
Lepidocyrtus thoracicus Mateos, 2023
|
publication ID |
https://doi.org/10.11646/zootaxa.5270.2.7 |
|
publication LSID |
lsid:zoobank.org:pub:B1AF056D-D785-47DD-8EA0-D7E223267B44 |
|
DOI |
https://doi.org/10.5281/zenodo.7865146 |
|
persistent identifier |
https://treatment.plazi.org/id/B3DF5C60-D0AC-4611-804C-5789C76C137D |
|
taxon LSID |
lsid:zoobank.org:act:B3DF5C60-D0AC-4611-804C-5789C76C137D |
|
treatment provided by |
Plazi |
|
scientific name |
Lepidocyrtus thoracicus Mateos |
| status |
sp. nov. |
Lepidocyrtus thoracicus Mateos sp. nov.
Figs 3–25 View FIGURE 3 View FIGURE 4 View FIGURES 5–9 View FIGURES 10–11 View FIGURES 12–14 View FIGURE 15 View FIGURES 16–17 View FIGURES18–20 View FIGURES 21–24 View FIGURE 25
urn:lsid:zoobank.org:act:
Type material. Holotype: Female on slide code CRBA-102730, Padevena, Veneto ( Italy), 920 m above sea level, lat/long coordinates N46.04116 E11.84655, collected using a modified leaf blower functioning as an aspirator (diameter of opening 14 cm), 04.v.2015, leg. B. Zhang. GoogleMaps Paratypes: 18 specimens on slides ( 5 female, 2 juvenile and 11 without visible sexual plate) and 11 specimens preserved in absolute alcohol, same data as holotype. Four paratypes prepared on slides were previously used in the molecular analyses GoogleMaps . Holotype and paratype female slide code CRBA-102731 saved in the collection of the Centre de Recursos de Biodiversitat Animal , Faculty of Biology, University of Barcelona (http://www.crba.ub.edu); other paratypes kept in the E. Mateos’ collection (lot LP386 ) .
Diagnosis. With dark blue pigment on Abd.II–III, lateral dot on Abd.IV, and coxae I–II. Th.II slightly projecting over the head. Ant.I–III, legs, ventral tube, and posterior region of manubrium with scales. Labial chaetotaxy M1M2REL1L2, R shortened. Dorsal cephalic and body macrochaetae formula A0[A2a]A2A3Pa5/10/0101+3. Abd.IV without chaeta s. Unguiculus truncate and with finely serrated outer margin.
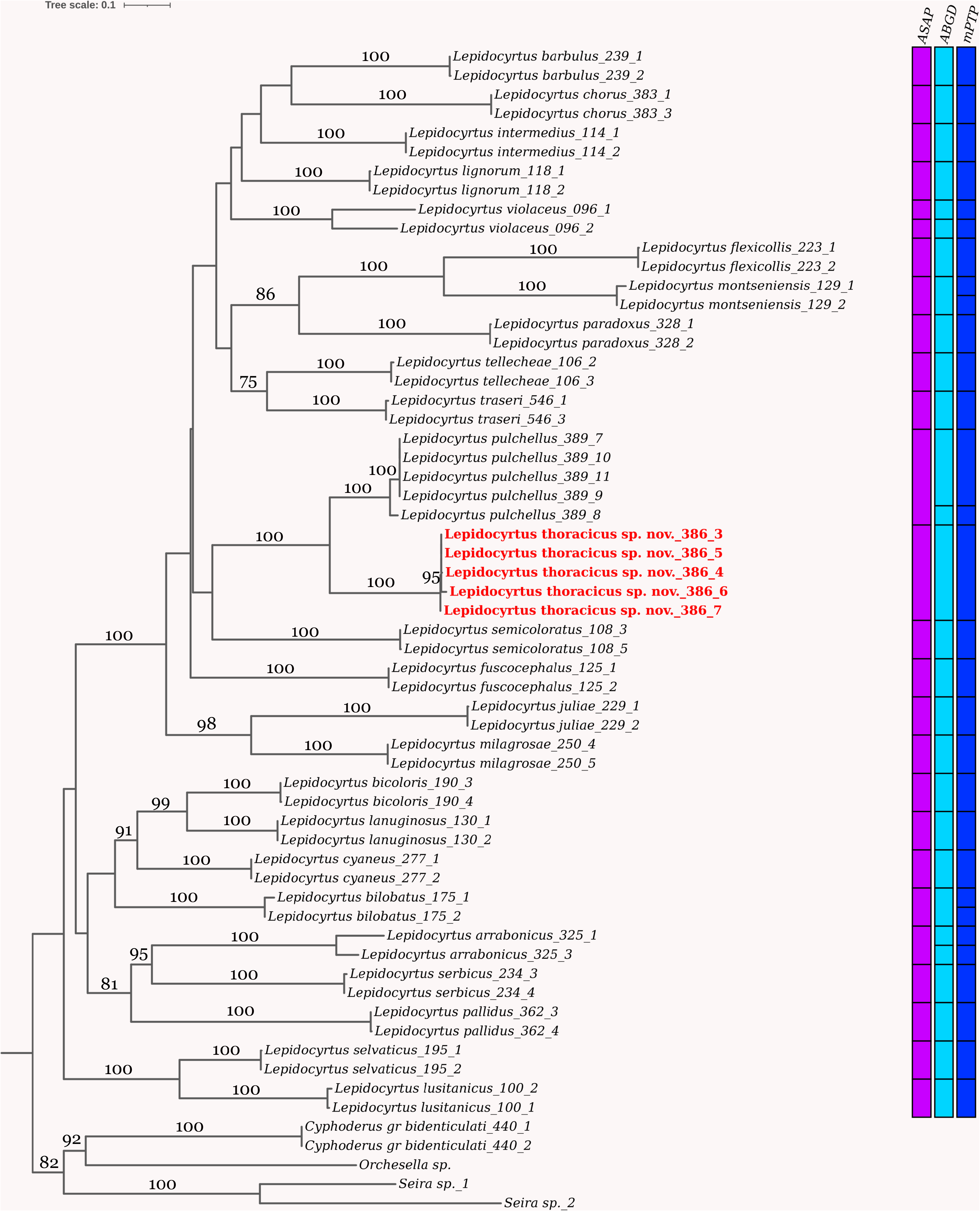
Molecular diagnosis. This species includes all populations that cluster with Cox2 and EF sequences of the individuals LP386-1 to LP386-5 ( Table 1), with significant support in an adequate molecular delimitation model.
Etymology. The species name refers to the presence of a pair of dorsal macrochaetae on mesothorax.
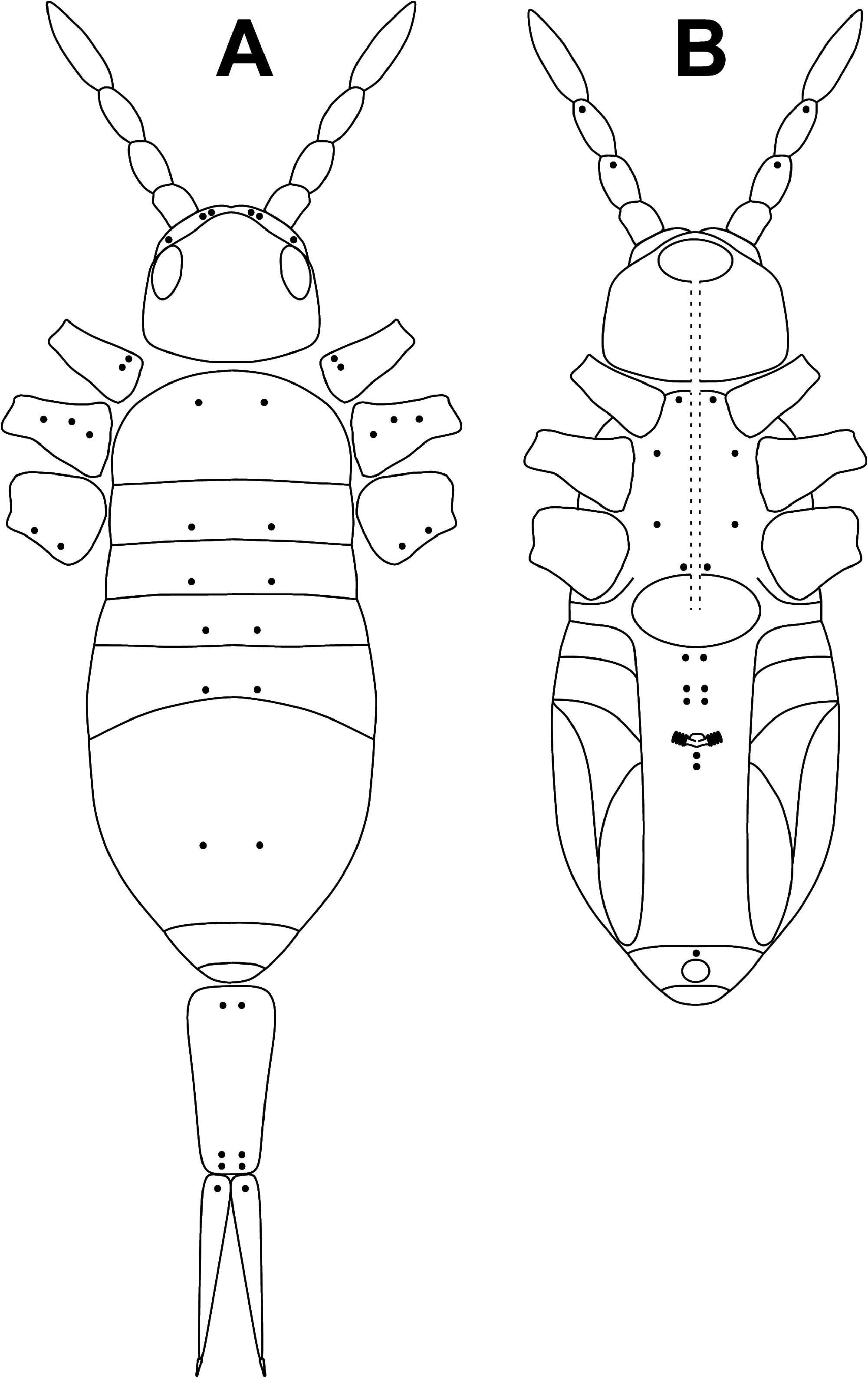
Description. Holotype body length (without head nor furca) 1.1 mm, paratypes 1.0– 1.3 mm. Body color pattern ( Fig. 3 View FIGURE 3 ) with dark blue pigment on Abd.II–III, lateral dot on Abd.IV, and coxae I–II; Ant.II-IV slightly pigmented; densely black pigmented ocular areas. Mesothorax slightly projected over the head.
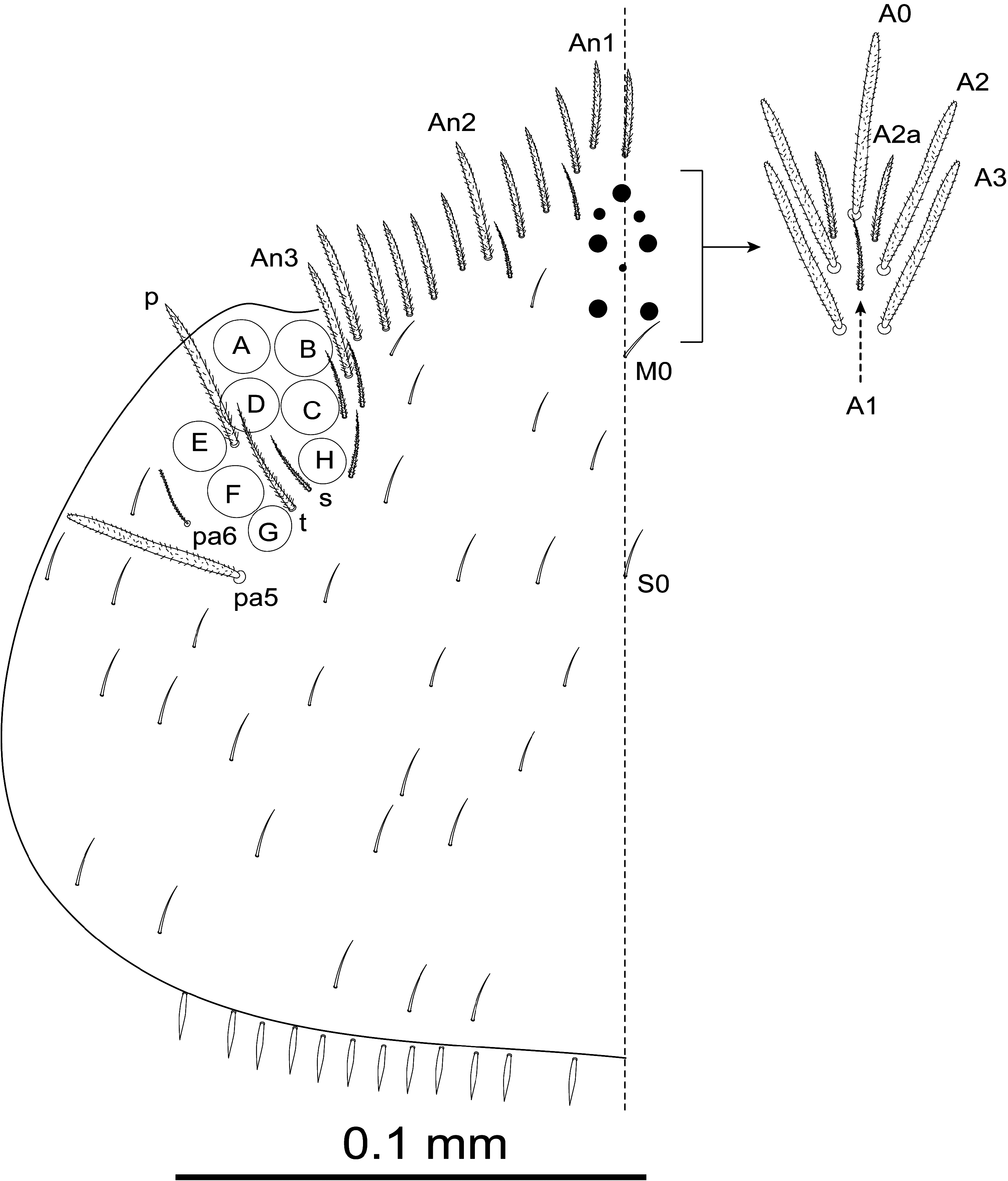
HEAD. Eyes 8+8; eyes A to F subequal, G and H slightly smaller, ratio F/G and C/H ≈ 1.6 ( Fig. 4 View FIGURE 4 ).
Dorsal cephalic macrochaetae A0, A2, A3, Pa5, with pair of smaller supplementary macrochaetae A2a between A0 and A2; maximum number of macrochaetae An on head 11+11. Interocular chaetotaxy with s, t, p ciliated chaetae and 2–3 scales ( Fig. 4 View FIGURE 4 ).
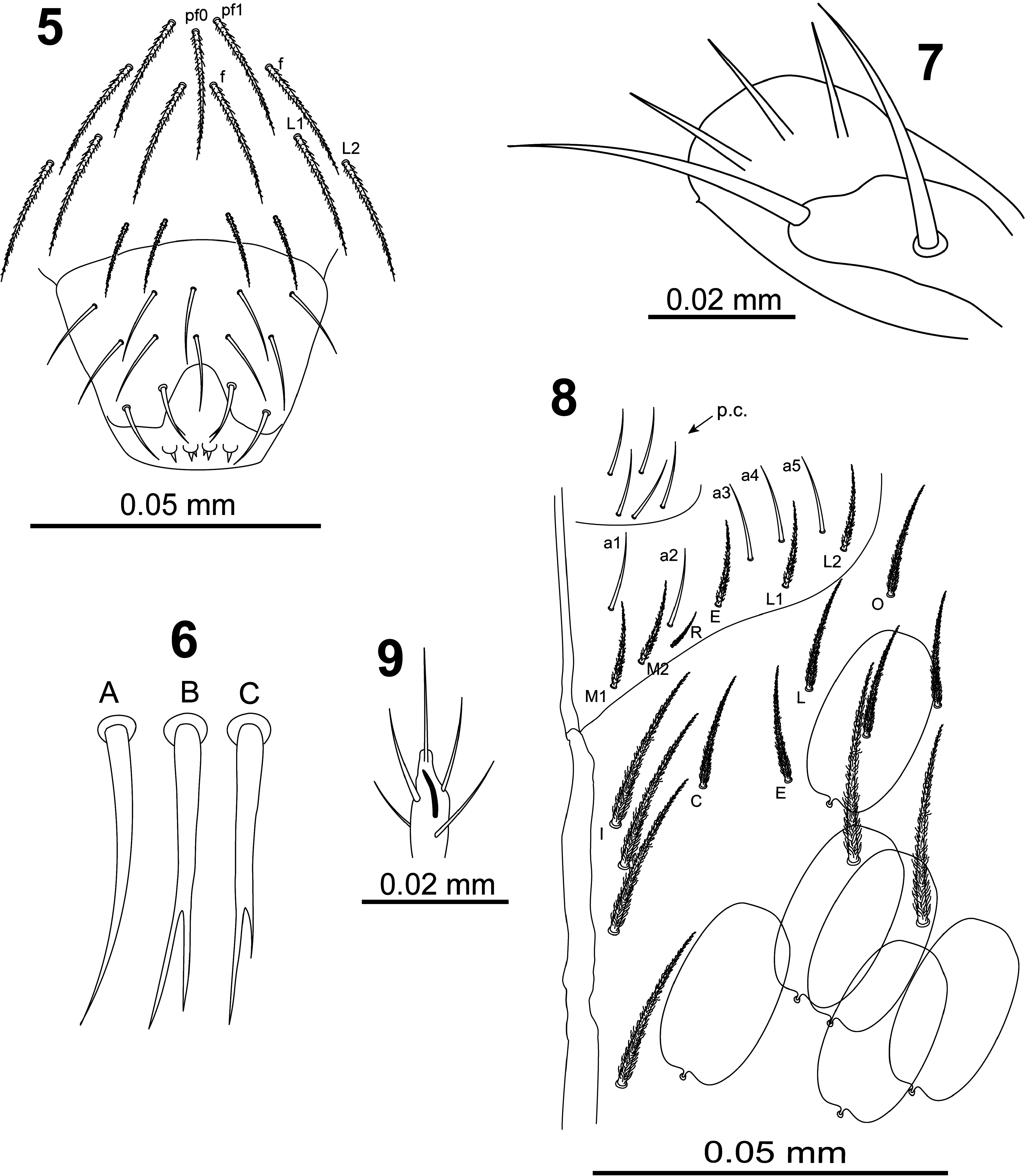
Clypeus ( Fig. 5 View FIGURES 5–9 ) with three prefrontal chaetae (1 pf0 and 2 pf1), four facial chaetae ( f), and four lateral chaetae (2 L1 and 2 L2), all these chaetae ciliated.
Labrum ( Fig. 5 View FIGURES 5–9 ) with prelabral and labral chaetae in typical number 4/554, prelabral chaetae ciliated, first and second rows of labral chaetae smooth, lateral chetae of apical row curved and pointed ( Fig. 6-A View FIGURES 5–9 ), medial chaetae bifurcated ( Fig. 6-B, C View FIGURES 5–9 ); labral intrusion as an inverted U; four rounded labral papillae with 1–3 small spines.
Maxillary palp outer lobe ( Fig. 7 View FIGURES 5–9 ) with smooth apical appendage and basal chaeta, sublobal plate with three smooth appendages and a minute distal process.
Labial and postlabial chaetotaxy as in Fig. 8 View FIGURES 5–9 ; with five smooth proximal chaetae at base of labial palp; labial anterior row with five smooth chaetae ( a1–a5); posterior row formula M1M2REL1L2 all ciliate; chaeta R shortest, ratio M2/R = 2–2.3; postlabial chaetaxy with all chaetae ciliated, row I (along ventral cephalic groove) with four chaetae. Lateral process of outer labial papilla ( Fig. 9 View FIGURES 5–9 ) finger-shape, slightly curved, not reaching apex of papilla.
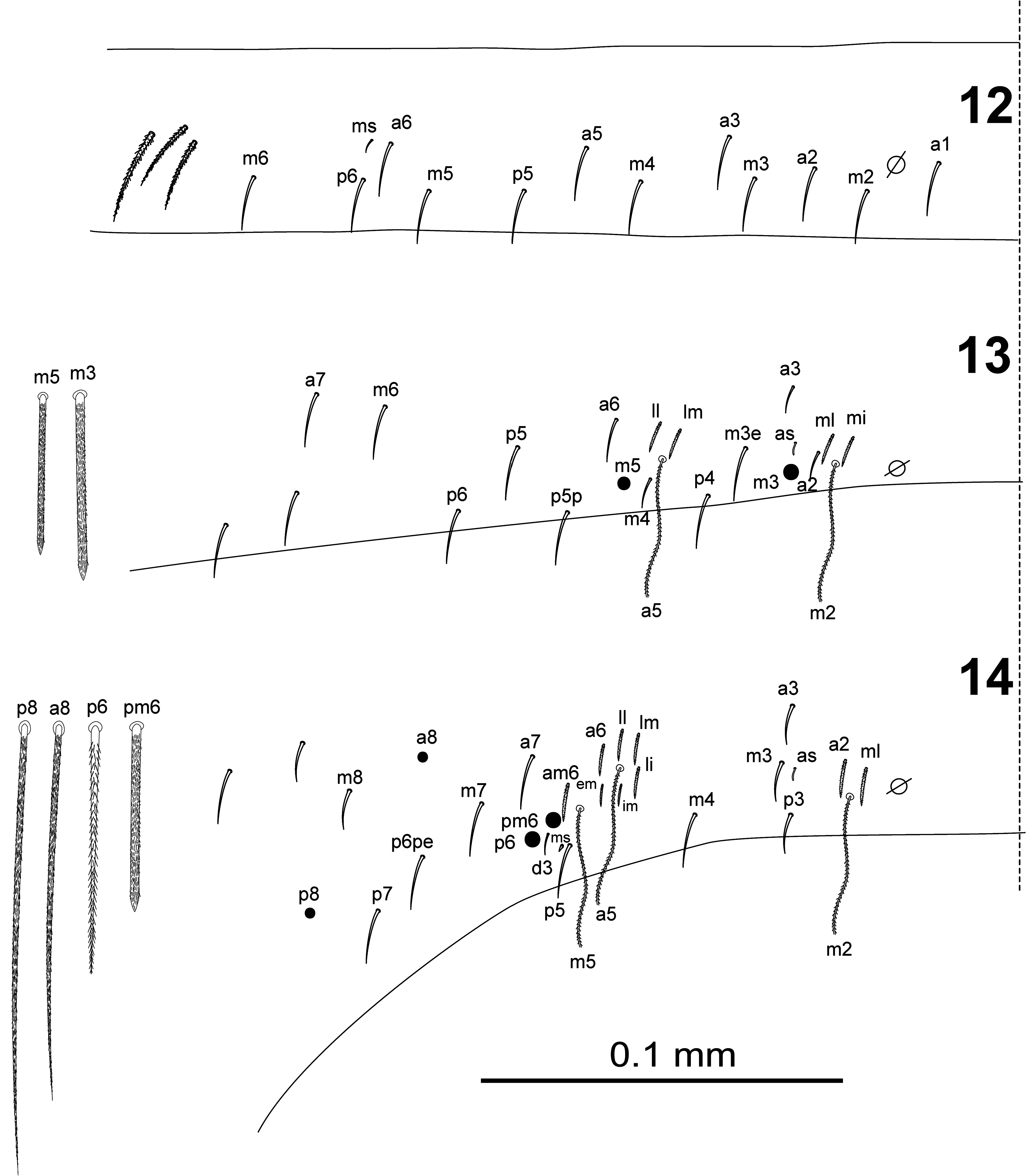
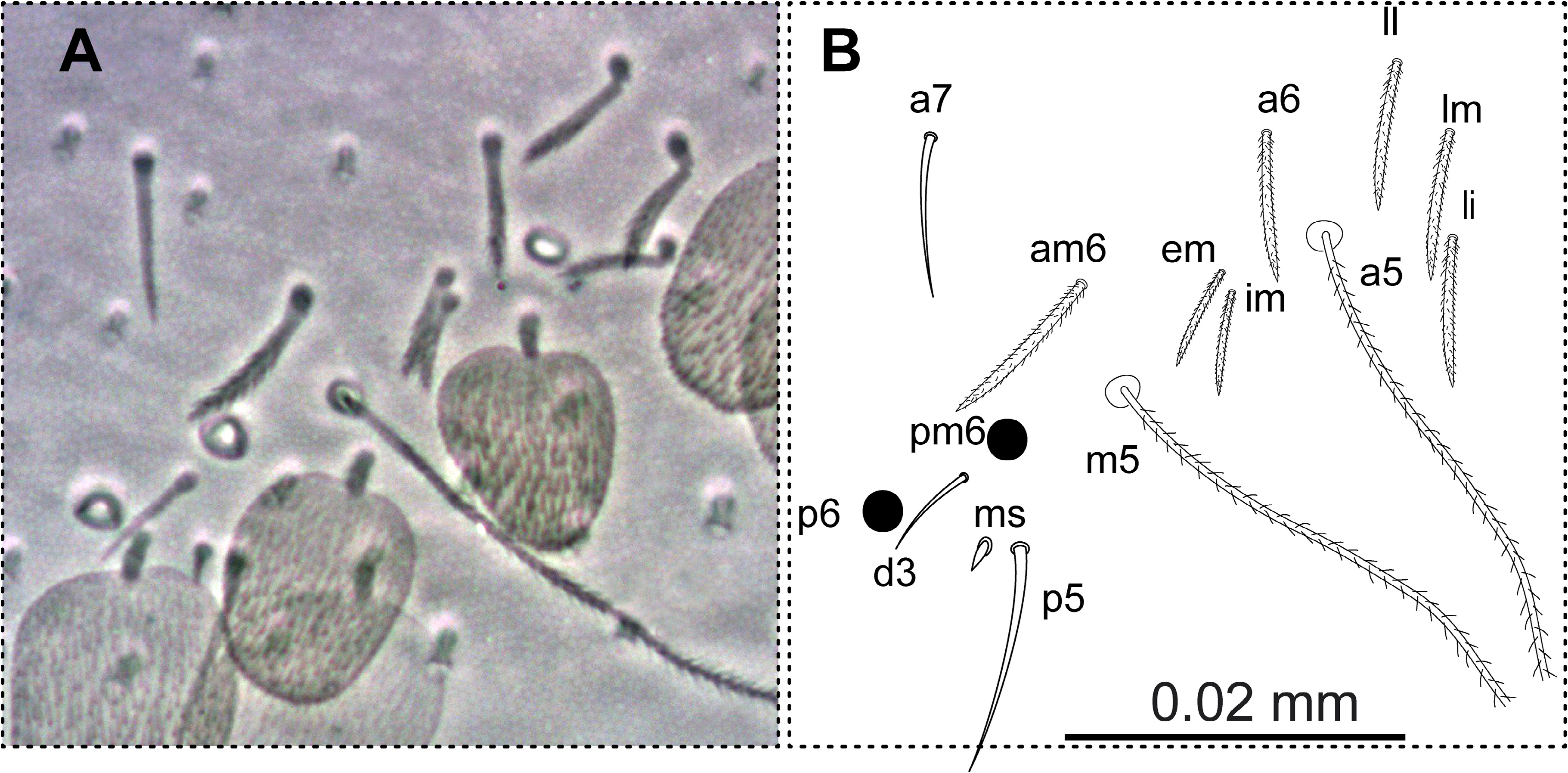
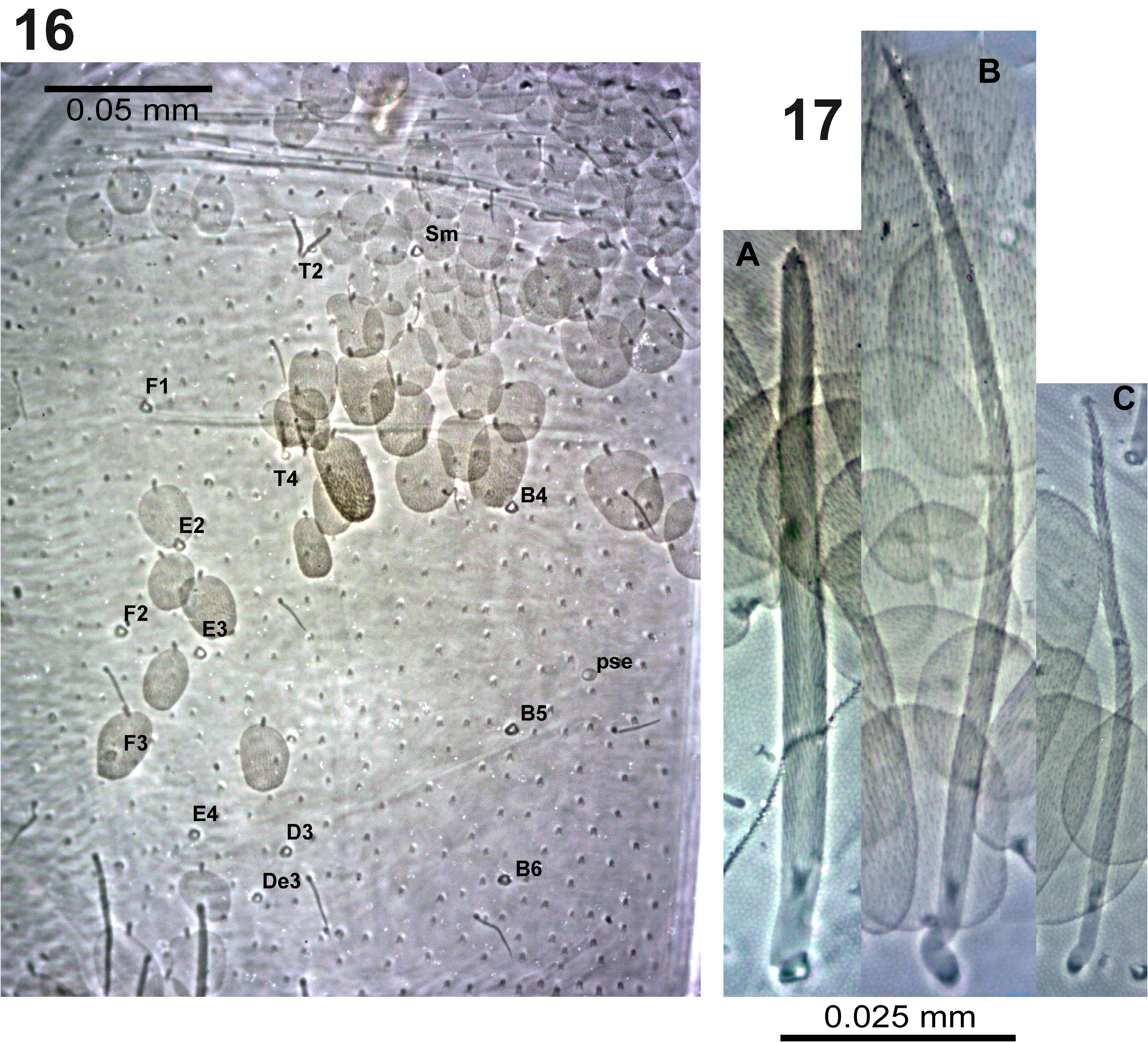
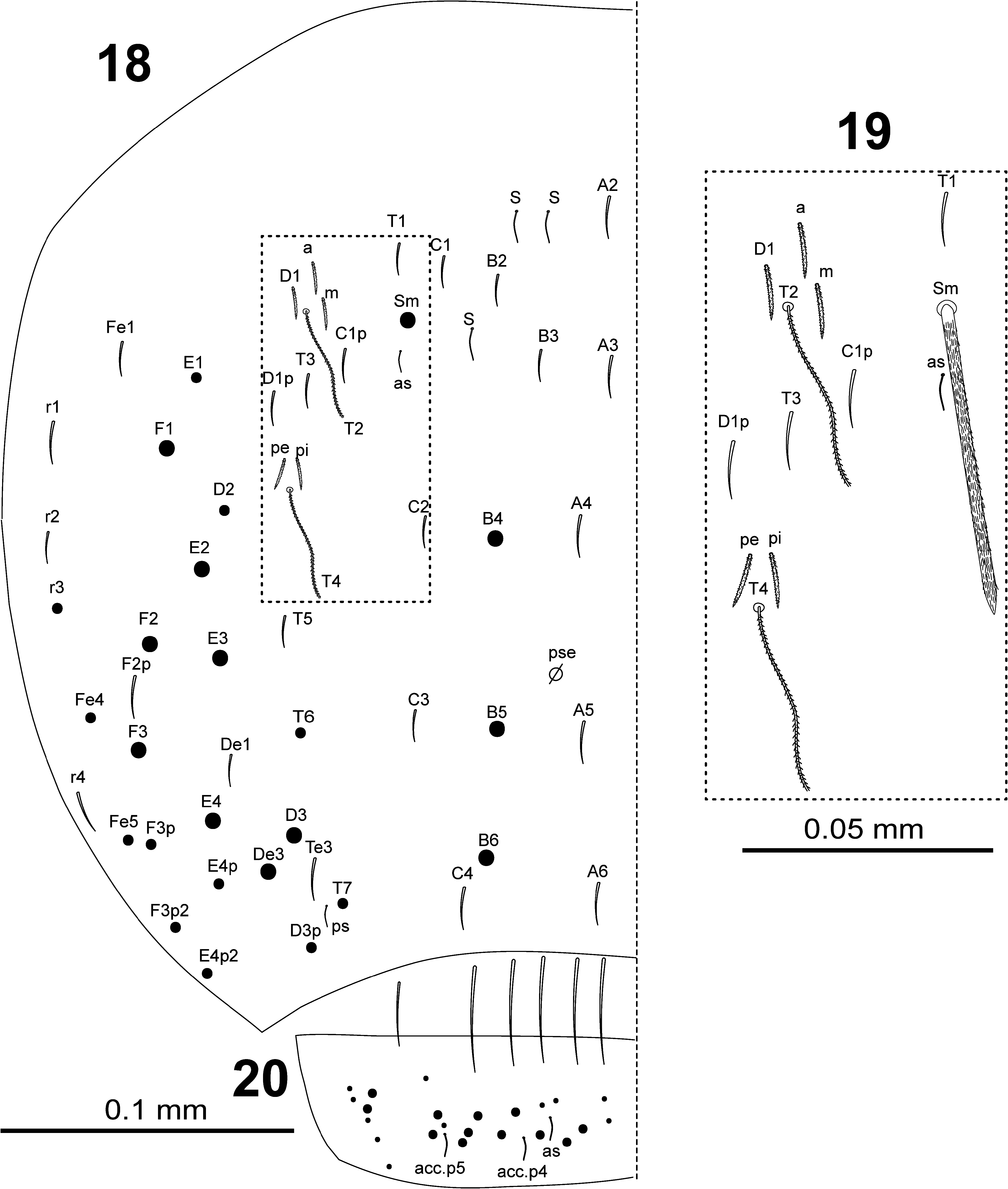
BODY. Dorsal body macrochaetae formula 10/0101+3 (macrochaetae p3 on Th.II, m3 on Abd.II, and Sm+B4, B5, B6 on Abd.IV). Dorsal chaetotaxy of Th.II–III as in Figs 10–11 View FIGURES 10–11 . Th.II with 2 lateral S-chaetae ( al and ms) and with one macrochaetae ( p3) in dorsal position. Th.III with a lateral sensillum ( al) close to several ciliated chaetae. Chaetotaxy of Abd.I–III as in Figs 12–14 View FIGURES 12–14 .Abd.I with a lateral S-microchaeta ( ms) external to a6. Abd.II macrochaeta m3 1.2 times longer than macrochaeta m5. Abd. III chaeta mi absent, with chaeta d3 between macrochaetae pm6 and p6, and with S-chaetae as and ms. All chaetae associated with the trichobothria on Abd.II–III strongly ciliate ( Fig. 15 View FIGURE 15 ). Chaetotaxy of Abd.IV as in Figs 16–19 View FIGURES 16–17 View FIGURES18–20 ; macrochaetae Sm, B4, B5, B6, D3, De3, E2, E3, E4, F1, F2, F3 with large socket ( Fig. 17-A View FIGURES 16–17 ); macrochaetae T6, T7, D2, E1, E4p, E4p2, Fe4, Fe5, F3p, F3p2, r3 longer or shorter but always with socket of minor diameter ( Fig. 17-B,C View FIGURES 16–17 ); macrochaeta F2 inserted above macrochaeta E3; the ratio of distances between macrochaetae Sm–B4 / B4–B6 as 0.7–0.8; ratio of distances between macrochaetae B4–B5 / B5–B6 as 1.0–1.3; accessory chaeta s associated with trichobothrium T2 absent; chaetae a, D1, m, pe and pi associated with trichobothria T2 and T4 strongly cilate ( Fig. 19 View FIGURES18–20 ); sens chaetotaxy composed of three anterior dorsomedial elongate S-chaetae, and short chaetae as and ps.; posterior margin with 6+6 smooth mesochaetae; lateral region and BP4 without pseudopori. Dorsal chaetotaxy of Abd.V ( Fig. 20 View FIGURES18–20 ) with S-chaetae as, acc.p4 and acc.p5.
APPENDAGES. Antennal segments I–III with scales on all faces. Ratio antenna:cephalic diagonal ≈ 1.6 (head diagonal measured from cervical edge to apex of mouth part); ratio Ant.I:II:III:IV as 1:1.9:1.6:2.8. Proximal margin of Ant.I dorsally with three microchaetae arranged in triangle (Ant.I-organ); ventro-distal membranous margin of Ant.I with a short curved S-chaeta. Ant.III organ composed of two subcilindrical and curved sensory rods. Ant.IV without apical bulb.
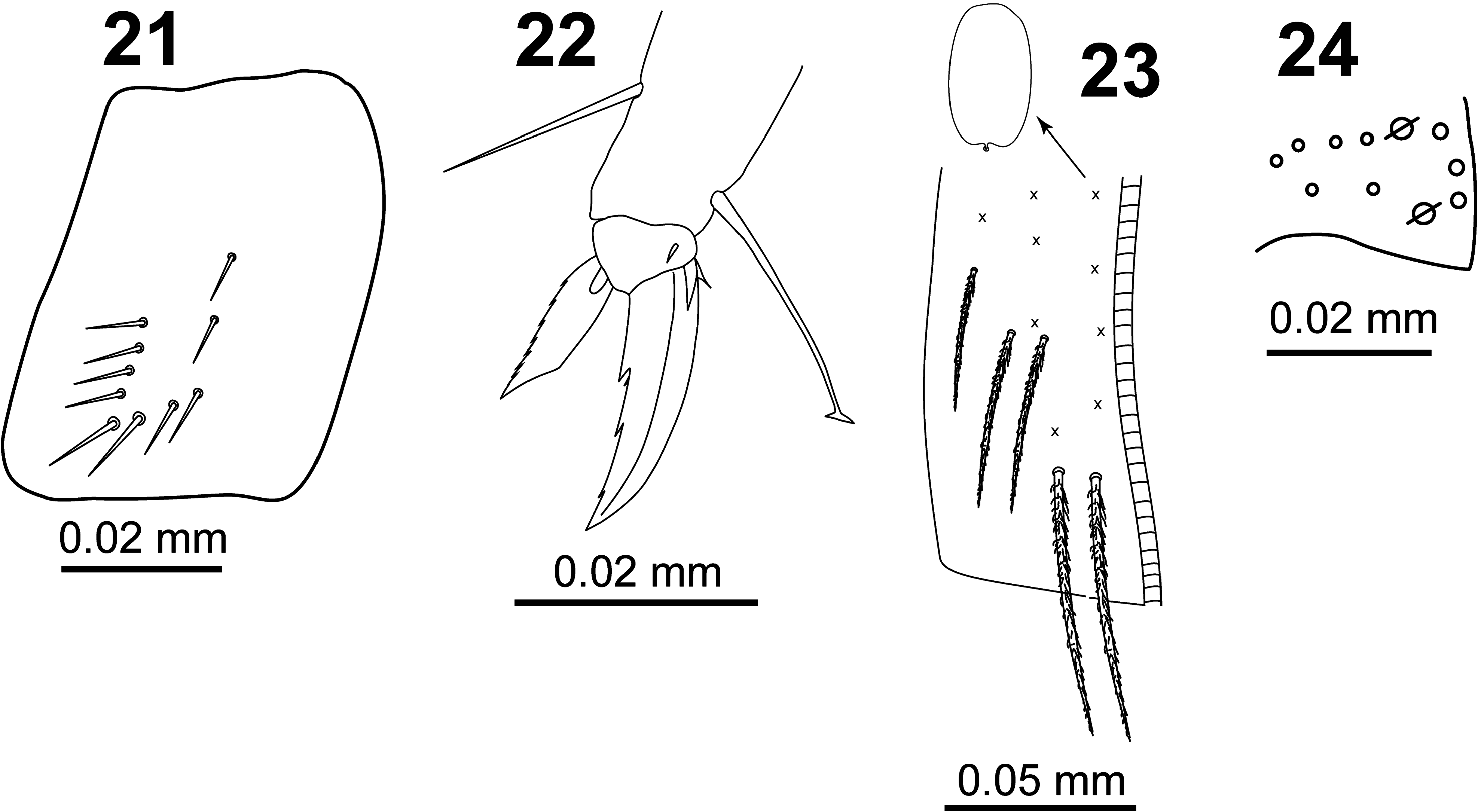
All legs segments with scales. V-shaped trochanteral organ ( Fig. 21 View FIGURES 21–24 ) formed by a maximum of 12 smooth straight chaetae. Unguis ( Fig. 22 View FIGURES 21–24 ) with basal pair of teeth at 47% from base of inner edge and with two inner unpaired teeth at 65% and 84% from base of inner edge, respectively; apical inner tooth minute and sometimes difficult to see; one external tooth and a pair of lateral teeth also present. Unguiculus truncate with finely serrated outer margin. Tenent hair spatulate, smooth and a little longer than claw (ratio TH/claw ≈ 1.1); ratio of supra-empodial chaeta (smooth chaeta on tibiotarsus III opposite to tenent hair) / unguiculus ≈ 1.3.
Ventral tube with 5+5 ciliated chaetae on anterior side ( Fig. 23 View FIGURES 21–24 ) and 8+8 ciliated chaetae on posterior side; scales present on anterior and posterior sides; lateral flaps with a maximum of 17 laterodistal chaetae (7–10 ciliated and 7 smooth).
Manubrium with scales on anterior and posterior surfaces, with 2+2 ciliated apical chaetae on anterior side. Ratio manubrium:dens:mucro as 15:17:1. Manubrial plate ( Fig. 24 View FIGURES 21–24 ) with two pseudopores, three inner chaetae, and a maximum of six outer chaetae. Dental tubercle absent. Mucro with two subequal teeth, basal spine smooth (without spinelet).
PSEUDOPORES. Pseudopores distribution on dorsal and ventral regions of head, body, and appendages as in Figs 25-A, B View FIGURE 25 .
Ecology and distribution. All specimens were collected from grass in a small grassland surrounded by forests. The only known locality is the type locality in Italy.
Discussion. Apart from the presence of one macrochaeta on Th.II, all the other morphological characters of the Lepidocyrtus thoracicus sp. nov., as well as the molecular analyses, indicate that it belongs to the Lepidocyrtus lignorum -group ( sensu Mateos 2011). The presence of a dorsal macrocheata in Th.II is a character not found so far in any species of the L. lignorum -group, and it represents an excellent diagnostic character to differentiate the new species within the group. It should be noted that in the Lepidocyrtus species described so far, when they have a mesothoracic macrochaeta, it is always p3. In the European fauna, this mesothoracic macrochaeta is present in all species of the lusitanicus -group and lanuginosus -group (see Mateos et al. 2021). Out of Europe, the only species in which the presence of the mesothoracic p3 macrochaeta has been described are L. vireticulus Mari Mutt, 1986 and L. diminutus Mari Mutt, 1986 , both from Puerto Rico. Molecular analyzes place the new species as a sister group to L. pulchellus Denis, 1926 . Morphologically, both species are very similar, and the new species differs from L. pulchellus by the presence of the mesothoracic macrochaeta p3, the color pattern, and slightly smaller body size. Molecularly both species are clearly differentiated ( Fig. 2 View FIGURE 2 ). Within the L. lignorum -group, four species have truncated unguiculus, L. peisonis Traser & Christian, 1992 , L. pulchellus , L. ruber , and L. uzeli Rusek, 1985 (see Mateos 2020); the presence of the mesothoracic macrochaeta p3 and the color pattern clearly differentiate the new species from all of them.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |