
Myzocallis ( Myzocallis ) macrolepidis, Barbagallo & Massimino Cocuzza, 2022
|
publication ID |
https://doi.org/10.11646/zootaxa.5183.1.15 |
|
publication LSID |
lsid:zoobank.org:pub:9C30ACCC-9B0D-459A-A490-9DE99000EF31 |
|
DOI |
https://doi.org/10.5281/zenodo.7075116 |
|
persistent identifier |
https://treatment.plazi.org/id/9F3787E3-103B-FFC6-9DC8-3EE1FD5AF87E |
|
treatment provided by |
Plazi |
|
scientific name |
Myzocallis ( Myzocallis ) macrolepidis |
| status |
sp. nov. |
Myzocallis ( Myzocallis) macrolepidis sp. n.
General diagnosis
Body pale yellow in alate morph, with rather small and weakly pigmented spinal and marginal abdominal sclerifications. Head and thorax moderately pigmented all around, with a narrow brown line on anterior sides of prothorax. Antennae about ¾ of body length and pale, with apical part of III–V and the sensillar area of VI joints brown pigmented. Wings hyaline, with a part for a small brownish area at bases of pterostigma and the base of cubital veins in fore wings. Legs mostly pale, with slightly brownish distal part of tibiae and tarsi. Siphunculi short, truncate conical and rather pale. Cauda pale, short and knobbed. Alatoid nymphs paler than alates, with numerous and capitate pale setae on dorsum, in paired groups of spinals and marginals; antennal joints pigmentation similar to that of alates.
Morphology
Alate viviparous female ( 28 specimens examined), Fig. 1a–h View FIG and Fig. 2a View FIG ; Tab. 2 View TABLE 2
A delicate and rather small aphid species, 1.05–2.10 mm in body length. Colour in life pale yellow with red eyes. Body pigmentation given by two marginal narrow brown stripes on prothorax and two paired abdominal series of spinal and marginal brown patches; these latter are moderately sclerified and show a finely spiculated cuticle, mainly evident on marginal spots. The eighth urotergite with a narrow transverse browning band. Dorsal body hairs all thick and more or less distinctly capitate or rarely blunt, as sometimes they are on urotergites posterior to siphunculi, including the 8 th urotergite. Ventral body hairs all finely acute apically (26–34 µm long on 3 rd urosternite). One brachypterous available specimen has dorsal body hairs even strongly capitate and slightly longer compared to normal winged specimens.
No. 1 is the holotype; ns. 2–16 are paratypes. All specimens collected on Q. macrolepis at Tricase ( Lecce province ), Italy; ns. 1,12: 3.XI.90; ns. 2–5, 11, 13, 14: 6. V.91; ns. 6–9, 15: 10. V.18; ns. 10, 16: 11. V.18. Abbreviations : Bl = body length; Al = antennal length; Ajl = III– VI antennal joints length; Rhin. III = no. of secondary rhinaria on III antennal joint; Urj = ultimate rostral joint length; IIht = second hind tarsomer length; Sl = siphuncular length .
Head with a sinuate frontal profile, due to the low antennal tubercles and the median prominence bearing the third ocellus. Frontal hairs, mostly 24–32 µm long, rarely up to 55 µm as seen in one or two specimens. Dorsal chaetotaxy performed by two couples of anterior discals, placed on small wart-like elevations, and three pairs of posterior (occipital) hairs. All discal hairs are as a rule thicker than frontals, 26–40 µm long or 1.6–2.5 times the basal articular diameter of III antennal joint (IIId).
