
Cathorops festae (Boulenger 1898)
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4170.1.7 |
|
publication LSID |
lsid:zoobank.org:pub:1FB49F66-80A4-43E7-AD96-1CDFF90723F9 |
|
DOI |
https://doi.org/10.5281/zenodo.5619840 |
|
persistent identifier |
https://treatment.plazi.org/id/974B87B4-1C3B-FFB5-FF66-FDAC5443CBE4 |
|
treatment provided by |
Plazi |
|
scientific name |
Cathorops festae (Boulenger 1898) |
| status |
|
Cathorops festae (Boulenger 1898)
Figure 1 View FIGURE 1 and 2 View FIGURE 2 , Table 2 and 3 View TABLE 3
Arius festae Boulenger, 1898:5 (original description; Naranjal, western Ecuador).— Kailola & Bussing, 1995:866 (as status uncertain).—Marceniuk & Ferraris in Reis et al. 2003:453 (list, as species inquirenda).—Betancur-R. & Acero P., 2004:13 (as species inquirenda).— Ferraris 2007:56 (as species inquirenda).— Marceniuk & Menezes 2007:46 (as species inquirenda).— Marceniuk et al. 2009:274 (as species inquirenda).
Cathorops fuerthii View in CoL (non Steindachner).— Sanchez, 2008:10, 20, 28 (list; in part).— Barriga, 2012: 113 (list; in part).— Moncayo & Noboa, 2014:2 (list; in part).— Jiménez-Prado et al., 2015: 399 (identification key; in part).
Diagnosis. Cathorops festae is distinguished from the eastern Pacific subcongeners by having a supraoccipital process broader on posterior portion (3.4−4.0% SL vs. 2.2−3.3% SL in all other representatives, excluded C. fuerthii ), longer nuchal plate (7.2−7.9% SL vs. 4.7−7.1% SL in all other representatives, excluded C. liropus , C. multiradiatus , C. taylori and C. tuyra ), and the edges of the distal third of the occipital process which present a marked anterior-posterior convergence (vs. parallel or subtly convergent in all other forms, except C. fuerthii and C. raredonae , Fig. 2 View FIGURE 2 f,g). The species can be further differentiated as follows: from C. fuerthii , which ranges from Costa Rica to Panama, by its shorter distance between anterior nostrils (3.3−4.2% SL vs.4.4−6.0% SL), shorter distance between posterior nostrils (4.1−5.8% SL vs. 5.7−7.3% SL, rarely 5.8%), larger orbital diameter (4.1−5.4% SL vs. 3.2−4.1% SL, rarely 4.2%), longer supraoccipital process (12.0−14.1% SL vs. 10.4−11.9% SL), and longer caudal-fin upper lobe (31.3−32.8% SL vs. 28.3−30.7% SL); from C. hypophthalmus , which is found in Panama, by having 17-18 gill rakers on first brachial arch (vs. 37−40), 16−19 rakers on second brachial arch (vs. 37−40), narrower mouth (9.5−11.1% SL vs. 13.1−13.2% SL), shorter distance between anterior nostrils (3.3−4.2% SL vs. 6.9−7.1% SL), shorter distance between posterior nostrils (4.1−5.8% SL vs. 8.2−8.5% SL), deeper caudal-peduncle (7.8−9.0% SL vs. 7.3−7.5% SL), and caudal-fin lobes wide, rounded posteriorly (vs. narrow and pointed posteriorly); from C. liropus , which is found in Mexico, by caudal-fin lobes wide, rounded posteriorly (vs. narrow and pointed posteriorly); from C. manglarensis , which ranges from Colombia to Peru ( Fig. 3 View FIGURE 3 ), by having 17−18 gill rakers on first brachial arch (vs. 13−16, Table 2), wider nuchal-plate (7.2−8.1 vs. 6.4−7.1, Fig. 4 View FIGURE 4 a), and shorter anal-fin base (15.3−19.3% SL vs. 21.0−24.4% SL, Fig. 4 View FIGURE 4 b); from C. multiradiatus , which ranges from Panama to Peru ( Fig. 3 View FIGURE 3 ), by its longer distance from snout to dorsal fin (36.1−40.6% SL vs. 31.2−34.8% SL, Fig. 4 View FIGURE 4 c), longer supraoccipital process (12.0−14.1% SL vs. 9.8−11.5% SL, Fig. 4 View FIGURE 4 d), and shorter anal-fin base (15.3−19.3% SL vs. 20.0−23.2% SL, Fig. 4 View FIGURE 4 b); from C. raredonae , which ranges from Mexico to El Salvador, by its longer supraoccipital process (12.0−14.1% SL vs. 8.7−10.5% SL), and wider nuchal-plate (7.2−8.1% SL vs. 5.9−7.1% SL); from C. steindachneri , which ranges from El Salvador to Panama, by its longer caudal-fin upper lobe (31.3−32.8% SL vs. 25.1% SL), longer caudal-fin lower lobe (29.7−32.8% SL vs. 23.8% SL), and caudal-fin lobes wide, rounded posteriorly (vs. narrow and pointed posteriorly); from C. taylori , which ranges from Guatemala to El Salvador, by its longer supraoccipital process (12.0−14.1% SL vs. 9.4−10.6% SL), and caudal-fin lobes wide, rounded posteriorly (vs. narrow and pointed posteriorly); from C. tuyra , which is found in Panama, by having 23−25 anal-fin rays (vs. 19−20); 17−18 gill rakers on first arch (vs. 19−22); and deeper caudal-peduncle (7.8−9.0% SL vs. 5.8−7.8% SL). Additionally, Cathorops festae is differentiated from the subgenus Precathorops [currently monotypic with C. (P.) dasycephalus ] found in the eastern Pacific by having 17−18 gill rakers on first brachial arch (vs. less than 10), vomerine tooth plates absent (vs. present in Precathorops ), and accessory tooth plates bearing molariform teeth (vs. conical in Precathorops ).
Gill rakers on the first gill arch 13 14 15 16 17 18 19 C. festae 1 3 C. manglarensis 1 2 4 2
C. multiradiatus 1 1 2 3 Cathorops festae is distinguished from the Western Atlantic subcongeners by having a supraoccipital process broader on posterior portion (3.4−4.0% SL vs. 2.0−3.2% SL in all other representatives, excluded C. arenatus and C. nuchalis ), and 17−18 gill rakers on first arch (vs. 14−16 or 19−24 in all other representatives, excluded C. arenatus , C. melanopus , C. spixii , and C. wayuu ). The species can be further differentiated from the Western Atlantic subcongeners as follows: from C. aguadulce , which is found in Mexico, by having 23−25 anal-fin rays (vs. 21); from C. arenatus , from mouth of the Orinoco River to Brazilian north coast, by having osseous medial groove long and progressively larger to posterior-anterior direction (vs. short and narrow at middle portion), shorter distance between anterior nostrils (3.3−4.2% SL vs. 4.1−5.8% SL, rarely 4.2%); from C. belizensis , from Belize, by having 23−25 anal-fin rays (vs.18−20), shorter distance between anterior nostrils (3.3−4.2% SL vs. 5.1−6.1% SL); from C. kailolae , which ranges from Mexico to Guatemala, by its wider cephalic shield ate supracleithrum area (17.7−19.8% SL vs. 15.3−17.3% SL), and longer nuchal-plate (7.2−7.9% SL vs. 5.3−6.9% SL); from C. mapale , which is found in Colombia, by its longer nuchal-plate (7.2−7.9% SL vs.6.8−7.1% SL, rarely 7.2%); from C. melanopus , which is found in Guatemala, by having 23−25 anal-fin rays (vs. 19−20); from C. nuchalis , which ranges from Venezuela to Guyana, by its shorter distance between anterior nostrils (3.3−4.2% SL vs. 4.3−5.5% SL, rarely 4.2% SL); from C. wayuu , which ranges from Colombia to Venezuela, by its shorter distance between anterior nostrils (3.3−4.2% SL vs. 4.1−5.2% SL, rarely 4.2%), and longer supraoccipital process (12.0−14.1% SL vs. 9.1−11.5% SL).
Description. Morphometric data in Table 3 View TABLE 3 . Head depressed, profile elevated posteriorly, straight at frontal and supraoccipital area. Snout short and transversely rounded. Anterior nostril round, with fleshy edge, posterior nostril covered by flap of skin; nostrils relatively close to one another and moderately close to orbit, not connected by fleshy furrow. Eye lateral and large; eyes distant to one another. Three pairs of moderately long teretiform barbels; maxillary barbel usually reaching or past base of pectoral-fin spine, lateral mental barbel reaching gill membrane edge, and mesial mental barbel not reaching gill membrane edge. Osseous bridge formed by lateral ethmoid and frontal long and slender, evident under skin. Cephalic shield exposed, rough and granulated; moderately long and large on lateral ethmoid, frontal, and supracleithrum areas. Anterior portion of dorsomedial groove of neurocranium fleshy, conspicuous and not continuous to level of posterior nares; posterior portion of osseous groove, deep and conspicuous, with straight margins, narrowing posteriorly. Supraoccipital process funnel-shaped, very long and wide on posterior portion, and profile straight. Nuchal plate crescent-shaped, very long and wide.
Mouth subterminal to terminal, relatively large; lower jaw arched. Lips moderately thick, lower lip as thick as upper lip. Vomerine tooth plates absent. One pair of oval shaped accessory tooth plates, variable in size, closer to one another anteriorly, with molariform teeth. Premaxilla relatively long and narrow, with sharp teeth. Dentary separated at midline, with posterior expansion and sharp teeth on anterior portion, molariform teeth on posterior portion and some conical teeth interspersed. Gill membranes fused, attached to isthmus. Seventeen to 18 acicular gill rakers on first arch, 16–19 spike-shaped gill rakers on second arch. Mesial surfaces of all gill arches with developed gill rakers, lateral and mesial surfaces of first and second gill arches lacking fleshy papillae intercalated with gill rakers.
Body width greater than depth in pectoral girdle area, progressively more compressed from pectoral fin to caudal peduncle. Lateral line sloping ventrally on anterior one-third, extending posteriorly to caudal peduncle, bending abruptly onto dorsal lobe of caudal fin. Dorsal-fin spine moderately long and thick; anterior margin with granules; posterior margin with serrations along almost its entire length. Seven soft dorsal-fin rays. Pectoral-fin spine moderately long, shorter than dorsal-fin spine; anterior margin with granules on basal two-thirds and distal one-third with short serrations; posterior margin straight on basal one-fourth, distal three-fourths with serrations. Ten soft pectoral-fin rays. Posterior cleithral process exposed, smooth and triangular shaped, short and pointed posteriorly. Pelvic fin deep and long at base, with six rays. Adipose-fin base short, less than one-half the length of anal-fin base, anterior origin at level of anterior half of anal fin. Anal fin deep and moderately long at base, with 23–25 rays and distal margin slightly concave. Caudal peduncle relatively high. Caudal fin forked, dorsal and ventral lobes moderately long, wide and rounded posteriorly; dorsal lobe longer than ventral lobe.
Coloration. Grayish to dark brown on dorsum, silvery on flanks and white on abdomen. In alcohol, dark brown on dorsum, flanks brownish to silvery and white on venter; fins brownish. Maxillary barbel dark, mental barbel lighter ( Fig. 1 View FIGURE 1 a,b, 2a,b).
Sexual dimorphism. Sexual dimorphism was not observed in the specimens examined (159–179 mm SL).
Distribution and habitat. Cathorops festae has been described from freshwater environments in Ecuador. The new specimens collected (LBP 19381) indicate that the species also occurs in the estuaries of northern Peru ( Fig. 3 View FIGURE 3 ).
Size. The largest examined specimen had a standard length of 179 mm (LBP 19381).
Molecular evidence and phylogenetic relationships. Based on the relationships among the 16 Cathorops species inferred from the mitochondrial sequences using the maximum likelihood approach in RAxML and Bayesian Inference, C. festae from Ecuador and Peru are conspecific and together with C. multiradiatus , found from Pacific Panama to Ecuador, represent the most basal group in the subgenus Cathorops ( Fig 5 View FIGURE 5. A ).
Remarks. Arius festae Boulenger, 1898 was originally described from Naranjal, western Ecuador, remaining for a long time known only from the holotype (MZUT 1479), with its taxonomic status being considered uncertain ( Kailola & Bussing, 1995), or as a species inquirenda ( Marceniuk & Ferraris, 2003; Ferraris 2007; Marceniuk & Menezes 2007, Marceniuk et al. 2009). Its status has been challenged on the basis of third-party examination and photographs of the holotype, as well as molecular data obtained from specimens collected near the type locality ( Marceniuk et al., 2009, Betancur-R., et al., 2012). In the present study, Cathorops festae is recognized as a valid species, distinct from the other members of the genus found in Ecuador and Peru, based on the analysis of the holotype and other, non-type specimens, in which the profile of the distal third of the occipital process is characterized by the marked anterior-posterior convergence of the edges ( Fig. 2 View FIGURE 2 a,b,f), the wider nuchal plate, the shorter base of the anal fin, the greater distance between the snout and the dorsal fin, and the greater length of the occipital process ( Fig. 4 View FIGURE 4 ). Differences in morphometric ratios between the holotype from Ecuador (83 mm SL) and non-type specimens from Peru (159-179 mm SL) are probably the result of ontogenetic variation, which is common in other species of Cathorops .
Material examined. Type-specimen: holotype, MZUT 1479 View Materials , Naranjal , Ecuador . Non-type specimens: LBP 19381 (4, 159−179), Tumbes, Peru, in November 2014.


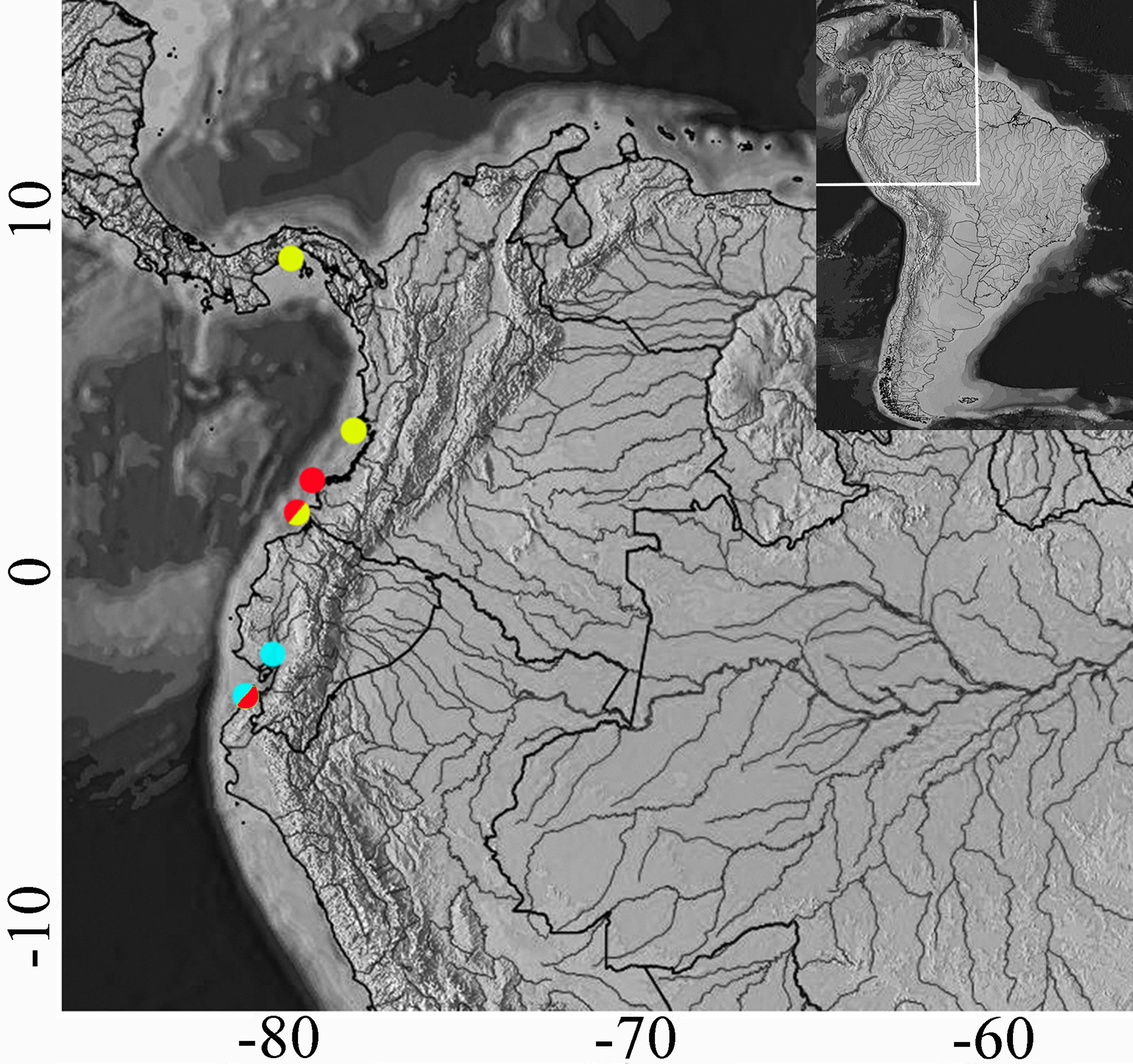


TABLE 3. Morphometrics for Cathorops festae. Standard length is expressed in millimeters, other measurements are percents of standard length. Range include non-type specimens only.
| Cathorops festae | holotype | N | Mean | Range |
|---|---|---|---|---|
| Standard length (mm) | 83 | 4 | 159−179 | |
| Head length | 25.3 | 4 | 27.5 | 26.5−28.2 |
| Snout length | 8.4 | 4 | 7.7 | 7.0−8.6 |
| Distance between anterior nostrils | 3.3 | 4 | 4.0 | 3.9−4.2 |
| Distance between posterior nostrils | 5.2 | 4 | 5.1 | 4.1−5.8 |
| Distance between anterior nostril and orbit | 4 | 7.0 | 6.2−7.7 | |
| Distance between posterior nostril and orbit | 4 | 5.1 | 4.4−5.8 | |
| Orbital diameter | 5.4 | 4 | 4.3 | 4.1−4.4 |
| Interorbital distance | 12.3 | 4 | 14.4 | 13.4−15.0 |
| Maxillary barbel length | 30.4 | 4 | 28.1 | 26.5−31.0 |
| Lateral mental barbel length | 26.7 | 4 | 22.0 | 18.8−24.5 |
| Mesial mental barbel length | 19.4 | 2 | 13.7 | 12.4−14.9 |
| Mouth width | 9.5 | 4 | 10.4 | 9.7−11.1 |
| Width of cephalic shield at lateral ethmoid area | 13.8 | 4 | 12.6 | 12.0−13.3 |
| Width of cephalic shield at frontals area | 7.8 | 4 | 6.3 | 6.2−6.5 |
| Width of cephalic shield at epioccipital area | 4 | 12.7 | 12.4−13.2 | |
| Width of cephalic shield at supracleithrum area | 17.7 | 4 | 19.5 | 19.1−19.8 |
| Supraoccipital process length | 12.0 | 4 | 13.2 | 12.7−14.1 |
| Supraoccipital process width | 4 | 3.7 | 3.4−4.0 | |
| Nuchal-plate length | 7.2 | 4 | 7.5 | 7.3−7.9 |
| Nuchal-plate width | 7.2 | 4 | 7.8 | 7.6−8.1 |
| Body depth | 16.2 | 4 | 19.0 | 18.1−19.9 |
| Body width | 19.0 | 4 | 22.6 | 21.7−23.2 |
| Distance from snout to pectoral fin | 21.7 | 4 | 24.3 | 23.1−26.5 |
| Distance from snout to dorsal fin | 36.1 | 4 | 39.3 | 37.9−40.6 |
| Distance from snout to pelvic fin | 48.2 | 4 | 53.1 | 51.6−54.1 |
| Distance from snout to adipose fin | 70.0 | 4 | 79.5 | 77.8−80.8 |
| Distance from snout to anal fin | 66.3 | 4 | 71.9 | 71.3−72.4 |
| Caudal-peduncle height | 7.8 | 3 | 8.5 | 8.2−9.0 |
| Pectoral-fin spine length | 18.3 | |||
| Dorsal-fin spine length | 20.6 | |||
| Pelvic-fin base length | 4.2 | 4 | 4.3 | 3.7−4.9 |
| Pelvic-fin height | 13.1 | 4 | 15.5 | 13.9−16.4 |
| Adipose-fin base length | 4 | 7.1 | 6.4−7.6 | |
| Adipose-fin height | 4 | 3.9 | 3.3−4.6 | |
| Anal-fin base length | 19.3 | 4 | 15.9 | 15.3−16.5 |
| Anal-fin height | 17.2 | 4 | 12.9 | 12.1−13.7 |
| Caudal-fin upper lobe length | 4 | 31.9 | 31.3−32.8 | |
| Caudal-fin lower lobe length | 4 | 30.6 | 29.7−32.8 |
| MZUT |
Museo di Zoologia, Instituto di Zoologia e Anatomia Comparata Universita di Torino |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Cathorops festae (Boulenger 1898)
| Marceniuk, Alexandre P., Marchena, José & Betancur-R, Ricardo 2016 |
Cathorops fuerthii
| Jimenez-Prado 2015: 399 |
| Moncayo 2014: 2 |
| Barriga 2012: 113 |
| Sanchez 2008: 10 |