
Dentex carpenteri, Iwatsuki, Yukio, Newman, Stephen J. & Russell, Barry C., 2015
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3957.1.9 |
|
publication LSID |
lsid:zoobank.org:pub:5C8F0550-A56A-44B2-88E6-CCB4758B4BF8 |
|
DOI |
https://doi.org/10.5281/zenodo.5664237 |
|
persistent identifier |
https://treatment.plazi.org/id/B4CF5E5A-ECB7-4FBE-940C-2830B344FCC6 |
|
taxon LSID |
lsid:zoobank.org:act:B4CF5E5A-ECB7-4FBE-940C-2830B344FCC6 |
|
treatment provided by |
Plazi |
|
scientific name |
Dentex carpenteri |
| status |
sp. nov. |
Dentex carpenteri new species
New common name: Yellow Snout Seabream Figure 1 View FIGURE 1 A–B
Dentex tumifrons View in CoL (not of Temminck & Schlegel): Allen 1997: 132 (Western Australia); Carpenter 2001: 3000 (Western Pacific, in part).
Allotaius spariformis: Hutchins 2001: 35 View in CoL ( Australia,?in part).
Holotype. WAM P.33486-001 (X), 219 mm SL, female with gonad, off Ningaloo Reef near Exmouth, Western Australia (22°10.02’S, 113°46.21’E), 26 March 2011, trapped, depth 200– 210 m.
Paratypes. WAM P.33490-001 (8 specimens), 145–189 mm SL, off Ningaloo Reef near Exmouth, Western Australia (22°10.02’S, 113°46.21’E), 26 March 2011, trapped, depth 200–210 m (only 171 mm SL, X; sex not determined but presumably male, Fig. 1 View FIGURE 1 A).
Diagnosis. A species of the Dentex hypselosomus complex ( Iwatsuki et al. 2007) with the following combination of characters: posterior hind margin of upper jaw not reaching to or reaching slightly beyond vertically through anterior margin of eye; often yellow wide region in greater part from snout to infraorbital plus often yellow upper jaw; deeper suborbital depth (9.7–10.9% SL; Fig. 3 View FIGURE 3 A); orbit diameter larger (11.7–13.5% SL Fig. 3 View FIGURE 3 B); considerably deeper body, especially in specimens of about 100−200 mm SL (50.0–55.0% SL; Fig. 1 View FIGURE 1 A– B).
Description. Meristic values counts and measurements as percentages of standard length (SL) for the holotype and eight paratypes of Dentex carpenteri n. sp. are in Table 1 View TABLE 1 . Characters presented in the diagnosis are not repeated.
Body compressed, very deep, especially in specimens <200 mm SL ( Fig. 1 View FIGURE 1 ); anterodorsal profile ascending abruptly, rostro-occipital profile somewhat convex, body slender; orbit diameter large, slightly greater than interorbital width; snout length much greater than orbit diameter; mandibular profile weakly convex; preopercle entire, with rounded angle; preopercular flange fully scaled; first to third dorsal-fin spines becoming progressively longer, third or fourth dorsal-fin spine longest; predorsal length clearly less than body depth, slightly less than dorsal-fin base length; pectoral-fin tip reaching to or beyond first anal-fin spine base, pectoral-fin length greater than head length; anal-fin spines stout, second usually more stout and usually longest, longer than orbit diameter; caudal fin weakly forked, lobes short, broad; most upper gill rakers on lower limb of gill arch long, a few lower gill rakers sometimes abruptly shortened or rudimentary.
Color when fresh —based on color photographs of the holotype ( Fig. 1 View FIGURE 1 B) and the paratype specimens ( Fig. 1 View FIGURE 1 A) for both prior to preservation, and four uncatalogued color photographs (thawed, Fig. 2 View FIGURE 2 for one of the four specimens and fresh; lost after collection but tissue sample, herein analyzed and same in holotype’s sequence data in MUFS 42666) by S. J. Newman: body basically pinkish, becoming whitish in abdomen, upper part of pectoralfin rays somewhat more pinkish colour but pinkish hyaline in lower part; anal-fin membrane and rays pale reddishhyaline; caudal-fin membrane and rays mostly reddish hyaline.
Color of preserved specimens —upper sides of head and trunk uniformly yellowish-tan; life colors lost in others.
Etymology. This species is named in honour of Dr. Kent E. Carpenter (ODU) for his many contributions to ichthyology and activities pertaining to ichthyology at the FAO and IUCN.
Remarks. Nominal species were discussed by Iwatsuki et al. (2007). Dentex spariformis Ogilby, 1910 has long been considered a junior synonym of D. tumifrons (= D. hypselosomus of Iwatsuki et al. 2007; Allen, 1997; Akazaki and Séret, 1999; Carpenter, 2001) but Iwatsuki et al. (2007) regarded Dentex spariformis as a valid species, known only from northern and northeastern Australia, Bali, and Lombok Island, southern Indonesia, and the Arafura Sea (fig. 6 of Iwatsuki et al. 2007), at depths between about 100 and 264 m, being a deeper water inhabitant.
A large photographed fish ( Fig. 2 View FIGURE 2 , about 310 mm SL) was collected from Western Australia before the type material was collected. This specimen was lost but tissue was taken, and the sequence data confirmed that this was a specimen of D. carpenteri n. sp.
The types of D. carpenteri n. sp. were compared with example of the four known valid species from the western Pacific: Dentex abei and D. hypselosomus from the Northern Hemisphere, and D. fourmanoiri and D. spariformis from the Southern Hemisphere, respectively. We concluded that the specimens of the Dentex species from Western Australia are a distinct new species with clear morphological and genetic differences compared with those of the four known species (see different morphology of Comparisons below; Fig. 4 View FIGURE 4. K 2 P ).
Comparisons. Differences between the four species of the Dentex hypselosomus complex ( D. abei , D. fourmanoiri , D. hypselosomus , and D. spariformis ) have been previously discussed ( Iwatsuki et al. 2007). Dentex carpenteri n. sp. is most similar to D. spariformis in overall appearance of the body, but can be differentiated from the above species in having the following combination of characters: posterior margin of upper jaw not reaching to or reaching slightly beyond vertical at anterior margin of eye; often a wide yellow region on the greater part from the snout to the second infraorbital, often also on the upper jaw; suborbital depth greater (9.7–10.9% SL; Fig. 3 View FIGURE 3 A); considerably deeper body in specimens of 100−200 mm SL ( Figs. 1 View FIGURE 1 A −B & 2, vs. posterior hind margin of upper jaw reaching clearly beyond a vertical through anterior margin of eye or reaching beyond a vertical through anterior margin of black iris; no yellow region from snout to infraorbital; suborbital depth shallow (7.6–9.6% SL in D. spariformis ); relatively deep body in specimens of about 100−200 mm SL in D. spariformis ; Fig. 1 View FIGURE 1 B; 50.0– 55.0% SL vs. 45.5–51.4% SL in D. spariformis ).
The orbit diameter in SL measurements of D. carpenteri n. sp. and D. spariformis are almost the same ( Fig. 3 View FIGURE 3 B), but are greater than those of D. abei and D. hypselosomus . Conversely, the suborbital depth of D. carpenteri n. sp. is similar to that of D. hypselosomus ( Fig. 3 View FIGURE 3 A). In addition, we observed that D. carpenteri n. sp. often has a much deeper body than that of D. spariformis in specimens <200 mm SL ( Fig. 1 View FIGURE 1 A −B).
Furthermore, squamation of the anterior margin of head scales is similarly rounded or pointed in Dentex carpenteri n. sp., D. spariformis , D. fourmanoiri and D. abei , but only Dentex hypselosomus has an anterior margin of dorsal head scales that is straight or somewhat concave ( Fig. 5 View FIGURE 5 A–B).
Distribution. Dentex abei , D. fourmanoiri , D. hypselosomus , and D. spariformis have allopatric and nonequatorial distributions in the western Pacific (fig. 5 of Iwatsuki et al. 2007). In the Northern Hemisphere, D. abei is currently known only from the Ryukyu Islands including the Okinawa Trough, and Ogasawara Islands, Japan, northeastern Taiwan, and off Luzon Island, Philippines; D. hypselosomus is currently known from the East Asian Shelf, including southern Japan (except the Ryukyu and the Ogasawara Islands), southern Korea, Taiwan (generally the western part of Taiwan), and China.
In the Southern Hemisphere, Iwatsuki et al. (2007) reported that D. spariformis is distributed off Australia (except the southwestern part), while D. formanoiri is currently known only from off New Caledonia, including the Chesterfield Islands. The type specimens of D. carpenteri n. sp. were collected at depths between 200 - 215 m, and appear to be a much deeper inhabitant than the other species ( Iwatsuki et al. 2007).
Although we could not specify the exact distributional boundary between D. carpenteri n. sp. and D. spariformis , the distribution pattern of these species may follow a similar pattern to that of the deepwater congeneric trichiurids ( Burhanuddin et al. 2002; Burhanuddin and Iwatsuki 2003; Chakraborty et al. 2005) in the Southern Hemisphere. For example, Trichiuris nikolensis in northwestern Australia, and T. australis in northeastern Australia, exhibit a boundary zone around the Gulf of Carpentaria. Species in both Dentex and Trichiurus are known to be basically benthic and deep-sea inhabiting species strongly connected with a continental shelf ( Nakamura and Parin 1993 for Trichiurus ) and are not strong swimmers (e.g. like species of the carangid genus Seriola ). They may therefore exhibit similar distributional boundaries.
In the Northern Hemisphere, Dentex species have a clearly allopatric distribution in the East Asian Shelf ( D. abei and D. hypselosomus ), while in the Southern Hemisphere, an allopatric distribution may also be evident ( D. carpentari n. sp. from Western Australia, D. spariformis from northern and northeastern Australia and D. fourmanoiri from New Caledonia; fig. 5 of Iwatsuki et al. 2007 and this study).
Mohsin and Ambak (2010; plate 270) recently reported Dentex angolensis from Sabah, Borneo (off Kota Kinabalu and Sandakan), Malaysia (M. A. bin Ambak pers. comm.). Dentex angolensis is known to be distributed in the Eastern Atlantic, and is unlikely to be an Indo-Pacific species. The reported specimen may be D. carpenteri n. sp. based on the posterior margin of the upper jaw in the photographed specimen clearly not extending beyond a vertical at the anterior margin of the eye. The Malaysian specimens of Dentex need further examination.
Phylogenetic analysis. The phylogenetic tree (mitochondrial 16S ribosomal RNA genes, 545 bp) has the clade of Dentex carpenteri n. sp. included in a clade of D. spariformis and D. fourmanoiri , all of the Southern Hemisphere. As this clade includes D. carpenteri n. sp., it clearly demonstrates that each species is distinct ( Fig. 4 View FIGURE 4. K 2 P ). In overall body appearance D. carpenteri n. sp. is most similar to that of D. spariformis , which is known from off northern and eastern Australia (Northern Territory, Queensland, and New South Wales; fig. 5 of Iwatsuki et al. 2007).
Chiba et al. (2009; figs. 2−3) reported a gene tree for eastern Atlantic Dentex species ( Dentex angolensis , D. maroccanus , D. macrophthalmus , plus Spicara alta ) and westen Pacific Dentex species ( D. abei , D. hypselosomus , and D. spariformis ), with different clades within the same monophyletic group, supporting a previous study of the two sparids ( Dentex hypselosomus and Spicara alta ) by Orrell & Carpenter (2004). However, Dentex dentex , D. gibbosus , D. canariensis , plus Cheimerius nufar were more distinct and formed a different clade in the tree of Chiba et al. (2009; figs. 2−3). Discernible morphological differences in the cranium among the three groups has been detected (Tanaka and Iwatsuki pers. obs.), and further work on the generic definition of the genera, Dentex , Cheimerius , and Spicara is urgently required so as to understand interrelationships within these Sparidae . A new insight on sparid phylogeny is possible if all known species can be examined and further genetic analysis of complete mitochondrial DNA and greater regions of nuclear DNA are undertaken, in association with detailed morphological studies, which included all previous sparid work ( Akazaki 1962; Carpenter and Johnson 2002; Day 2002).


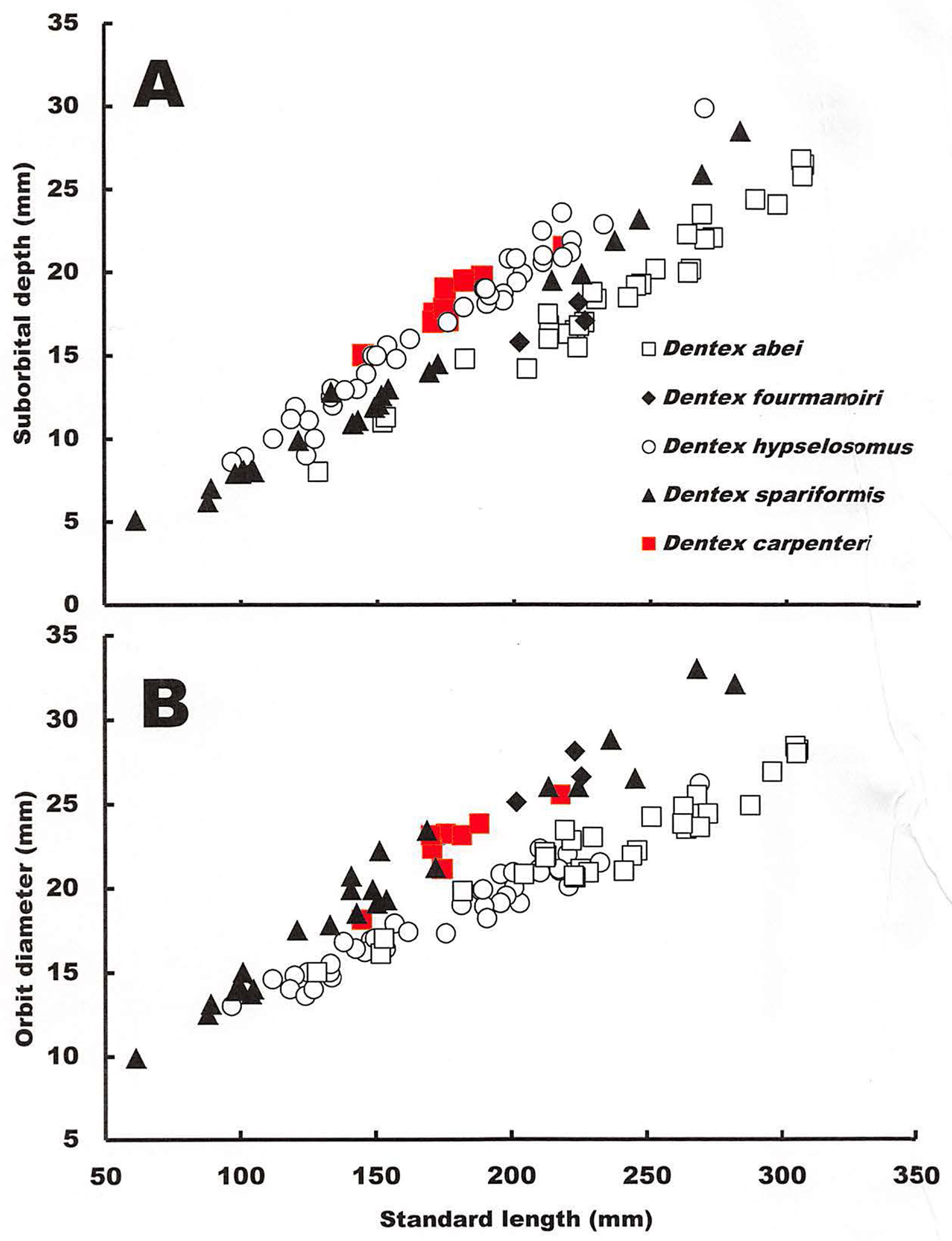
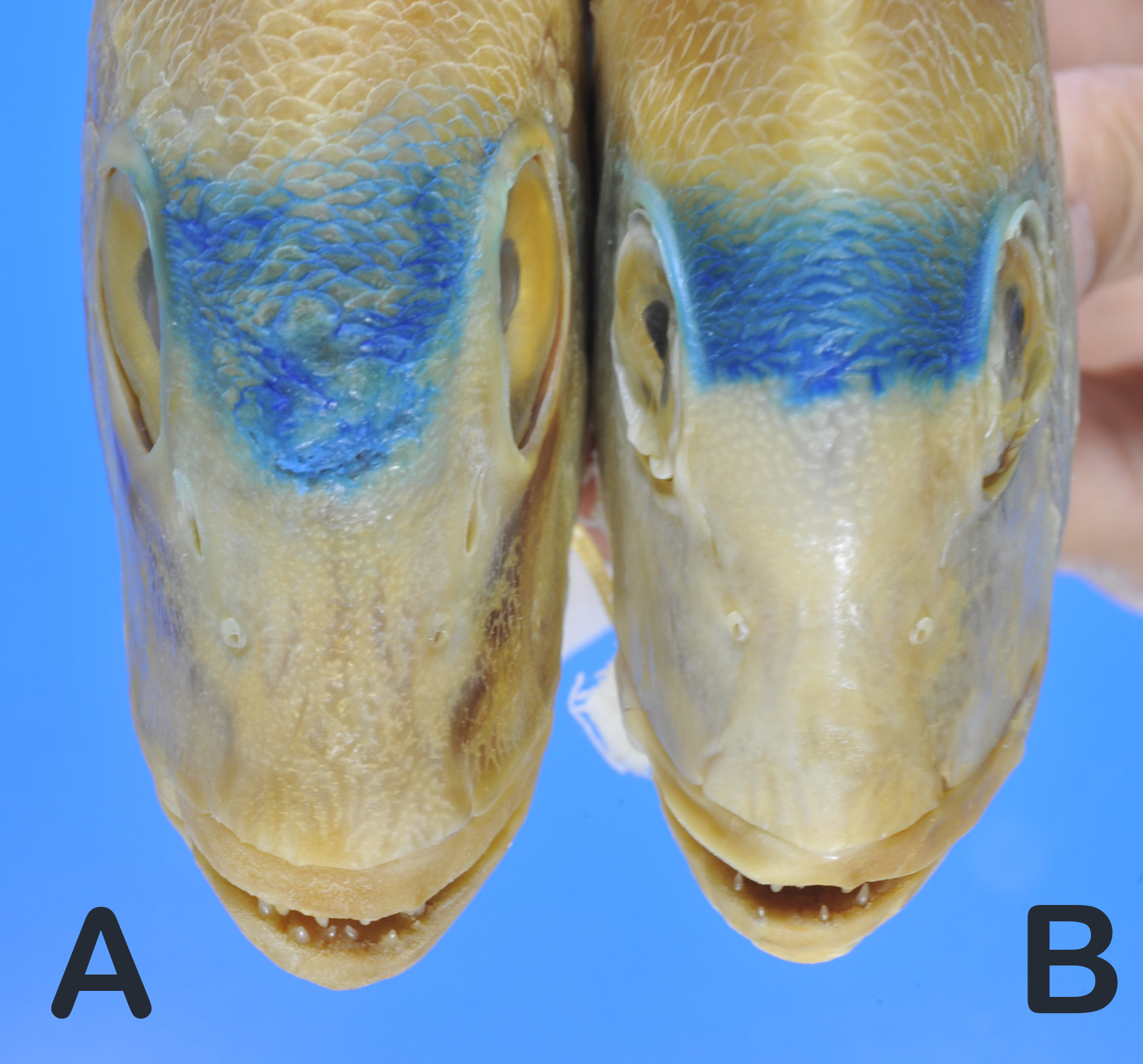
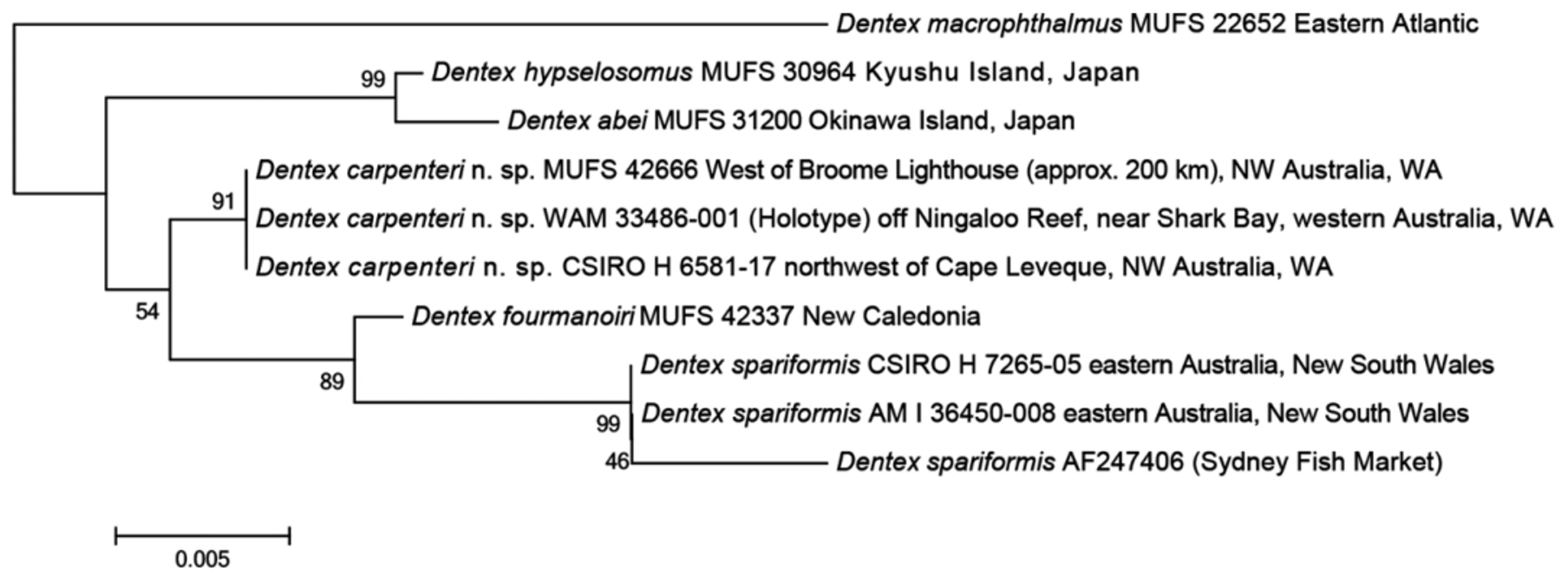
TABLE 1. Counts and proportional measurements of type and non-type specimens of Dentex carpenteri n. sp., expressed as percentages of standard length (means in parentheses).
| Holotype WAM P.33486-001 n = 1 | Paratypes WAM P.33490-001 n = 8 | |
|---|---|---|
| Dorsal and anal fin rays Pelvic and pectoral fin rays Pored lateral line scales | XII, 10; III, 8 I, 5; i+14 47 | XII, 10; III, 8 I, 5; i+14 47−49 |
| Scale rows above and below lateral line Scales between 5th dorsal fin spine base and lateral line Squamation on preopercular flange | 6 / 15 4 ½ fully scaled | 6 / 14−15 4 ½ fully scaled |
| Gill rakers Standard length (mm) Body depth | 7+1+11 = 19 219 53 | 8−9+1+10−11 = 19−21 145−189 50−55 |
| Head length Body width at pectoral fin base Snout length | 36 18 15 | 36−40 15−19 15−17 |
| Orbit diameter Dermal eye opening Suborbital depth | 12 10 10 | 12−14 10−11 10−11 |
| Interorbital width Upper jaw length Caudal peduncle depth | 10 15 12 | 9−11 13−15 12−13 |
| Caudal peduncle length Predorsal length Preanal length | 21 47 66 | 18−22 46−51 62−67 |
| Prepelvic length Dorsal-fin base Anal-fin base | 41 55 20 | 36−42 51−55 18−20 |
| Pelvic-fin spine length First pelvic-fin ray length Pectoral-fin ray length | 17 25 40 | 14−18 23−27 39−43 |
| First dorsal-fin spine length Second dorsal-fin spine length Third dorsal-fin spine length | 4 (broken) 6 (broken) 5 (broken) | 6−8 10−14 15−17 |
| Fourth dorsal-fin spine length First dorsal-fin ray length12 First anal-fin spine length | 16 12 8 | 15−22 10−13 6−9 |
| Second anal-fin spine length Third anal-fin spine length First anal-fin ray length | 12 12 11 | 12−15 10−12 11−13 |
| WAM |
Western Australian Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Dentex carpenteri
| Iwatsuki, Yukio, Newman, Stephen J. & Russell, Barry C. 2015 |
Allotaius spariformis:
| Hutchins 2001: 35 |
Dentex tumifrons
| Carpenter 2001: 3000 |
| Allen 1997: 132 |