
Hamacantha (Zygherpe) hyaloderma
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3926.1.3 |
|
publication LSID |
lsid:zoobank.org:pub:CE5964E2-751F-466A-BC1B-6C16C009F6A8 |
|
DOI |
https://doi.org/10.5281/zenodo.5665688 |
|
persistent identifier |
https://treatment.plazi.org/id/914287DC-6359-FFE5-FF76-FA8FFCAEFB64 |
|
treatment provided by |
Plazi |
|
scientific name |
Hamacantha (Zygherpe) hyaloderma |
| status |
|
Hamacantha (Zygherpe) hyaloderma (de Laubenfels, 1932)
Figs 2 View FIGURE 2 B, 4, Table 1 View TABLE 1
Material studied. CZA 12146 (fragments deposited in MNRJ 12146, RBINS 12146, MHNG INVE 76161), Quebrada Ancupita (16°50’13.3’’ S – 72°17’28.3’’ W), Matarani, Peru, 3.7 m depth, coll. Y. Hooker & U. Zanabria, 27.xi.2008. CZA 12160 (fragments deposited in MNRJ 12160, RBINS 12160, MHNG INVE 76163), Bahia Quilca (16°42’06.10’’ S – 72°26’54.0’’ W), Quilca, Peru, 10.4 m depth, coll. Y. Hooker & M. Vilchez, 30.xi.2008. CZA 12162 (fragments deposited in MNRJ 12162, RBINS 12162, MHNG INVE 76160), Bahia Quilca (16°42’06.10’’ S – 72°26’54.0’’ W), Quilca, Peru, 8.0 m depth, coll. Y. Hooker & M. Vilchez, 30.xi.2008.
Comparative material: Holotype, USNM 22060, Point Lobos (California, USA), 13.vii.1930, det. M.W. de Laubenfels. MNRJ 3428, Kanuka Bay (San Juan Island, Washington, USA), 25.iv.1959, det./leg. G.J. Bakus. MNRJ 14504, Samsun Narrrows opposite Bold Bluff (British Columbia, Canada), 06.v.2008, det./leg. W.C. Austin. CNPGG 363, Bahía Tortugas (Baja California Sur , Mexico), 15.iv.1997, det. P. Gómez.
Diagnosis. This is the only encrusting Hamacantha with tylostyles, regular diancistras and two categories of smooth sigmas.
Description ( Fig. 2 View FIGURE 2 B). All three specimens collected were encrusting over granitic boulders, and were about 1 mm thick. The largest one, CZA 12162 covered approximately 20 x 10 cm in area. The sponge is yellow (CZA 12146) or orangy-yellow (CZA 12162) alive, and becomes beige in ethanol. Its surface bears conspicuous meandering subectosomal canals, leading to a few scattered oscula up to 1 mm in diameter. Texture is smooth, and consistency somewhat fragile.
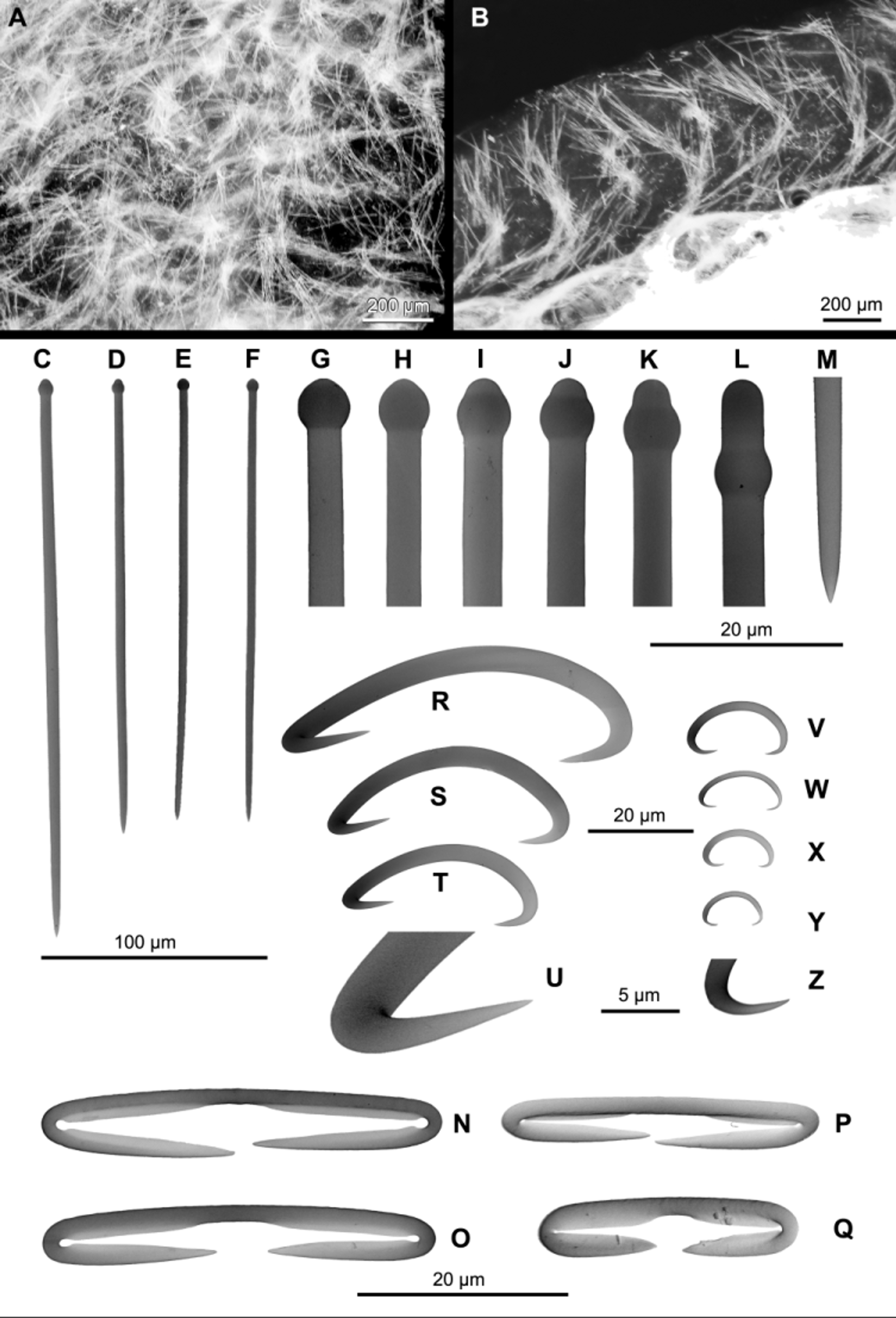
Skeleton ( Figs 4 View FIGURE 4 A–B). Ectosomal architecture unspecialized. Only scattered microscleres occur, and the wispy terminations of ascending choanosomal tracts ( Fig. 4 View FIGURE 4 A). Choanosomal architecture with sinuous ascending wispy tracts of megascleres ( Fig. 4 View FIGURE 4 B).
Spicules ( Figs 4 View FIGURE 4 C–Z, Table 1 View TABLE 1 ). Megascleres. Tylostyles ( Figs 4 View FIGURE 4 C–M), smooth, mostly slender and straight, heads well pronounced, usually spherical, frequently subterminal, 149–240 / 4.4–6.1 Μm. Microscleres. Diancistras ( Figs 4 View FIGURE 4 N–Q), small, smooth, mostly contorted, with conspicuous notches, hooks run parallel to axis which has fimbriae on both apical thirds, 26–38 Μm. Sigmas I ( Figs 4 View FIGURE 4 R–U, uncommon), relatively stout, smooth, contorted, 30–89 Μm. Sigmas II ( Figs 4 View FIGURE 4 V–Z, abundant), relatively stout, smooth, contorted, 14–23 Μm.
Distribution and ecology. Specimen CZA 12162 was partly epibiotic over a gastropod, and several cirripeds and polychaete tubes. In Peru the species was found between 3.7 and 10.4 m depth, between Matarani (16°42’S) and Quilca (16°50’S). Water temperature in the collecting sites was 14-15°C. The species appeared to be one of the dominant members of the community in shaded habitats in the Quebrada Ancupita. This species was reported by Goddard (1983) to be common in Oregon and one of the preferred items in the diet of the opisthobranch mollusc Montereina nobilis MacFarland, 1905 .
Remarks. Hamacantha hyaloderma was originally reported from California (de Laubenfels, 1932; Lee et al., 2007), and subsequently found in Washington ( Bakus, 1966), Oregon ( Goddard, 1983) and British Columbia ( Austin et al., 2012), thus suggesting the species to be of cold temperate affinity. The exception to this was Carreón-Palau et al. ’s (2003) record from Lower California. Surprisingly, given the large geographic gap, we were able to find this species in southern Peru. In the absence of molecular data to show the contrary, we are confident to determine both SE and NE Pacific specimens as conspecific. Table 1 View TABLE 1 compares micrometric measurements obtained from the materials studied here and those available in the literature (or re-evaluated), and together with Fig. 5 View FIGURE 5 , shows how similar SE and NE Pacific materials are in terms of spicule dimensions. On the other hand, the morphologic variability compiled for NE Pacific specimens might be hiding an underlying genetic structure. For instance, the British Columbia specimen analyzed has abundant small sigmas (13–23 µm), and exceedingly rare large ones (> 46 µm). This is markedly distinct from what is observed in the species’ holotype, where rather small sigmas (<23 µm) appear to be absent. If there is a hidden species complex, the Peruvian specimens might quite likely pertain to a new species given their apparent isolation.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |