5281/zenodo.10545856 |
|
persistent identifier |
https://treatment.plazi.org/id/AA7B8713-7154-5157-0CFA-202DFDE8FA36 |
|
treatment provided by |
Valdenar |
|
scientific name |
Swima fulgida |
| status |
sp. nov. |
SWIMA FULGIDA View in CoL SP. NOV.
Green bomber sp. 1 ( Osborn et al., 2009), shining bomber ( Osborn & Rouse, 2010).
Common name: Shining bomber
Type material: Holotype, collected off the central coast of California 7.x.2004 at 3267 m over a bottom depth of 3546 m by K. J. O. and S. H. D. H., deposited at the Benthic Invertebrate Collection of Scripps Institution of Oceanography ( SIO BIC A1285 ; 35°46.38′N, 122°50.24′W). Two female paratypes collected by K. J. O. ( SIO BIC A1286 , 22.vi.2006, at 3478 m, 36°20.08′N, 122°55.00′W, undetermined live length; A1675, 26.ii.2009, at 3625 m, 35°7.61′N, 122°55.60′W,> 30 mm). GoogleMaps
Diagnosis: Member of Swima with a darkly pigment anterior gut and buccal organ. Possessing single, subulate, medial branchia, thick, transparent gelatinous sheath penetrated throughout by narrow clavate papillae, simple noto- and neurochaetae, and three achaetous anterior segments supporting ellipsoid, bioluminescence-producing, derived branchiae that are less than 1.2 mm in length.
·
Etymology: Named for the shining or gleaming (fulgidus) bioluminescence produced by the four pairs of ellipsoid, segmental branchiae. Feminine.
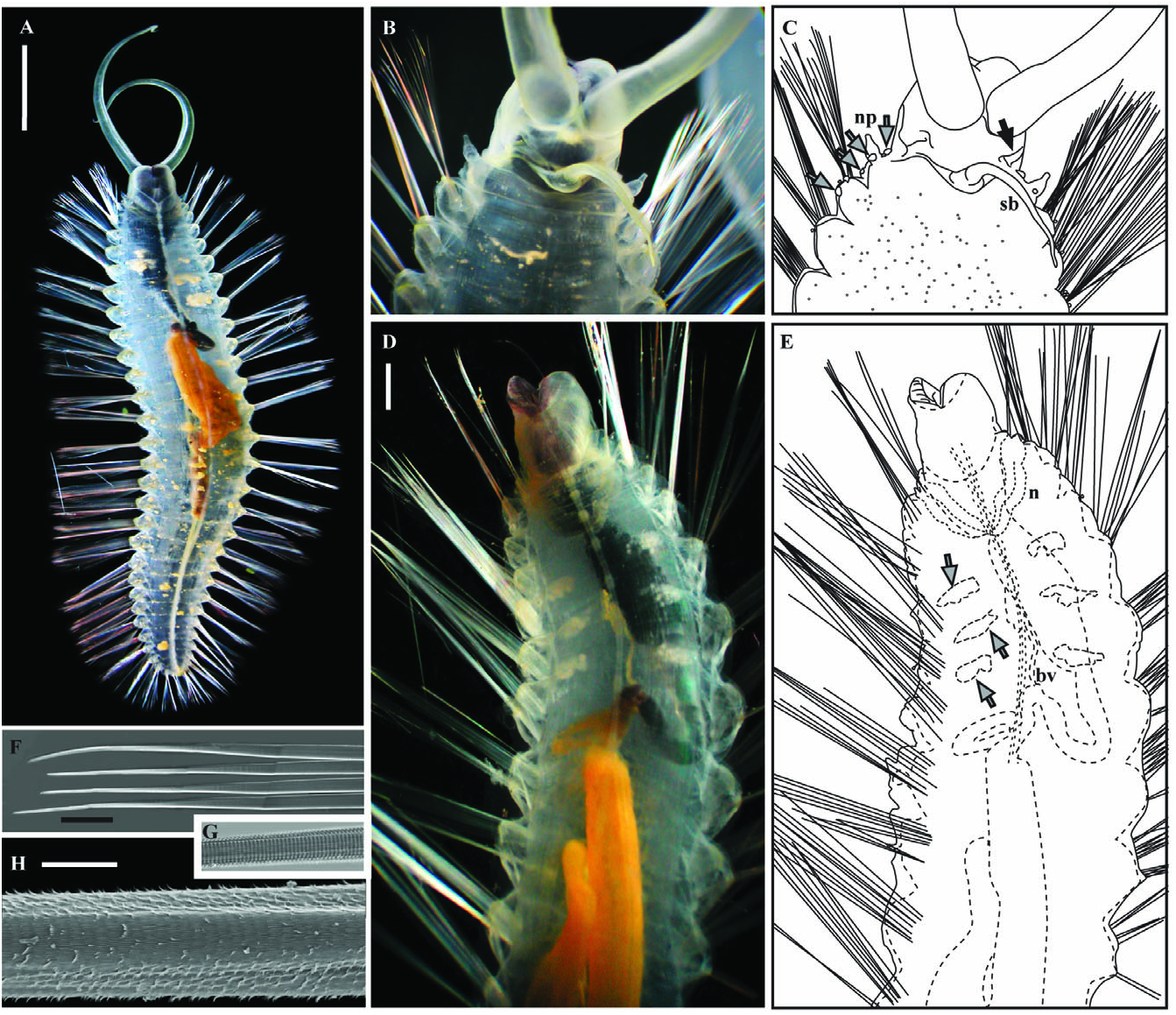
Holotype description: Body transparent, 27 chaetigers, 29 mm live body length, distinct parapodial lobes, numerous long chaetae, posterior half smoothly tapered, broadest segments after chaetiger 3 ( Fig. 4A). Thick gelatinous sheath through which narrow, clavate, yellow (in life) papillae, extend ( Fig. 4B). Papillae especially numerous on parapodia and dorsum of anterior segments. Pygidium unadorned.
Head consists of prostomium, peristomium, and three achaetous segments possessing what is interpreted here as three forms of branchiae ( Fig. 4B, E, but see Variation). Prostomium limited to tissue posterior to palps supporting a pair of low, ciliated, oblique ridge-like nuchal organs (shown in paratype 1 Fig. 5C View Figure 5 ). No eyes. Grooved frontal peristomial palps transparent to yellow in life, tapered, coiling tips, long, reaching at least sixth chaetiger ( Fig. 4A). Peristomium surrounds prostomium completely. Buccal organ anteroventrally located, unarmed, bilobed, forming eversible lateral lips. Lateral lips with dark pigment, purple-brown in life, inner lobe transparent to purple-brown in life ( Fig. 4B). Slight ridge found posterior to nuchal organs (anterior to digitiform and medial branchiae) possibly indicating a segment margin ( Fig. 4B). Three forms branchiae: (1) single, long (reaches to fourth chaetiger), tapered/subulate, medial projection interpreted here as branchia, transparent to white ( Fig. 4B) peristomial or on segment 1; (2) more than 40, fine, digitiform respiratory branchiae present across lateral and dorsal surface segment 1 or 2 in tightly packed, single row, yellow in life ( Fig. 4B); and (3) three pairs elliptical lobe-like branchiae on achaetous anterior segments (posterior to all other forms branchiae) and one pair on chaetiger 1 (referred to colloquially as ‘bombs’ because they burst into light when dropped by animal; Fig. 4E, F). Segmentally occurring, elliptical, lobe-like branchiae greenish-yellow in life, autofluorescent, produce green bioluminescence when detached, often autotomized, 0.7 to 1.1 mm in length ( Fig. 4E). All elliptical branchiae detached from holotype during collection, examination, and/or storage. Scars from detached elliptical branchiae distinguishable in live and preserved specimen as slightly raised rings thickened tissue, four pairs: one slightly ventral from lateral midline just posterior to digitiform branchiae, one posterior on medial half of nephridiopore papillae, one posterior to nephridiopore papillus at lateral midline, one posterior to chaetae on first notopodium (shown in paratype 1 Fig. 5B View Figure 5 ).
Chaetigers similar along body. Noto- and neuropodial lobes form continuous, nearly smooth parapodium ( Fig. 4H) with more fine papillae relative to rest of body surface. One to four, white to brown, clavate papillae with rounded bulbous tips and narrow bases (‘lollipop’-shaped) found between noto- and neuropodial lobes, projecting well beyond gelatinous sheath, tips solid ( Fig. 4C). Noto- and neurochaetae indistinguishable except by position, simple, with no articulations ( Fig. 4D, G, shown in paratype 1 Fig. 5F View Figure 5 ). High magnification reveals fine whorls of spines making distal tips appear segmented ( Fig. 4D, G, shown in paratype 1 Fig. 5G View Figure 5 ), bases appear striated. Distal edge of spinous whorls project as frayed edges on worn and longest chaetae ( Fig. 4D, shown in paratype 1 Fig. 5H View Figure 5 ). Chaetigers 4–6 each with a pair of gonopores, low, hollow papillae at ventral base of neuropodia ( Fig. 4E, F).
Internal anatomy visible through transparent body wall and gelatinous sheath ( Fig. 4A, shown in paratypes Fig. 5A, D View Figure 5 ). Ventral nerve cord with two paired, fused ganglia in each segment, diverge just posterior to peristomium to surround buccal organ, fused again just posterior to palp attachment points ( Fig. 4E, shown in paratypes Fig. 5A, D View Figure 5 ). Single pair of large, anterior, semitransparent nephridia reaching second chaetiger, largely lying in ventral part of coelum, overlapping each other in ventral portions of first and second chaetigers, folding back anterodorsally, then narrowing, with each nephridium leading to a lateral nephridiopore (shown in paratype 2 Fig. 5D–E View Figure 5 ). Gut running from buccal organ straight for one-third body length (approximately to chaetiger 10) after which a wide, single loop is formed. Gut in this loop portion broadens (to at least one quarter body width) and continues back to approximately chaetiger 20 before narrowing and turning anterior. Gut continues anterior to near first loop then folds rearward and continues directly to pygidium. Foregut (region anterior to the first loop) darkly pigmented, appearing black through body wall, expandable to near body width ( Fig. 4A, E, shown in Paratypes Fig. 5A, D, E View Figure 5 ). Heart body and dorsal blood vessel first distinguishable just posterior to digitiform branchiae, extends through anterior one-third of body until apparently merging with anterior-most dorsum of broadened portion of gut (shown in paratype 2 Fig. 5D, E View Figure 5 ). Gonads form at posterior margin of chaetigers 4–6 (shown in paratype 2 Fig. 5D, E View Figure 5 ).
Variation: The two paratypes had 27 and more than 28 chaetigers (specimen incomplete), respectively. Each was more than 30 mm total body length when alive. Swima are particularly prone to distortion when injured and to preservation artefacts. If specimens were not completely relaxed prior to fixation, they contracted dramatically during initial fixation. Additionally, when specimens were injured they often contracted around the injury site. The degree of buccal organ lateral lobe eversion varied amongst preserved specimens.
Palps were easily dislodged from specimens during handling leaving obvious scars. Palps varied in length relative to body length. The length of the single, medial branchia also varied in length relative to body length. Digitiform branchiae were not easily lost from the holotype and left obvious scars when they were. Digitiform branchiae were absent from both paratypes genetically confirmed to belong to the species. Elliptical branchiae were easily autotomized, even during the gentlest collection, and were often found on the floor of the sampling device upon recovery of the ROV or were detached during laboratory examination. As no specimens were recovered with all elliptical branchiae attached, it was not possible to determine if more than one pair can be attached to a single segment at one time, but this seems unlikely based on the scars. Various sized elliptical branchiae, ranging from 0.6–1.2 mm in longest dimension, were found on a single individual. Smaller ‘bombs’ were nearly spherical whereas larger branchiae were ellipsoidal.
Paratype 1 with obvious gonopores only on chaetiger 6 and developing gonads in the right posterior margin of chaetiger 4 and left posterior margin of chaetigers 4–6 ( Fig. 5A View Figure 5 ). Paratype 2 with developing gonads in chaetigers 4–6 ( Fig. 5D View Figure 5 ), obvious gonopores only in chaetiger 5. Oocytes in female up to 0.5 mm in diameter.
Remarks: Swima fulgida sp. nov. is most similar to S. bombiviridis . The presence of dark pigment on the foregut and lateral lips of the buccal organ, as well as the differences found in COI and cyt b sequences clearly separate the two species. Additionally, specimens of S. fulgida sp. nov. tend to be larger than those of S. bombiviridis and are broadest after the third chaetiger, unlike S. bombiviridis , which tapers from the head and first chaetiger. Swima fulgida sp. nov. has fewer prominent interramal lollipop-shaped papillae than S. bombiviridis . See Remarks below for comparison to S. tawitawiensis sp. nov.
It cannot be determined at this time if the structures referred to here as subulate and digitiform branchiae are peristomial or segmental. Unlike other branchiae known in acrocirrids and flabelligerids ( Spies, 1975), these structures are not easily detached. The digitiform branchiae were not lost from the two paratypes by handling or damage to the specimen; they were absent. Absence of digitiform branchiae was also observed in S. bombiviridis where specimens were observed with numerous long, few short, or absent digitiform branchiae. Pulling on both digitiform and subulate branchiae of both live and dead specimens resulted in breakage of the structure at various positions along their lengths and left ragged, torn edges. This contrasts with the removal of palps and elliptical lobe-like branchiae, which always come away from the body at the base and leave a regular, sealed scar.
The spherical tips of the lollipop-shaped papillae found in S. bombiviridis and S. fulgida sp. nov. should not be confused with the balloon-like tips of the notopodial papillae of Flabelliseta incrusta . The tips of the lollipop-shaped papillae are solid, with a granular outer appearance whereas the balloon-shaped papillae are described as hollow with a smooth outer surface.
Ecology: Swima fulgida sp. nov. was found off of the central California coast at 3267–3625 m depth, from 30–340 m above the seafloor. Similar animals were observed via ROV but not collected (identification unconfirmed) off the Oregon coast (45°24.02′N, 126°43.00′W), as well as the Gulf of California (24°18.99′N, 109°11.95′W) by K. J. O. Animals were not observed on the seafloor although they were sometimes observed within sight of it via the ROV. The proximity to the seafloor at great depth, ability to swim, and delicate body are probably the reasons why this species was only recently discovered GoogleMaps .
Animals were observed hanging horizontally in the water column with the dorsal surface uppermost and the palps hanging forward and downward over the buccal organ, which typically projects anteroventrally. Swima fulgida sp. nov., like others in the clade, swims by lateral undulation of the body coupled with expansion on the power stroke and contraction on the recovery stroke of the chaetal fans. Observations of parapodia removed from the animal suggest that this expansion and contraction of chaetal fans can be attributed to passive mechanical mechanisms, not requiring musculature. Swimming was observed to be both forward and rearward, which was difficult to distinguish unless the ROV was completely still and camera zoomed in enough to identify the anterior end. Animals were seldom observed in situ in close enough detail to determine the direction of their initial ·
swimming when disturbed, but in the instances where it was possible, it was consistently rearward. This is consistent with the direction of escape swimming observed in other Swima species. A rearward escape response is also consistent with use of anteriorly located, autotomizable, bioluminescent decoys by removing the bulk of the body away from the released body part or decoy.
Specimens were recovered in various conditions and possessed from two to eight elliptical branchiae of various sizes. The first pair of elliptical branchiae differed slightly from all other pairs in that the first pair was always smaller than, or the same size as, the smallest bomb found on an individual. The second to fourth bombs varied in size; this was assumed to be because of regeneration of previously autotomized bombs. Manual stimulation of animals at any point along the body or head resulted in release of a bomb or two, which immediately produced green bioluminescence. Further stimulation would result in release of additional bombs if available. Bioluminescence was seen as a steady glow from bombs that had been autotomized. Bombs that were separated from the animal could be triggered to produce light again by gently squeezing them with forceps. The glow of an individual bomb lasted several seconds.
| SIO |
Scripps Institution of Oceanography |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |