
Weberella bursa Vosmaer, 1885
|
publication ID |
https://doi.org/10.11646/zootaxa.4951.1.12 |
|
DOI |
https://doi.org/10.5281/zenodo.4681132 |
|
persistent identifier |
https://treatment.plazi.org/id/85294C5A-4423-FFD2-FF5C-FAE6FB3FFD26 |
|
treatment provided by |
Plazi (2021-04-01 08:12:46, last updated 2024-11-29 17:58:16) |
|
scientific name |
Weberella bursa Vosmaer, 1885 |
| status |
|
Figs 2a–c View FIGURE 2 , 3a–f View FIGURE 3
Weberella bursa Vosmaer, 1885: 16 , pl. I figs 9, 12, 19, pl. III figs 6–8, 15– 20, 26–28; Topsent, 1928: 149; Uriz, 1975: 440; Boury-Esnault et al., 1994: 76, fig. 51; Van Soest, 2001: 89; Plotkin, 2004: 544, figs 1k, 2k; Plotkin et al. 2012: 25, fig. 1j.
Polymastia bursa ( Müller, 1806) ; Koltun, 1964: 149; Koltun, 1966: 76, pl. IX fig. 2, pl. 23 figs 1–2, pl. 24: figs 1–2, text-fig. 49.
Weberella bursa ( Müller, 1806) : Boury-Esnault, 2002: 214, fig. 11; Plotkin et al. 2018: 1324, fig. 36; (not: Alcyonium bursa? sensu Müller, 1806: 19 , pl. 98 figs 1–2)
Material examined. Lectotype RMNH Por. 12422, Norway, Finnmark, Barents Sea, Willem Barents Expedition 1881, stat. 08, 72.6014°N 24.9514°E, depth 252–263 m, coll. M. Weber, 28 June 1881 (specimen of 8 x 6 x 5 cm ( Fig. 2a View FIGURE 2 ) taken from a collective sample, RMNH Por. 706 (cf. Fig. 2c View FIGURE 2 ), indicated as ‘type’ by Vosmaer. GoogleMaps
Paralectotypes, six remaining specimens in RMNH Por. 706 ( Figs 2b–c View FIGURE 2 ) including the sectioned specimen shown in Fig. 2b View FIGURE 2 , same data as the lectotype GoogleMaps ; ZMA Por. 02419, Norway, Finmark, Barents Sea , Willem Barents Expedition 1881, no station data, approximately 72°N 36°E, coll. M. Weber GoogleMaps ; ZMA Por. 02420, Norway, Finnmark, Barents Sea, Willem Barents Expedition 1881, stat. 06, 70.6667°N 31.1667°E, hard bottom, depth 238 m, coll. M. Weber, 24 June 1881 GoogleMaps ; ZMA Por. 02691, Norway, Finnmark, Barents Sea, Willem Barents Expedition 1881, stat. 08, 72.6014°N 24.9514°E, depth 252–263 m, coll. M. Weber, 28 June 1881 GoogleMaps .
Additional non-type material: ZMA Por. 03256, Iceland, south coast, approximately 64°N 13°W, coll. L.C. Langenbroek, on board s.s.’ Anna’, June 1927 GoogleMaps ; ZMA Por. 03257, same data GoogleMaps ; ZMA Por. 03297, North East Atlantic, no further data, coll. Oldmark, on board s.s.’ Alkmaar’, April 1938 ; ZMA Por. 03300, Iceland, 63.35°N 19.50°W, coll. Van der Duin, no further data GoogleMaps .
Description. Colour in alcohol: yellowish to grey externally; dirty white inside.
Shape, size, surface and consistency ( Figs 2a–c View FIGURE 2 ): Globular to massive, covered with papillae. Size up to 13 cm in diameter, but usually smaller, 2–10 cm (holotype 8 x 6 x 5 cm, cf. Fig. 2a View FIGURE 2 ). Consistency compact, solid, firm. Papillae conico-cylindrical, up to 5 mm high, 2 mm in diameter. Normally there are several dozens of papillae in a single specimen, but their number depends on size of the sponge. Most papillae bear a terminal osculum, those without visible opening are contracted. At first glance the papillae are more or less evenly distributed over the sponge, although they may be more numerous on the upper body part. Some papillae may occasionally be grouped in small clusters or even fused together. Just few, if any, ostia are visible on the walls of papillae. The sponge surface between the papillae is velvety, with regularly distributed ostia.
Skeleton ( Figs 3a–d View FIGURE 3 ): Between the papillae, there is a distinct cortex of up to 1.5 mm in thickness ( Figs 3a–b View FIGURE 3 ), strengthened by a 200–300 µm thick palisade of small tylostyles ( Fig. 3b View FIGURE 3 ), underneath of which there are subdermal lacunae amid tissues lacking choanocytes, overlying a mass of intercrossing single large tylostyles, here and there united in bundles making an irregular subectosomal reticulation. In the choanosome the skeleton is made of large tylostyles arranged in an irregular reticulation of vague bundles with many loose single spicules. The skeleton of the papillae ( Figs 3c–d View FIGURE 3 ) is more compressed, with subdermal reticulation largely absent, although the intercrossing spicule layer is clearly present ( Fig. 3d View FIGURE 3 ).
Spicules ( Figs 3e–f View FIGURE 3 ): (Tylo-)styles in two clear categories, small tylostyles of the ectosome ( Fig. 3e View FIGURE 3 ), slightly curved, rounded heads, 133– 166.2 –232 x 2– 2.73 – 4.8 µm (overall measurements from various reliably identified specimens 125– 270 x 2–6.5 µm), and large subtylostyles ( Fig. 3f View FIGURE 3 ) of the subdermal and choanosomal region, slightly fusiform, with ovoid, often mucronate, not very prominent heads, 402– 518.8 –652 x 9.8– 11.7 – 13 µm (overall measurements from various reliably identified specimens 330–700 x 5–16 µm).
Ecology: Attached on rocks and stones, from the upper sublittoral zone down to deep water in shelf and upper bathyal habitats, depth range 16–932 m ( Plotkin et al. 2018; and pers. comm.).
Distribution: From the Arctic Ocean and Nordic Seas to the boreal North Atlantic: Russia (Franz Josef Land, Novaya Zemlya and Murman Coast), Norway ( Svalbard, Bear Island, and along the mainland coast from Finnmark to Trøndelag), Iceland, South Greenland, Atlantic coast of France, west of Gibraltar, north of the Azores, Eastern Canada.
Etymology: bursa (Latin) = purse or bag, referring to the shape of the sponge. The genus name honours Dr Max Weber, director of the Zoological Museum of Amsterdam around the turn of the 20 th century, leading scientist of the Siboga Expedition (1899–1900) to Indonesian waters, and author of several papers on sponges.
Remarks. The combination of a globular shape, short surface papillae, solid consistency and an irregularly arranged choanosomal reticulation make this a very distinctive species. Apart from more northern localities, including a misidentification of Burton (1959) as Polymastia uberrima from Iceland, it has been reported convincingly from the mouth of the river Loire ( Topsent, 1928, depth 150–932 m) and from the western inlet to the Strait of Gibraltar ( Boury-Esnault et al. 1994, depth 133–137 m).
In the Western Mediterranean, a second species of Weberella is found, W. verrucosa Vacelet, 1960 (cf. also Vacelet, 1969), smaller in size, with only a few, if any, papillae and slightly smaller spicules. Uriz (1975) made an extensive comparison between Topsent’s (1928) record of W. bursa and a large number of W. verrucosa specimens. We can confirm that the spicules of W. bursa are larger than those of W. verrucosa , although there is an overlap in length and thickness. Boury-Esnault et al. (1994) found the two species occurring sympatrically west of the Strait of Gibraltar.
Samaai & Gibbons (2005) described Weberella namibiensis from southern Africa. It has less prominent papillae and its large subtylostyles exceed those of W. bursa in length, averaging 782 µm.
Austin et al. (2014) described Weberella perlucida from shallow waters (10 m) off British Columbia, Western Canada. It differs quite strongly in shape, with numerous semi-transparent papillae and a densely confused skeleton. The larger tylostyles differ from W. bursa in having distinct tyles and being considerably thicker averaging 20 µm.
Austin, W. C., Ott, B. S., Reiswig, H. M., Romagosa, P. & McDaniel, N. G. (2014) Taxonomic review of Hadromerida (Porifera, Demospongiae) from British Columbia, Canada, and adjacent waters, with the description of nine new species. Zootaxa. 3823 (1), 1 - 84. https: // doi. org / 10.11646 / zootaxa. 3823.1.1
Boury-Esnault, N., Pansini, M. & Uriz, M. J. (1994) Spongiaires bathyaux de la mer d'Alboran et du golfe ibero-marocain. Memoires du Museum national d'Histoire naturelle, 160, 1 - 174.
Boury-Esnault, N. (2002) Family Polymastiidae Gray, 1867. In: Hooper, J. N. A. & Van Soest, R. W. M. (Eds.), Systema Porifera. A guide to the classification of sponges. Kluwer Academic / Plenum Publishers, New York, Boston, Dordrecht, London, Moscow, xviii, 1708, xix - xlviii pp. [pp. 201 - 219] https: // doi. org / 10.1007 / 978 - 1 - 4615 - 0747 - 5
Burton, M. (1959) Spongia. The Zoology of Iceland, 2 (3 - 4), 1 - 71.
Koltun, V. M. (1964) Sponges (Porifera) collected in the Greenland seas and from the region to the north of the Spitzbergen and Franz Josef Land, from expeditions of the ' F. Litke' 1955, ' Obb' 1956 and ' Lena' 1957 and 1958. Scientific results of the higher latitudes. Oceanographic Expeditions to the northern part of the Greenland Sea and the adjacent arctic basin. Publications Arctic Antarctic Scientific Institute, 259, 143 - 166. [in Russian]
Koltun, V. (1966) Four-rayed sponges of Northern and Far Eastern seas of the USSR (order Tetraxonida). (Zoological Institute of the Academy of Sciences of the USSR: Moscow, Leningrad). Opredeliti Faunei SSSR (Keys for the identification of the fauna of the USSR), 90, 1 - 112. [in Russian]
Muller, O. F. (1806) Zoologia Danica seu animalium Daniae et Norvegiae rariorum ac minus notorum Descriptiones et Historia. Volumen quartum. N. Christensen, Havniae, 5 + 46, pls. 121 - 160 pp.
Plotkin, A. S. (2004) Biodiversity and distribution of Polymastiidae (Demospongiae, Hadromerida) in the Arctic area. Bollettino dei Musei e degli Istituti Biologici dell'Universita di Genova, 68, 535 - 547.
Plotkin, A., Gerasimova, E. & Rapp, H. T. (2012) Phylogenetic reconstruction of Polymastiidae (Demospongiae: Hadromerida) based on morphology. In: Maldonado, M., Turon, X., Becerro, M. A. & Uriz, M. J. (Eds.), Ancient Animals, New Challenges, Sponge Research Developments. Hydrobiologia, 687, 21 - 41. https: // doi. org / 10.1007 / s 10750 - 011 - 0823 - 0
Plotkin, A., Gerasimova, E. & Rapp, H. T. (2018) Polymastiidae (Porifera: Demospongiae) of the Nordic and Siberian Seas. Journal of the Marine Biological Association of the United Kingdom, 98 (6), 1273 - 1335. https: // doi. org / 10.1017 / S 0025315417000285
Samaai, T. & Gibbons, M. J. (2005) Demospongiae taxonomy and biodiversity of the Benguela region on the west coast of South Africa. African Natural History, 1, 1 - 96.
Topsent, E. (1928) Spongiaires de l'Atlantique et de la Mediterranee provenant des croisieres du Prince Albert ler de Monaco. Resultats des campagnes scientifiques accomplies par le Prince Albert I. Monaco, 74, 1 - 376, pls. I - XI.
Uriz, M. J. (1975) Weberella verrucosa Vacelet 1960 (Demospongia), especie nueva para Espana. Investigacion pesquera, 39 (2), 429 - 442.
Vacelet, J. (1960) Eponges de la Mediterranee nord-occidentale recoltees par le ' President Theodore Tissier' (1958). Revue des Travaux de l'Institut des Peches maritimes, 24 (2), 257 - 272.
Vacelet, J. (1969) Eponges de la Roche du Large et de l'etage bathyal de Mediterranee (Recoltes de la soucoupe plongeante Cousteau et dragages). Memoires du Museum national d'Histoire naturelle (A, Zoologie). 59 (2), 145 - 219, pls. I - IV.
Van Soest, R. W. M. (2001) Porifera. In: Costello, M. J., Emblow, C. & White, R. (Eds.), (2001). European register of marine species: a check-list of the marine species in Europe and a bibliography of guides to their identification. Collection Patrimoines Naturels. 50. Museum national d'Histoire Naturelle: Paris, France, 463 pp. [pp. 85 - 103.]
Vosmaer, G. C. J. (1885) The Sponges of the ' Willem Barents' Expedition 1880 and 1881. Bijdragen tot de Dierkunde, 12 (3), 1 - 47, pls. I - V. https: // doi. org / 10.1163 / 26660644 - 01201001
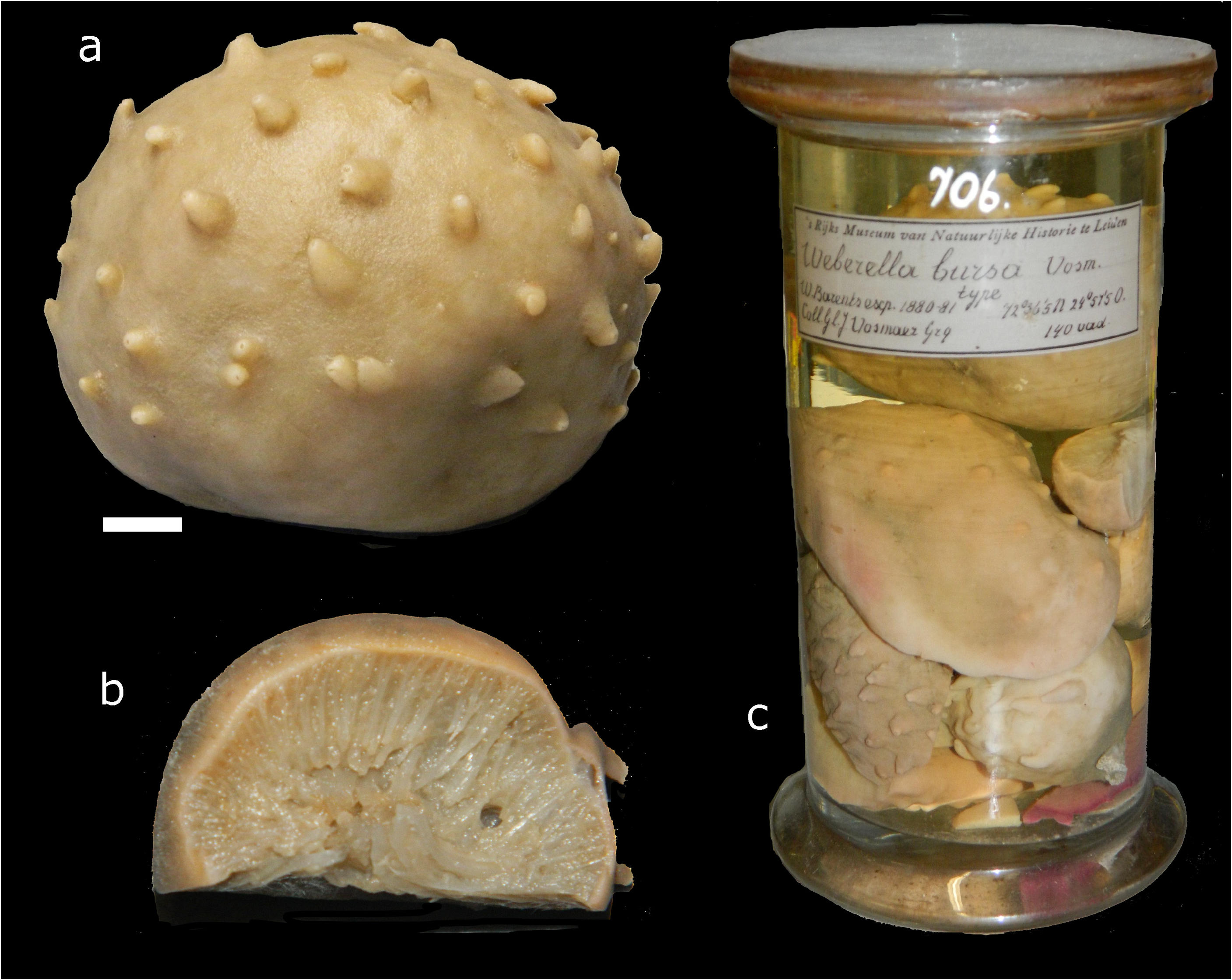
FIGURE 2. Specimens described by Vosmaer (1885) as Weberella bursa from the Barents Sea, incorporated in Naturalis Biodiversity Center, a. lectotype (proposed herein), RMNH Por. 12422 (scale bar = 1 cm), b, cross section of one of the paralectotypes to show inner structure (same scale as a), c, one of the sample bottles of Weberella bursa, RMNH Por. 706, from which the lectotype was selected (reduced scale).
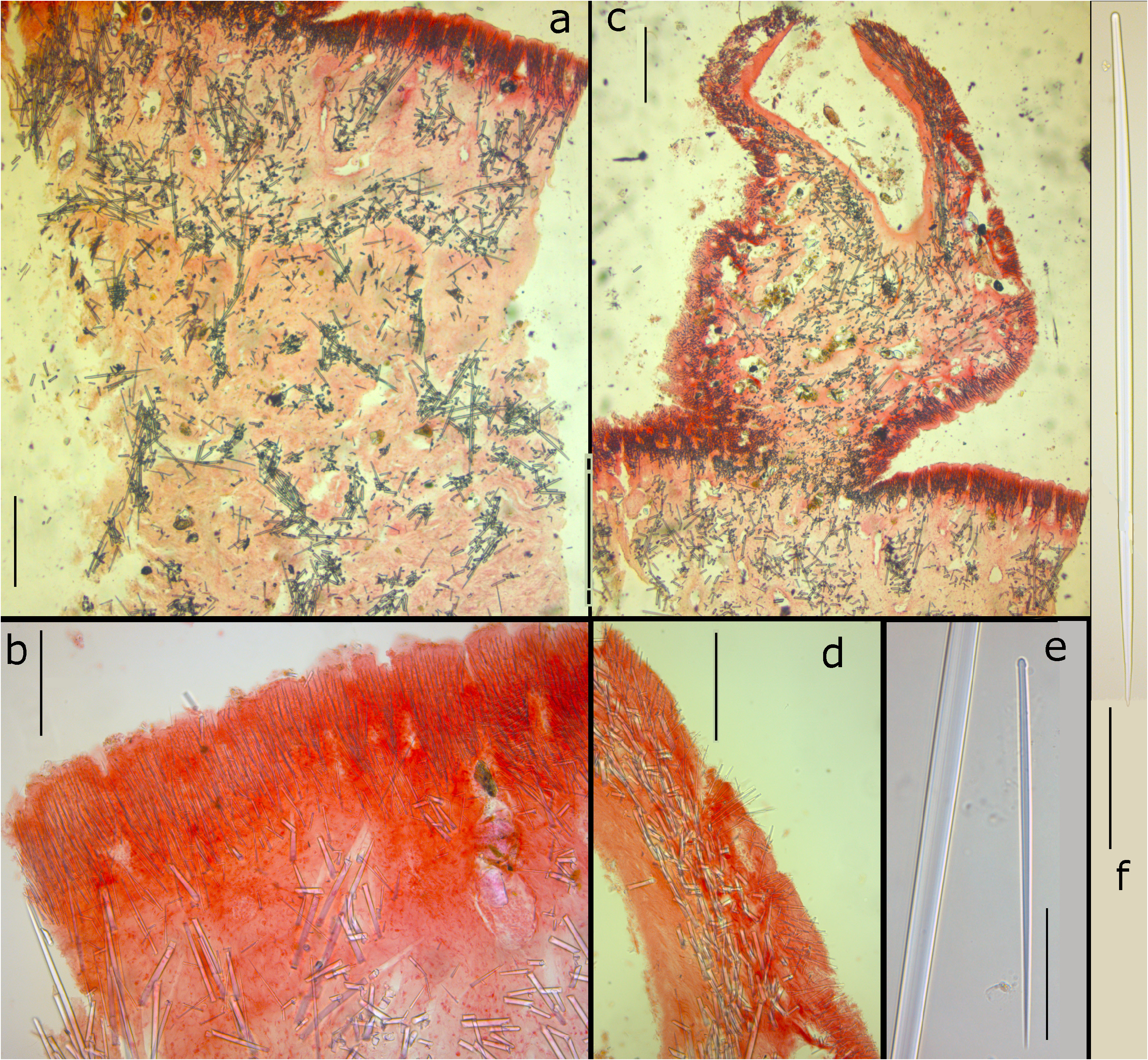
FIGURE 3. Sections and spicules of one of the paralectotypes of Weberella bursa, made by Vosmaer. a, cross section of main body showing ectosomal palisade of small tylostyles overlying a confused reticulation of choanosomal spicule bundles (scale bar = 500 µm), b, detail of peripheral palisade (scale bar = 200 µm), c, cross section of a papilla (scale bar = 500 µm), d, cross section of periphery of papilla showing ectosomal palisade overlying tangential mass of larger tylostyles forming the structure of the papilla wall (scale bar = 200 µm), e, small ectosomal tylostyle, set off against middle part of large subtylostyle to show size differences (scale bar = 50 µm), f, large subtylostyle (scale bar = 100 µm).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
SubClass |
Heteroscleromorpha |
|
Order |
|
|
Family |
|
|
Genus |
Weberella bursa Vosmaer, 1885
| Van Soest, Rob W. M. & Plotkin, Alexander 2021 |
Weberella bursa ( Müller, 1806 )
| Plotkin, A. & Gerasimova, E. & Rapp, H. T. 2018: 1324 |
| Boury-Esnault, N. 2002: 214 |
| Muller, O. F. 1806: 19 |
Polymastia bursa ( Müller, 1806 )
| Koltun, V. 1966: 76 |
| Koltun, V. M. 1964: 149 |
Weberella bursa
| Plotkin, A. & Gerasimova, E. & Rapp, H. T. 2012: 25 |
| Plotkin, A. S. 2004: 544 |
| Van Soest, R. W. M. 2001: 89 |
| Boury-Esnault, N. & Pansini, M. & Uriz, M. J. 1994: 76 |
| Uriz, M. J. 1975: 440 |
| Topsent, E. 1928: 149 |
| Vosmaer, G. C. J. 1885: 16 |
1 (by plazi, 2021-04-01 08:12:46)
2 (by ExternalLinkService, 2021-04-01 08:19:02)
3 (by ExternalLinkService, 2021-04-12 14:58:37)
4 (by ExternalLinkService, 2021-04-12 15:04:04)
5 (by ExternalLinkService, 2022-01-29 03:23:24)
6 (by plazi, 2023-11-02 04:26:14)