Antennae 0.66–0.98 of body length, pale in colour, except for the brown distal part of III–V joints and the sensillar area of VI. Joint III 1.00–1.60 (most frequently 1.10–1.30) of the distal part of VI joint. The latter has a processus terminalis 2.00–2.30 (on average 2.10, n=26) times its basal part. The 1–4 rounded and finely dotted secondary rhinaria are placed on 0.25–0.55 basal part of III antennal joint (IIIa); their diameter is 0.45–0.90 times of IIIa at the point of their insertion. Primary rhinaria regularly ciliated; the main one on VIa is 1.10–1.25 times the diameter of that on Va. First and second antennal joints bearing respectively 3 (rarely 4) and 2 hairs, the inner one of which is more or less distinctly capitate, 12–20 µm long or 0.75–1.25 times IIId. Hairs on antennal flagellum all short and acute at apex; those on IIIa are 4–5 in number and 7–14 µm long or 0.48–0.75 times IIId.
Rostrum 0.30–0.45 mm, pale with darkish apex and reaching the middle of mesosternum. Last rostral joint (urj) acute conical in shape, 0.110 –0.155 mm (measured from dorsal rim) and 3–4 times its basal diameter; it bears 5–8 supplementary hairs. Ratio urj/IIht 1.36–1.60 (average of 25 measured specimens, 1.49), and ratio urj/VIb 0.95–1.25 (average of 28 specimens, 1.06).
Thorax with a rather pale integument, except for the two brownish, spiculated small strips on antero-marginal sides of pronotum. Dorsal chaetotaxy of the latter represented by four couples of spinal hairs (18–48 µm long) on the discal area and one marginal pair (16–34 µm) placed on the external sides of posterior lobe. A couple of small pleural hairs (usually not present in other known species of subg. Myzocallis ) is present on anterior part of the pronotum in several specimens. Mesothoracic lobes each bearing about 8–12 mostly capitate or subcapitate hairs up to 30–52 µm long.
Forewing with hyaline membrane and pale venation, except for the brown basal part of Cu1–Cu2 and a small spot at the base of pterostigma, as usual in Myzocallis s. str. The latter, measured from the base of M vein, just behind its brown spot, is 3.50–4.70 times longer than wide. Rs vein complete, albeit faintly evident.
Legs mostly pale, except for the slightly infuscated distal part of femora and tibiae, including tarsi. Forelegs with coxae and femora slightly enlarged, comparatively to middle and hind legs. Distal part of tibiae and tarsi with spiculose cuticle. Hind tibiae about half (0.40–0.60) of body length. Dorsal femoral hairs subcapitate or blunt at apex, 16–25 µm long or 0.40–0.60 of trocantro-femoral suture diameter. Tibiae with their outer hairs blunt or moderately knobbed on basal part and progressively more acute towards their distal part, where they are 16–32 µm long or 0.85–1.40 times the median tibial width. Rastral organ represented by 3–4 spur-like setae, which are thicker in the fore tibiae compared to the other two pairs. First hind tarsomer with 2 dorsal and 5 ventral hairs. Empodial hairs flabellate.
Abdomen with dorsal hairs clustered into double spinal and marginal groups, each bearing 4–5 (6) hairs on 1 st –4 th urotergites and 2–4 on 5 th –7 th urites; their length varies from 10–40 µm (spinals) to 8–28 µm (marginals). The 8 th urotergite bearing a row of 5–8 blunt or subcapitate hairs of 18–36 µm.
Siphunculi truncate and mostly pale or slightly infuscated towards apex, with nearly smooth cuticle. They are 0.055 –0.070 mm long and subequal to or just longer (0.90–1.20) of their basal diameter.
Cauda pale, with its knobbed distal part shorter (0.75–0.90) than wide. Caudal hairs 9–12, of which four longer ones placed on the distal border and the remaining on the ventral side of the knob part of cauda, as usual in this aphid group.
Anal plate deeply indented, with 7–10 long hairs for each of the two lobes. Genital plate with 6–10 hairs along the distal margin and about 8–10 on its discal part. Gonochaetae in two groups of 4–6 small hairs each.
Fundatrix ( 1 specimen examined)
One alate viviparous with a comparatively shorter processus terminalis (collected in early May), is here tentatively regarded as a fundatrix, though not discarding the possibility of being it a fundatrigenia specimen of 2 nd generation. Its main difference (see Tab. 2 View TABLE 2 , specimen no. 11) from summer generation specimens is the lower ratio of antennal processus terminalis against the basal part of VI antennal joint (1.69 vs. 2.00–2.30); in addition, the same specimen has a slightly longer ultimate rostral segment (ratio urj/IIht = 1.64 vs. 1.36–1.60 of summer fundatrigeniae), which bears 8 supplementary hairs. No other significant morphological divergences within standard fundatrigeniae have been detected.
Embryo ( 10 specimens examined)
The embryo has a chaetotaxy protopattern of well knobbed long marginal and spinal hairs. The latter pairs are all mesially aligned and shortened on prothorax, 1 st and 5 th urites, as in all other known taxa of subgenus Myzocallis . Those on 1 st and 5 th urotergites are 15–20 µm and 16–25 µm, respectively; the formers are subequal to or not much longer (0.94–1.20) than diameter of siphuncular pori, which is 15–19 µm. Spinal hairs on 2 nd, 3 rd and 4 th urotergites are clearly longer, being respectively 40–54 µm, 46–55 µm and 50–78 µm, equivalent to 0.95–1.20 (those on 2 nd –3 rd urotergites) up to 1.20–1.70 (on 4 th urite) times the distance between their bases on respective abdominal segments.
Alatoid nymph ( 20 specimens examined), Fig. 1i–j View FIG and Fig. 2b View FIG ; Tab. 2 View TABLE 2
Body length 0.95–1.40 mm. Colour in life very pale yellow, with light brown spinal and marginal sclerifications. These pigmented patches tend to discolour during the specimen maceration process and therefore they can become hardly detectable on slides mounted material. Antennae pale, except for the brown distal part of III–V joints and the sensillar area of VIa. Rostrum, legs, and siphunculi pale.
Dorsal body hairs stout and well knobbed apically; the diameter of the main hairs distal knob is on average equivalent to or wider (1.2–1.3, rarely up to 1.4) than the basal width of the same hairs. Cephalic and pronotal hairs are all similar in length: frontals 60–110 µm, discals 65–115 µm, pronotals 65–125 µm (hairs length is inclusive of their basal papilla). Hairs on meso-metathorax and abdominal segments anterior to siphunculi clustered into spinal and marginal groups of (3)4–5 hairs each. Urotergites 6 th –7 th bearing 2–3 hairs only for each group, occasionally with a reduction to a single marginal hair on 7 th urotergite. Eighth urotergite with a transverse row of 3–5 dorsal hairs. The length of all these dorsal body hairs consistently varies within each group from about 10–50 µm of smaller ones to a maximum of 70–160 µm, either as spinals and marginals; the latters are on average slightly longer than the formers. In addition, smaller pleural hairs (1–2 for each side) are usually present from first to fifth urotergites; their length varies from 10–12 up to 30–35 µm.
Head with frontal profile moderately convex. Antennae usually shorter (0.70–0.95) or rarely a little longer (up to 1.10 in some smaller specimens) than the body. Joint III shorter (0.68–0.92) than processus terminalis of VI joint; the latter is 1.90–2.36 (mostly not less than twice) times the basal part of the same antennomer. First and II antennal joints each bearing a couple of capitate hairs, of which the inner one on II joint (35–53 µm long or 1.80–2.74 times the diameter of IIId) is usually just longer than the correspondent hair on I joint. The III antennal segment bears as a rule one capitate hair (12–30 µm long or 0.60–1.48 of IIId), placed at its inner third basal part; a second and much smaller, subcapitate or blunt hair (6–10 µm long) can be only occasionally present more apically on the same joint. In addition, a few inconspicuous and acute hairs (5–10 µm long, or 0.26–0.50 of IIId) are regularly distributed on flagellar joints.
Rostrum ventrally reaching the middle of mesothorax; its last joint is very similar to that of alate viviparous either in shape and size, 1.34–1.54 (but rarely less than 1.40) times the IIht and 1.02–1.32 of VIb; it bears 5–7 supplementary hairs likewise that of alate morph.
Legs with dorsal femoral hairs more or less knobbed and up to 22–35 µm long, or 0.50–0.75 of the trochantrofemoral suture on hind legs. Tibiae with outer hairs rather knobbed or blunt on their basal part, becoming finely pointed towards the distal part of the same tibiae; these latter hairs have a length of 32–55 µm and are 1.10–2.20 times the median tibial diameter. Hind tarsus with 5 ventral hairs. Distal part of tibiae and tarsi regularly spinulated.
Siphunculi truncated, with a smooth cuticle; they are 0.034 –0.048 mm long, being subequal in length (0.80– 1.10) to their basal width.
Types
As holotype is selected an alate viviparous female collected on Quercus macrolepis Kotschy at Tricase ( Lecce province), Italy, ( 39.927885N / 18.378744E, 70 m a.s.l.) GoogleMaps , now stored in the collection of the senior co-author ( S.B.) at the Dept. of Agriculture , Food and Environment, University of Catania ( Italy). Paratypes are ns. 28 viviparous alatae, and ns. 20 alatoid nymphs (4 th immature instar); all of them were collected on the same host plant species in the common area of Tricase, as the holotype, during the years 1990–92 and in 2018 by co-authors. Paratypes are stored in the collection of the senior co-author as the holotype and part deposited at the Natural History Museum of London (UK) and the collection of Prof. J.M. Nieto Nafria, Department of Biodiversity and Environment, University of León ( Spain) .
Molecular analysis
All species were successfully sequenced, obtaining DNA fragments of 623 bp in length with a base composition of A=34.7%, C=15%, T=40.3 and G=10%. Among Myzocallis species, 541 sites were conserved and 82 variables, of which 49 parsimony-informative. The interspecific genetic distances (expressed as percentage of sequence divergence) among all Myzocallis samples is on average 5.5%, ranging from 2.3% (between M. schreiberi and M. glandulosa ) to 8.0% (between M. schreiberi and M. occidentalis ). The mean genetic distance of M. macrolepidis from the other congeneric species is shown in Tab. 3 View TABLE 3 . These distances are graphically reported in the NJ tree ( Fig. 4 View FIG ) on which the species are subdivided into two main groups. One cluster joints together M. carpini , M. coryli , M. occidentalis , and more separately, M. boerneri . In the other group are included M. macrolepidis , distinct from M. glandulosa and M. schreiberi , that cluster together. M. macrolepidis showed a significant genetic distance comparatively to all other species of the subgenus, with the lowest value of 2.3% recorded against M. schreiberi , which is the most genetically related taxon.
Etymology The specific name of the new aphid taxon derives from that of its host plant species.
Ecology and distribution
The aphid described has been collected so far only on Valonia oak ( Q. macrolepis ) in the Apulia region, southern Italian peninsula (see ‘Types’ for details). The insect life cycle remains unknown at present, though very probably the aphid is a monoecious and holocyclic species, as most of its congeneric taxa are. That oak has an eastern Mediterranean distribution, which involves a very restricted area of S.E. Italy, Albany, and Greece (a ‘trans-ionian’ distributed oak species) with extension to the Middle East. It belongs, taxonomically, to the group (or ‘section’) of Cerroid oak species sensu Denk & Grimm (2010) and, as above mentioned, is strictly allied to Q. ithaburensis (the main host plant species of M. glandulosa ). M. macrolepidis could possibly have a coincident distribution within its host plant oak.
The aphid is not myrmecophilous and lives on the underside of leaves, sometimes in mixed colonies with the congeneric M. ( Pasekia) komareki (Pašek) . Nevertheless, the latter can be easily separated (use an 8x pocket lens) by its deeper yellow body colour and the darker, rather large abdominal dorsal sclerifications, which are also bordered by a darker pigmented ring on spinal ones.
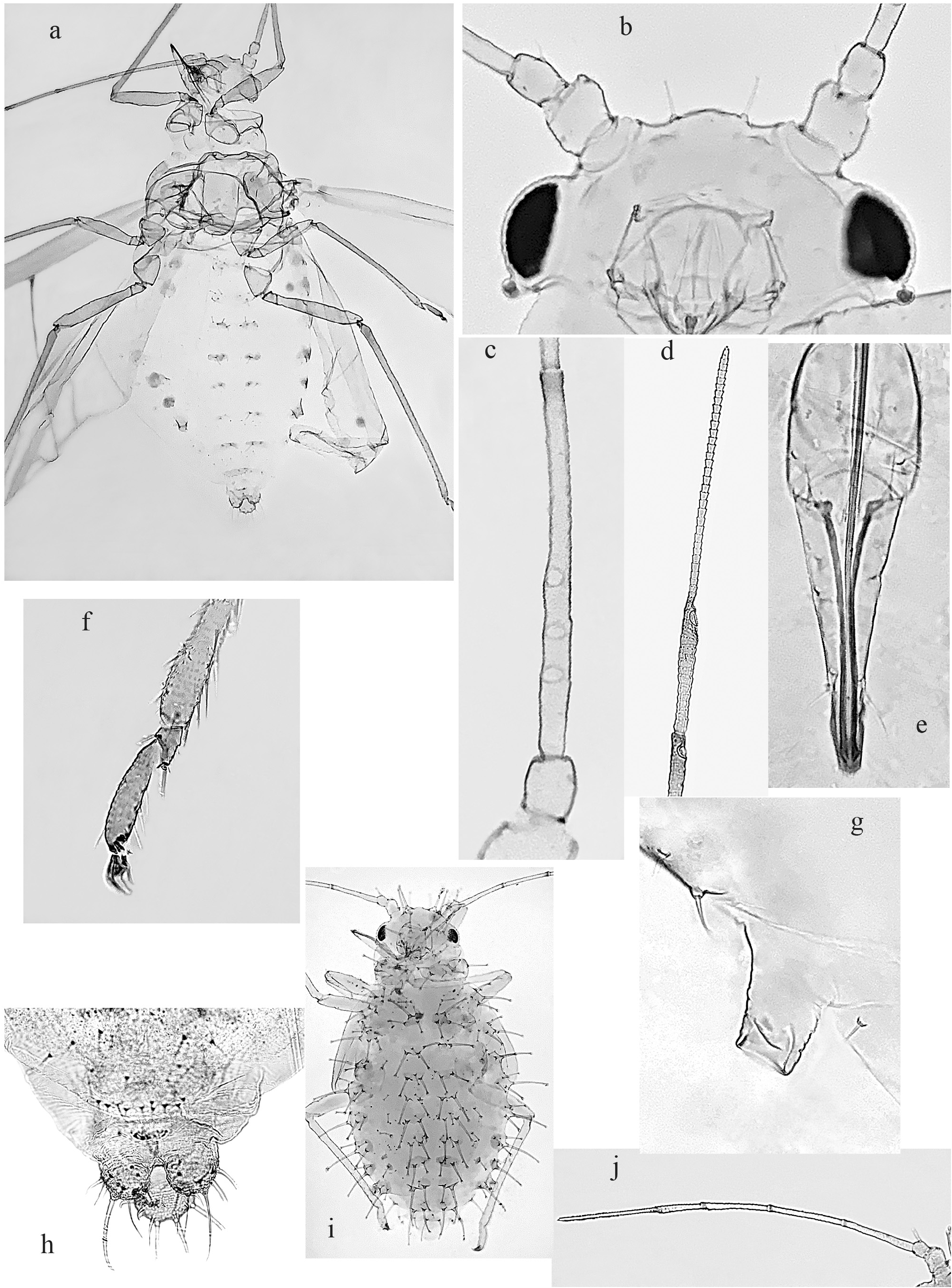
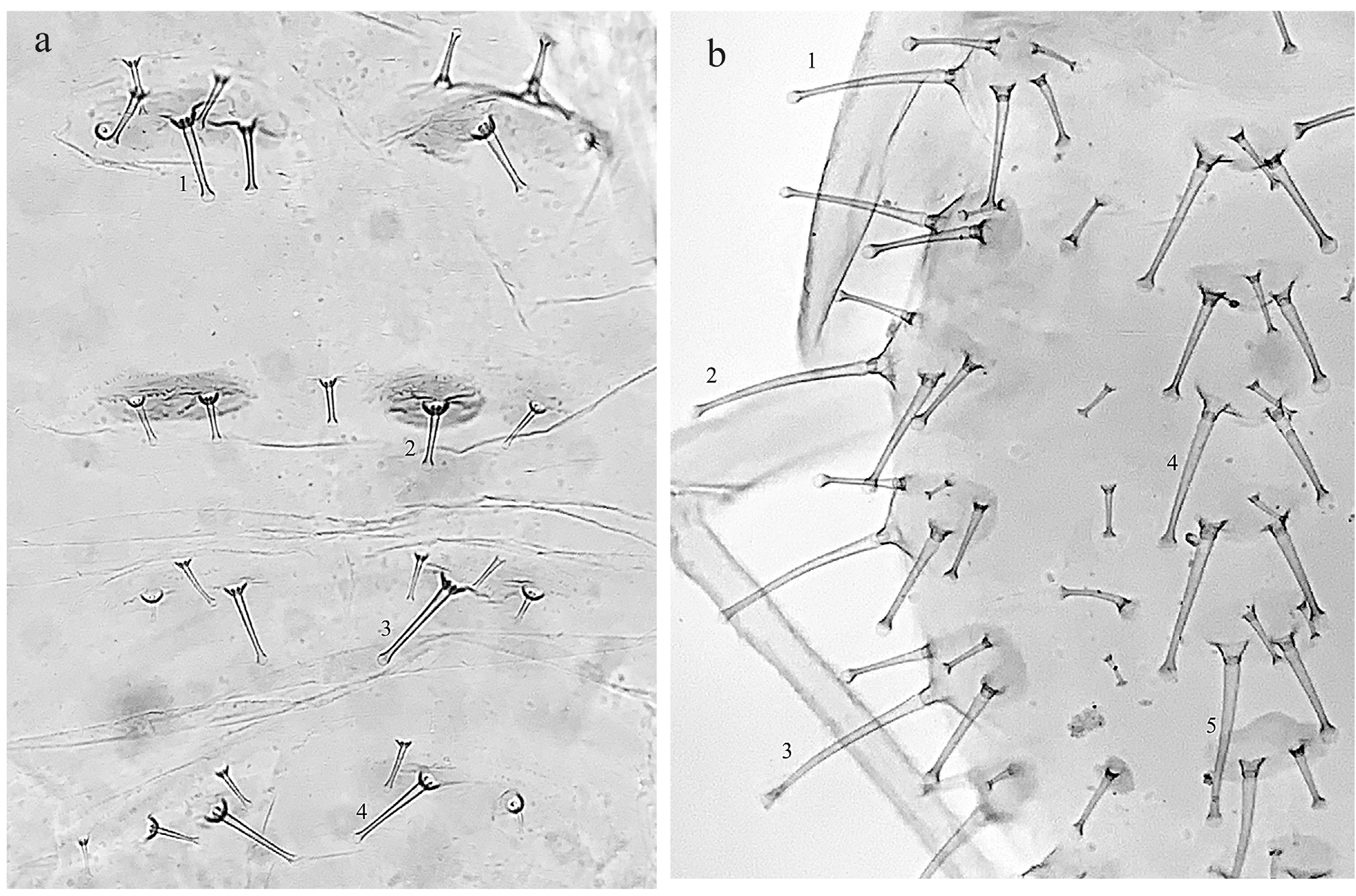
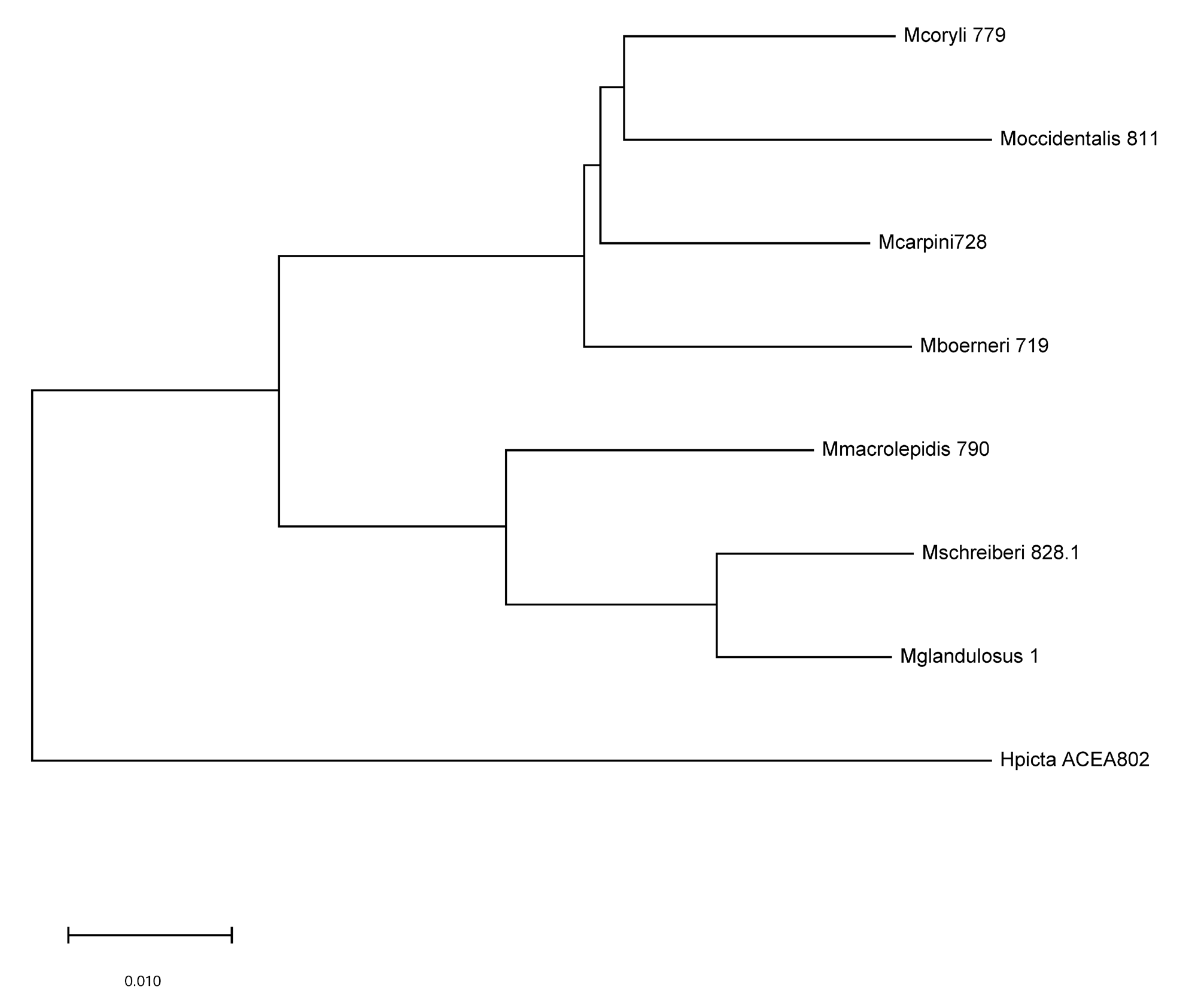
TABLE 3. P-distance and nucleotide divergences (expressed as percentage) of Myzocallis spp.
| V |
Royal British Columbia Museum - Herbarium |
| VI |
Mykotektet, National Veterinary Institute |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |