
Corethrella (Corethrella) tigrina Borkent & Grafe
|
publication ID |
https://doi.org/ 10.5281/zenodo.210864 |
|
DOI |
https://doi.org/10.5281/zenodo.5619818 |
|
persistent identifier |
https://treatment.plazi.org/id/9720782F-AA0A-FFA2-FF78-FDB146A30978 |
|
treatment provided by |
Plazi |
|
scientific name |
Corethrella (Corethrella) tigrina Borkent & Grafe |
| status |
sp. nov. |
Corethrella (Corethrella) tigrina Borkent & Grafe , new species
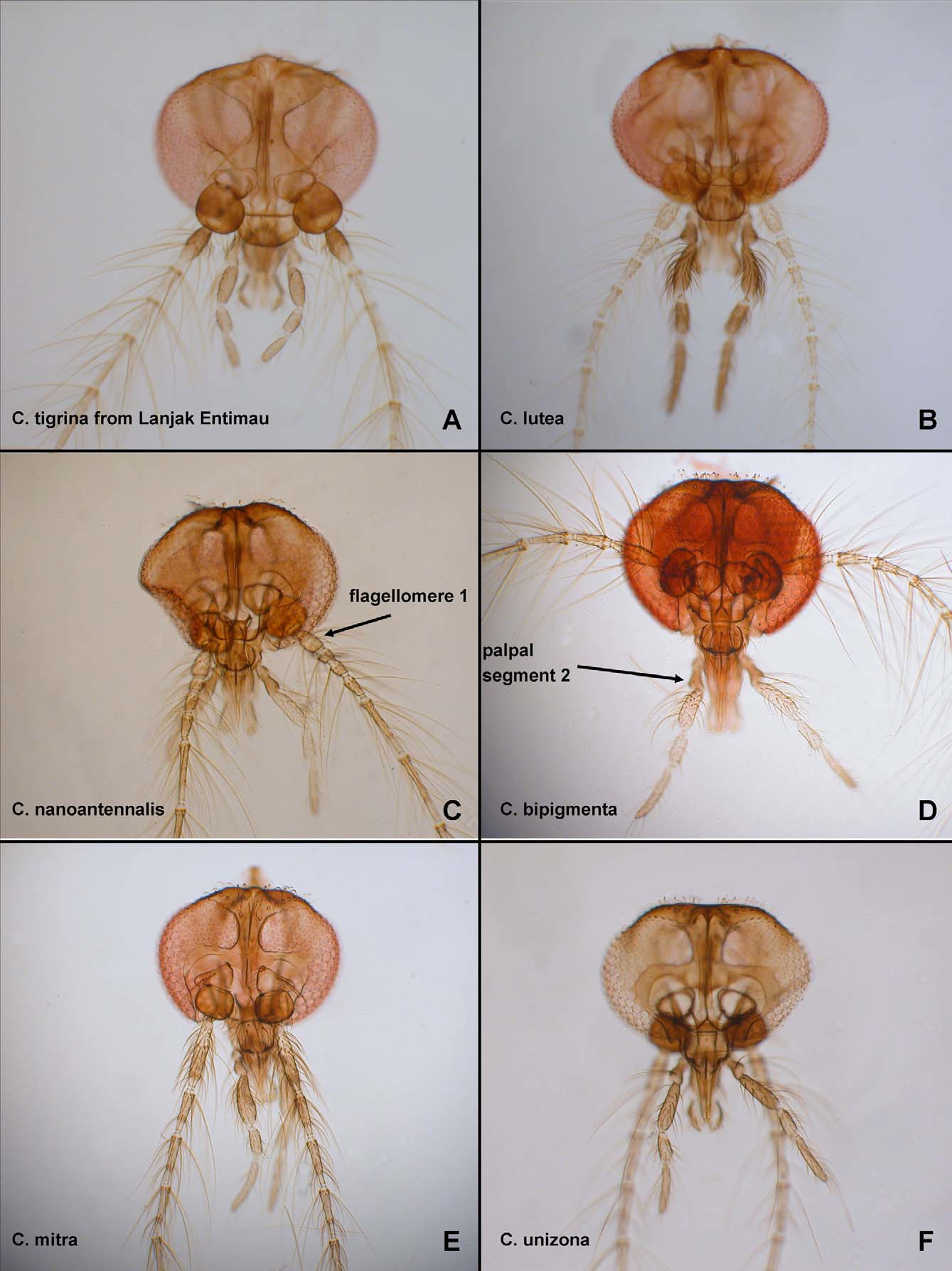
( Figs. 2 View FIGURE 2 A, 4A, 6A, 7E, G, 8A, L, M, 9A, 12A, 13A, 14B)
DIAGNOSIS: Male and female adults: Only extant species of Corethrella in the Palaearctic, Oriental, Oceanian and northern Australasian Regions with the combination of a wing with a subapical band ( Fig. 12A View FIGURE 12. A – K ) and with abdominal tergites 2–6 each with anterior dark pigmentation and lighter posteriorly ( Fig. 13A View FIGURE 13. A – J ). Only extant species in Borneo with the combination of a wing with a subapical band ( Fig. 12A View FIGURE 12. A – K ) and all femora and tibiae each with darker pigmentation at both base and apex ( Fig. 9A View FIGURE 9. A – D ).
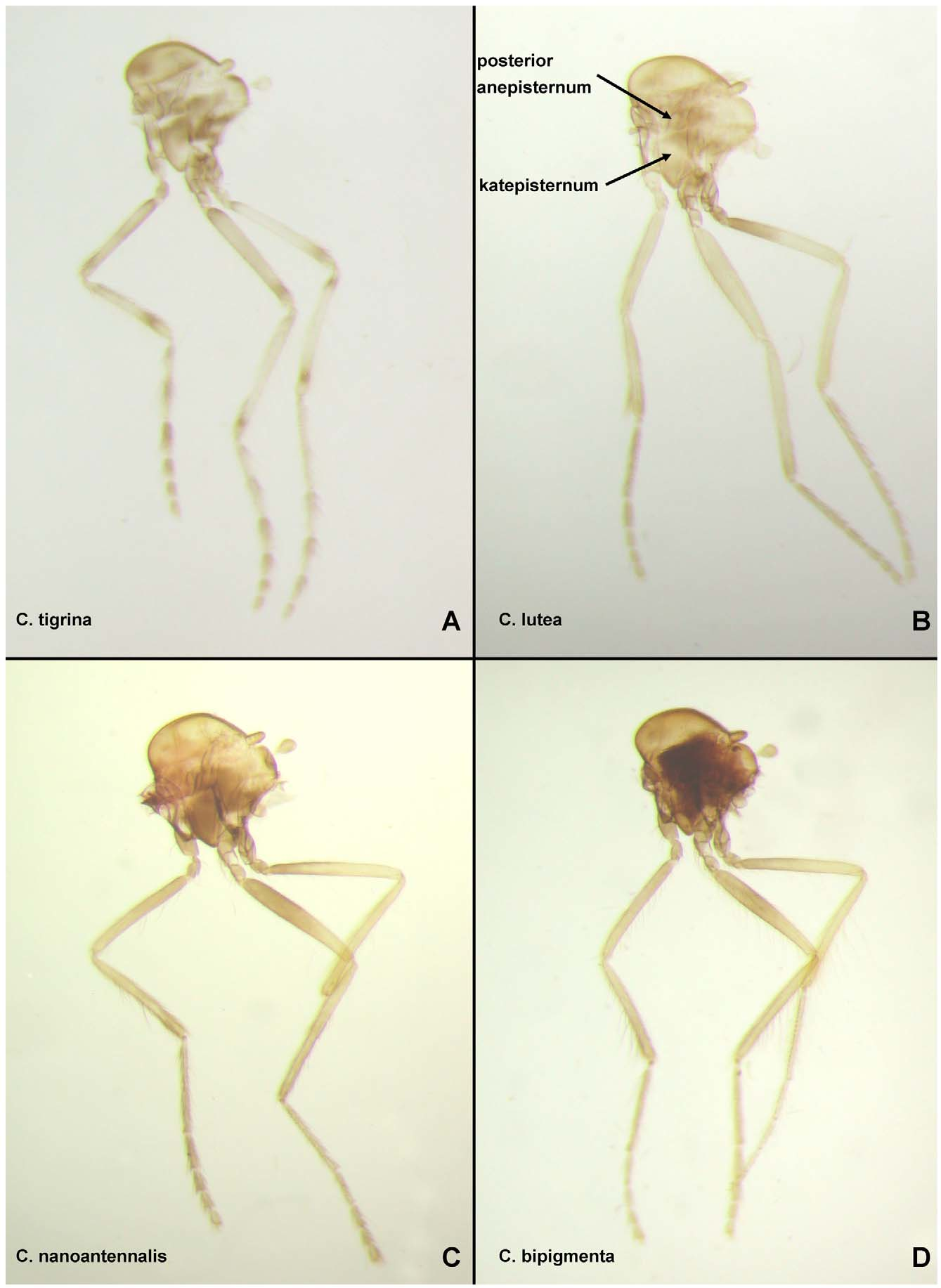
DESCRIPTION: Male adult. Head: Outline in anterior view somewhat circular (as in Fig. 6 View FIGURE 6 A). Two large setae on frons between ventromedial area of ommatidia. Antenna with flagellomere 1 dark brown, remainder pale; pedicel without distinctive elongate, stout setae; flagellomeres as in Fig. 8 View FIGURE 8 L; sensilla coeloconica on flagellomeres 1, 9–13, only flagellomere 1 with more than one; flagellomere 13 with well-developed apical bifurcation. Palpus brown; segment 3 of nearly constant width. Thorax (as in Fig. 9A View FIGURE 9. A – D ): Light brown with following medium to dark brown: paratergite, margin of scutum posterior to paratergite, dorsal portion of mediotergite, posterodorsal portion of anterior anepisternum, posterior anepisternum, dorsal and ventral portion of anepimeron (pale at midheight), most of katepisternum (pale area at about midheight on posterior margin), dorsal portion of metanepisternum. Posterior portion of dorsocentral row with 2 elongate setae situated somewhat lateral to one another. Prescutal suture elongate, thick, uninterrupted, extending to near dorsocentral row of setae. Anterior anepisternum divided diagonally by sinuous suture, dorsal portion about equal to ventral portion. Ventral portion of posterior anepisternum triangular, uniformly brown, with anterodorsal margin not thick. Wing (as in Fig. 12A View FIGURE 12. A – K but slightly lighter): Length = 0.95 mm. Apex of R2 slightly distal to apex of M1. Anterior margin with differently, discretely pigmented scales (indicating anterior margin of midlength, subapical bands), with midlength, subapical bands, with dark scales on R4+5 and M connecting the two bands; veins (other than costa and wing margin) with well-developed scales. Halter pale. Legs (as in Fig. 9A View FIGURE 9. A – D ): Light brown with forefemur with basal and apical darker pigmentation, each of following with discrete basal and apical dark pigmentation: foretibia, midfemur, midtibia, hind femur, hind tibia, fore-, mid-, hind leg tarsomeres 1 each with two bands, fore-, mid-, hind leg tarsomeres 2–4 with basal banding. With only slender setae, lacking scales. Midleg with thick, subapical setae on each of at least tarsomeres 1–3. Apices of fore-, midleg fifth tarsomeres undivided, with claws slightly subapical. Foreleg with tarsomeres 3/ 4 = 1.42. Claws of fore-, midleg equal in length, longer than those of hind leg. Each claw without inner tooth. Anterior claws of each leg without basal prong. Foreleg claws unequal. Midleg claws unequal. Foreleg tarsomere 3 longer than tarsomere 4. Empodia slender. Abdomen: Pale except as follows: tergites 1–7, sternites 3–6 each with anterior band of brown pigmentation, segments 8–9 light brown. Genitalia ( Fig. 8 View FIGURE 8 M): Gonocoxite uniformly pale, gently tapering; anteromedial area with spicules similar in length to those elsewhere on gonocoxite; with well-defined dorsal row of setae, with anterior setae of similar thickness as some other setae on gonocoxite, more posterior setae thicker, with row curving posteromedially with posteriormost seta of row positioned dorsomedially. With one dorsomedial stout seta, tapering near apex. Gonostylus (in retracted position) with slight concavity along posterior margin, curved subapically, thick, of more or less equal thickness for entire length, tapered apically; one elongate, thick subbasal seta, situated ventrally; with thick, blunt apical peg. Aedeagus slender, elongate, tapering gradually to apex, pointed apically, with lateral margins separate apically. Dorsomedial seta from base of gonocoxite/ length of gonocoxite = 0.39. Subbasal seta from base of gonostylus/ length of gonostylus = 0.37.
Female adult. Descriptive statistics: See Tables 1 View TABLE 1 –5. As for male, with following differences. Head: Antenna light brown, with flagellomere 1, apical portion of flagellomeres 4–10, all of flagellomeres 11–13 medium brown. Antennal flagellomeres as in Fig. 8 View FIGURE 8 A; sensilla coeloconica on flagellomeres 1, 9–13, only flagellomere 1 with more than one. Coronal suture elongate, extending ventrally past ommatidia. Clypeus ( Fig. 6 View FIGURE 6 A) somewhat wider than long, triangular in outline. Mandible with very small, pointed teeth. Palpus as in Fig. 6 View FIGURE 6 A. Wing ( Fig. 12A View FIGURE 12. A – K ): As for male but some with some darker pigmentation in subbasal area. Halter medium brown, somewhat paler or as dark as scutellum. Legs: Claws of each leg equal to those of others; equal on each leg, simple (without inner teeth). Abdomen ( Fig. 13A View FIGURE 13. A – J ): Pale or very light brown with anterior margin of each of tergites 1–7 and anterior portion of sternites 3–6 medium brown, segments 8–9 medium brown. Cercus medium brown.
Immature stages. Unknown.
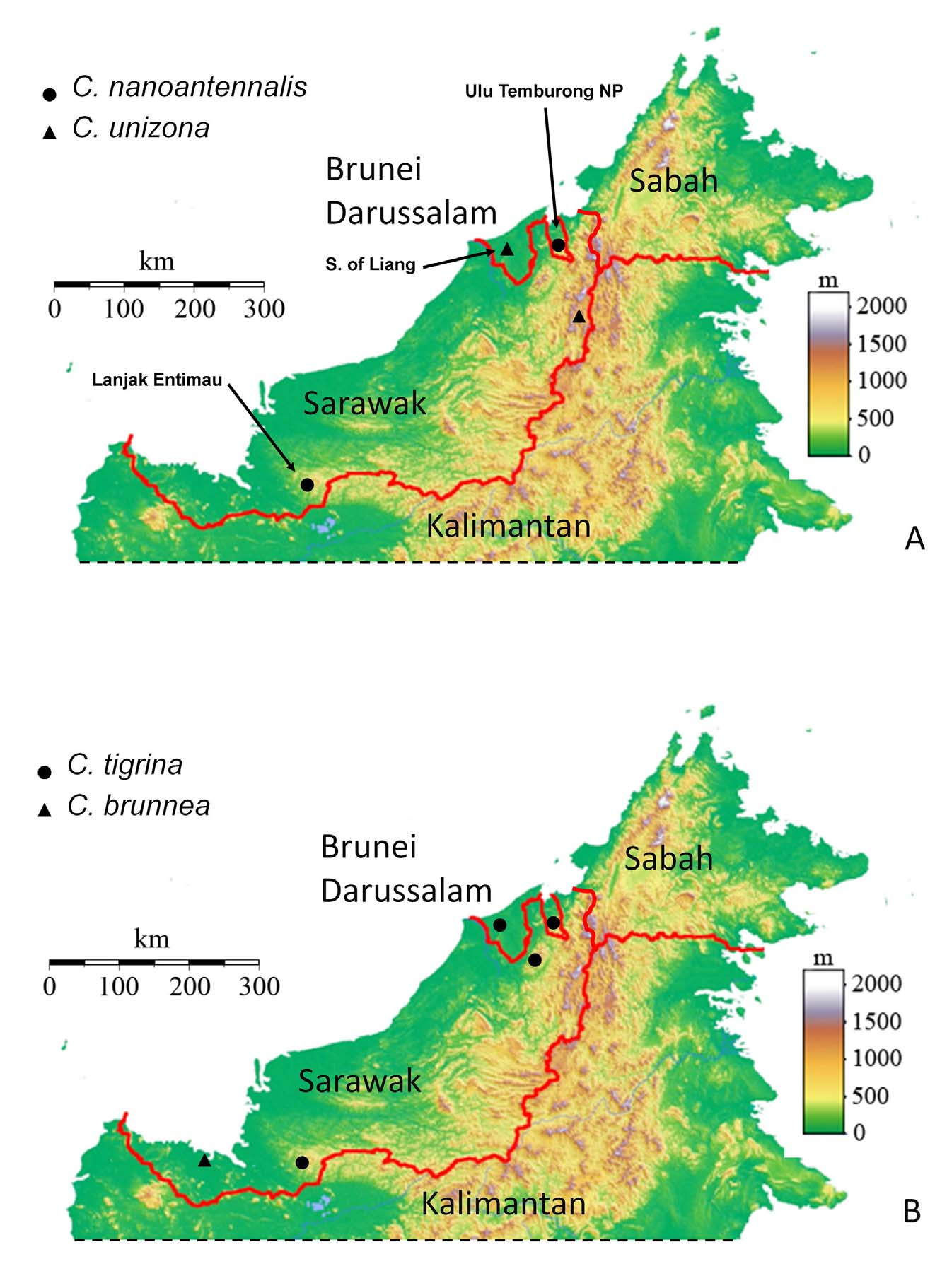
DISTRIBUTION AND BIONOMICS: Corethrella tigrina is known from Brunei and two localities in Sarawak, Malaysia at elevations of 30–220 m ( Fig. 14B View FIGURE 14. A – B ). Habitats include lowland peat swamps (including standing water and very slow moving streams) and steep terrain in mature mixed dipterocarp forest ( Figs. 1 View FIGURE 1 A, F). The sexes were associated on the basis of a male and two females collected with a frog-call trap at 15 km S. of Liang, Brunei on 7-VI-2007. This is the only instance we know of in which a male Corethrella has been collected with a frog-call trap and provides very limited additional evidence supporting the tentative suggestion by Borkent (2008) that males may be attracted to the vicinity of frog calls to mate.
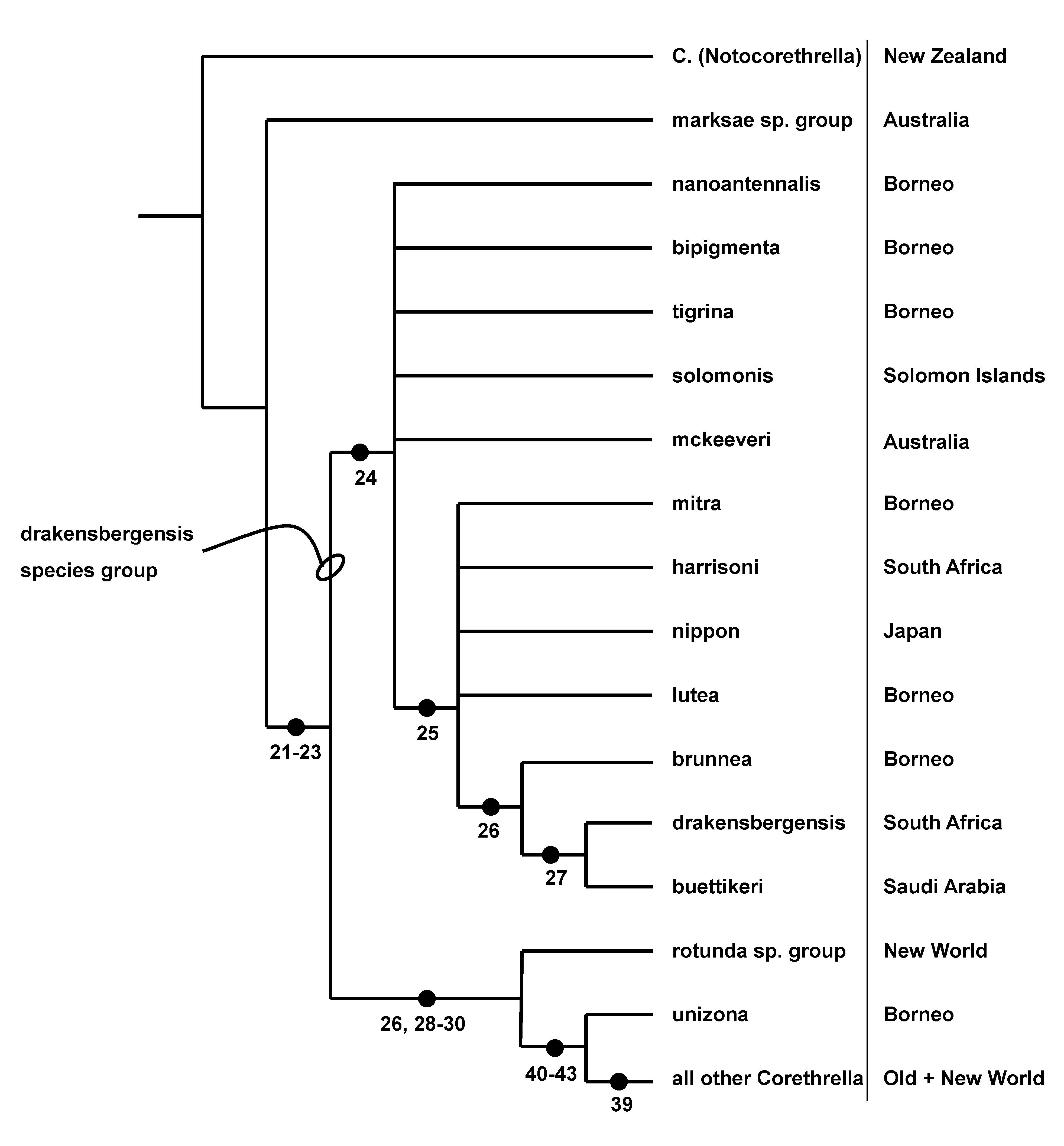
This species is a member of the drakensbergensis species group ( Fig. 19 View FIGURE 19 ) and morphologically very similar to C. solomonis Belkin. The latter species has been collected as larvae from the leaf axil of a Pandanaceae (possibly Sararanga ) ( Borkent, 2008), suggesting that C. tigrina immatures may occur in a similar habitat. However, as indicated in our conclusions, intense sampling from phytotelmata in Borneo has failed to find any immature Corethrella (other than C. calathicola in species of Nepenthes ).
Adult female C. tigrina have been collected with frog-call traps and from calling males of two species of frogs (Table 7).
TAXONOMIC DISCUSSION: There is puzzling variation in the size of the female adult flagellomeres 1–3, a feature which otherwise provides consistent evidence of differences between species of Corethrella . In specimens from Brunei each of these flagellomeres is short ( Fig. 7 View FIGURE 7. A – D E) and for most of those from Lanjak Entimau, Sarawak, each is relatively elongate ( Fig. 7 View FIGURE 7. A – D G). However, one specimen from Lanjak Entimau has flagellomeres that are somewhat intermediate in size; otherwise it is indistinguishable from three others collected at the same place and time with more elongate flagellomeres. We could find no other consistent differences between any of these specimens that would support considering them as separate species. Lim (2007) sequenced specimens from both Brunei and Lanjak Entimau and considered them the same, confirming the conclusion here that the differences in flagellomere lengths are intraspecific variation.
Corethrella tigrina is morphologically very similar to C. solomonis , a species known as adult females, pupae and larvae from the Solomon Islands but the latter is more setose and does not have a subapical wing band. The discovery of this new species, and especially the discovery of the male, provides phylogenetic information regarding its position within the genus. The presence of unequal midleg claws on the male indicates that C. tigrina and C. mckeeveri Colless form an unresolved clade with the remainder of the drakensbergensis species group ( Borkent, 2008). Because of the striking similarity between the females of C. tigrina and C. solomonis , we have placed the latter species in this species group, considering it likely that the male of C. solomonis , once discovered will also have unequal midleg claws. These modified relationships are depicted in Fig. 19 View FIGURE 19 .
The male genitalia of C. tigrina is very similar to that of C. mckeeveri but differs in having the parameres tapering more abruptly near its apex. In addition, the apex of the gonostylus tapers more gradually than that of C. mckeeveri .
Corethrella tigrina has a subapical band on the wing that was considered a synapomorphy of the Neotropical quadrivittata species group by Borkent (2008). Here it is clearly an instance of homoplasy (i.e. C. tigrina is a member of the drakensbergensis species group and lacks the series of synapomorphies leading to the quadrivittata species group).
Males and females of C. tigrina have a small pale area posterior to the prescutal suture, somewhat similar to the derived condition described in character 28 ( Borkent, 2008). It is either a precursor or a partially homoplastic expression of the derived condition. Because we did not reexamine material of many of the early lineages of Corethrella , we cannot confidently interpret this observation.
TYPES: Holotype, male adult on microscope slide, labeled " HOLOTYPE Corethrella tigrina Borkent and Grafe ", " 15 km S. Liang, Brunei, 7-VI-2007; frog-call trap with synthetic tone; U. Grafe; 075-5" (CNCI).
Allotype, female adult on microscope slide, labeled as for holotype but "75-5/1". Paratypes on microscope slides: 1 Ƥ from type locality (UBD); 3 Ƥ, 12 km S. Liang, Brunei, 30 m, 8-VI-2007, 11-VI-2007, 23-V-2007, respectively (2, CNCI; 1 UBD); 1 Ƥ, Lanjak Entimau (112° 4'E, 1° 28'N), Sarawak, Malaysia, 80 m, 18-VI-2008 (CNCI); 4 Ƥ, from previous locality but 22-VI-2008 (CNCI); 1 Ƥ, from previous locality but 19-VI-2008 (CNCI). Paratypes on pins: 4 Ƥ, 12.5 km S. Liang, Brunei, 33 m, 30-I-2009 (CNCI); 1 Ƥ, from previous locality but 21-II- 2009 (CNCI); 1 Ƥ, from previous locality but 23-III-2009 (UBD); 1 Ƥ, 12 km S. Liang, Brunei, 30 m, 15-V-2011 (CNCI); 1 Ƥ, from previous locality but 10-I-2009 (CNCI); 4 Ƥ, KBFSC-walkway, Ulu Temburong NP, Brunei, 23-VII-2008 (CNCI); 6 Ƥ, Sungai Mata Ikan, Ulu Temburong NP, Brunei, 110 m, 10-VII-2008 (5, CNCI; 1 UBD); 3 Ƥ, from previous locality but 12-VII-2008 (CNCI); 4 Ƥ, Sungai Baki, Ulu Temburong NP, Brunei, 120 m, 18- VII-2008 (CNCI); 1 Ƥ, lower Sungai Apan Ulu Temburong NP, Brunei, 110 m, 13-VII-2008 (CNCI); 1 Ƥ, from previous locality but 8-VII-2008 (CNCI); 2 Ƥ, Belalong Münd (estuary), Ulu Temburong NP, Brunei, 4-VII-2008 (CNCI); 12 Ƥ, 0.5 km S. Kuala Belalong Field Studies Centre, along Sungai Esu, Brunei, 60 m, 12-XI-2010 (CNCI); 1 Ƥ, Ashton Trail in the Ulu Temburong NP, Brunei, 220 m, 24-V-2010 (CNCI); 1 Ƥ, from previous locality but 15-VI-2010 (CNCI); 2 Ƥ, Gunung Mulu NP, Sarawak, Malaysia, 30 m, 7-VII-2008 (CNCI); 12 Ƥ, Lanjak Entimau,(112° 4'E, 1° 28'N), Sarawak, Malaysia, 80 m, 22-VI-2008 (CNCI); 1 Ƥ, from previous locality but 25-VI-2008 (CNCI); 1 Ƥ, from previous locality but 24-VI-2008 (CNCI); 2 Ƥ, from previous locality but 21- VI-2008 (CNCI).
DERIVATION OF SPECIFIC EPITHET: The name tigrina (tiger) refers to the patterned thorax and legs of females of this species.






No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |