
Echiniscus blumi Richters, 1903
|
publication ID |
https://doi.org/10.11646/zootaxa.5344.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:DCF48473-AC31-4CDB-808F-453F8F280002 |
|
DOI |
https://doi.org/10.5281/zenodo.8349250 |
|
persistent identifier |
https://treatment.plazi.org/id/8119D633-B94D-FFF9-1CED-FD8BB81BFECB |
|
treatment provided by |
Plazi (2023-09-14 15:19:06, last updated 2023-09-15 15:14:28) |
|
scientific name |
Echiniscus blumi Richters, 1903 |
| status |
|
4. Echiniscus blumi Richters, 1903 View in CoL View at ENA
Figure 4A View FIGURE 4
Synonyms
Echiniscus bisetosus Heinis, 1908 : Gąsiorek et al. (2019b)
Echiniscus blumi schizofilus Bartoš, 1941 syn. nov.
Echiniscus mediantus Marcus, 1930 : Gąsiorek et al. (2019b)
Echiniscus ramazzottii Binda & Pilato, 1969 : Maucci (1985)
Echiniscus trojanus Maucci, 1973 syn. nov.
Terra typica: Spitsbergen (a general reference) and Merok ( Norway) ( Richters 1903); Klaas-Billen Bay, Isfjord, Spitsbergen ( Richters 1904b). The species description was published independently in two separate papers ( Richters 1903, 1904b).
Etymology: The name commemorates J. Blum. A noun in the genitive singular.
Shortened description: Medium-sized to large (ca. 250–450 μm, typically 300–400 μm). Body appendage configuration A-(B)- C-Cd- D-Dd- E ( Fig. 4A View FIGURE 4 ); generally cirrous appendages in positions B, C, Cd, D, and spines in positions Dd, E, but a variety of intermediate stages can be observed, with some appendages frequently absent or lacking asymmetrically. Overall, chaetotaxy unstable. Dorsal plate sculpturing comprises densely arranged pores with thick epicuticular edges in the form of polygons ( Fig. 5A View FIGURE 5 ). Pedal plates weakly sculptured. Subcephalic and genital plates may be present in sexually mature females. Claws large; internal ones with primary spurs directed downwards, and external ones with secondary and sometimes even tertiary spurs directed upwards.
Phylogenetic position: Echiniscus blumi is firmly located at the base of the Echiniscus phylogenetic tree (Figs 1–2).
Remarks:Problems with species delimitation in the blumi-canadensis complex are almost as old as tardigradology itself ( Ramazzotti & Maucci 1982), and constitute a prime example of how fallacious the sole usage of chaetotaxy can be in the Echiniscidae . Guil (2008) summarised all transitional forms between morphotypes recognised as species at that time, concluding that basically they represent a morphological continuum, which agrees with findings of previous researchers ( Ramazzotti & Maucci 1983; Maucci 1985). A single molecular study on the populations from central Iberian Peninsula revealed the discordance between morphology and COI barcode, leading to a suspicion that cryptic speciation may take place in the case of this group ( Guil & Giribet 2009). Based on ample comparative material ( Argentina, Canary Islands, France, Georgia, Greenland, Iceland, Italy, Kazakhstan, Kyrgyzstan, North America, South Africa, Spain, Svalbard, Tanzania) deposited in our Department, we hypothesise that it is unlikely that E. canadensis Murray, 1910 and E. trisetosus Cuénot, 1932 will stand the test of time, but we temporarily left them in the list of valid species for two reasons: (1) type locality of E. canadensis is extra-European, and should be sampled in order to obtain DNA barcodes; and (2) these are the only two species of the complex that can be found as populations independent from the admixture of E. blumi . Irrespectively of the status of both species, we express our conviction that if there are any morphological differences between potentially cryptic lineages within the blumi-canadensis complex, they will be mostly quantitative, not qualitative. The application of geometric morphometry could aid in species delineation in such case ( Fontoura & Morais 2011). Echiniscus marleyi Li, 2007 nom. inq. also belongs in the blumi-canadensis complex and is probably synonymous with E. blumi , too. We establish two new synonymies for E. blumi . The first is E. blumi schizofilus Bartoš, 1941 syn. nov., described to accommodate individuals with doubled appendages, mainly in lateral positions, or with forked cirri. Such atypical forms are, however, common within populations of E. blumi , as there is a great variation in the development of cirri. Moreover, another example of a synonymous morphotype has already been established for a phenotype with aberrant appendages B (Maucci 1985). Consequently, E. blumi becomes a monotypic species. The second species is Echiniscus trojanus Maucci, 1973 syn. nov. ( Figs 5B–F View FIGURE 5 ), which could be differentiated from E. blumi based on the absence of cirri B and the development of spine E. Both traits are, however, variable within the blumi-canadensis complex, and cannot be treated as valid.
Bartos, E. (1941) Studien uber die Tardigraden des Karpathengebietes. Zoologische Jahrb ¸ cher Abteilung f ¸ r Systematik, Okologie und Geographie der Tiere, 74 (5 - 6), 435 - 472.
Binda, M. G. & Pilato, G. (1969) Tardigradi muscicoli delle Isole Eolie (Sicilia). Bollettino delle sedute dell'Accademia Gioenia di Scienze naturale Catania, Serie IV, 9, 634 - 651.
Cuenot, L. (1932) Tardigrades. In: Lechevalier, P. (Ed.), Faune de France, 24, pp. 1 - 96.
Fontoura, P. & Morais, P. (2011) Assessment of traditional and geometric morphometrics for discriminating cryptic species of the Pseudechiniscus suillus complex (Tardigrada, Echiniscidae). Journal of Zoological Systematics and Evolutionary Research, 49 (Supplement 1), 26 - 33. https: // doi. org / 10.1111 / j. 1439 - 0469.2010.00594. x
Gasiorek, P., Morek, W., Stec, D. & Michalczyk, L. (2019 b) Untangling the Echiniscus Gordian knot: paraphyly of the arctomys group (Heterotardigrada: Echiniscidae). Cladistics, 35 (6), 633 - 653. https: // doi. org / 10.1111 / cla. 12377
Guil, N. (2008) New records and within-species variability of Iberian tardigrades (Tardigrada), with comments on the species from the Echiniscus blumi-canadensis series. Zootaxa, 1757 (1), 1 - 30. https: // doi. org / 10.11646 / zootaxa. 1757.1.1
Guil, N. & Giribet, G. (2009) Fine scale population structure in the Echiniscus blumi-canadensis series (Heterotardigrada, Tardigrada) in an Iberian mountain range - When morphology fails to explain genetic structure. Molecular Phylogenetics and Evolution, 51 (3), 606 - 613. https: // doi. org / 10.1016 / j. ympev. 2009.02.019
Heinis, F. (1908) Tardigraden der Schweiz. Zoologischer Anzeiger, 32 (22), 633 - 638.
Li, X. (2007) Tardigrades from the Tsinling Mountains, central China with descriptions of two new species of Echiniscidae (Tardigrada). Journal of Natural History, 41 (41 - 44), 2719 - 2739. https: // doi. org / 10.1080 / 00222930701711046
Marcus, E. (1930) Beitrage zur Tardigradensystematik. Zoologische Jahrb ¸ cher Abteilung f ¸ r Systematik Okologie und Geographie der Tiere, 59, 363 - 386.
Murray, J. (1910) Tardigrada. British Antarctic Expedition 1907 - 1909. Reports on the Scientific Investigations, 1 (Biology, Part V), 83 - 187.
Ramazzotti, G. & Maucci, W. (1982) A history of tardigrade taxonomy. In: Nelseon, D. R. (Ed.), Proceedings of the Third International Symposium on the Tardigrada, 1980. East Tennessee State University Press, Johnson City, Tennessee, pp. 11 - 30.
Ramazzotti, G. & Maucci, W. (1983) Il Phylum Tardigrada. III edizione riveduta e aggiornata. Memorie dell'Istituto Italiano di Idrobiologia, 41, 1 - 1011.
Richters, F. (1903) Nordische Tardigraden. Zoologischer Anzeiger, 27 (5), 168 - 172.
Richters, F. (1904 b) Arktische Tardigraden. Fauna Arctica, 3, 495 - 508.
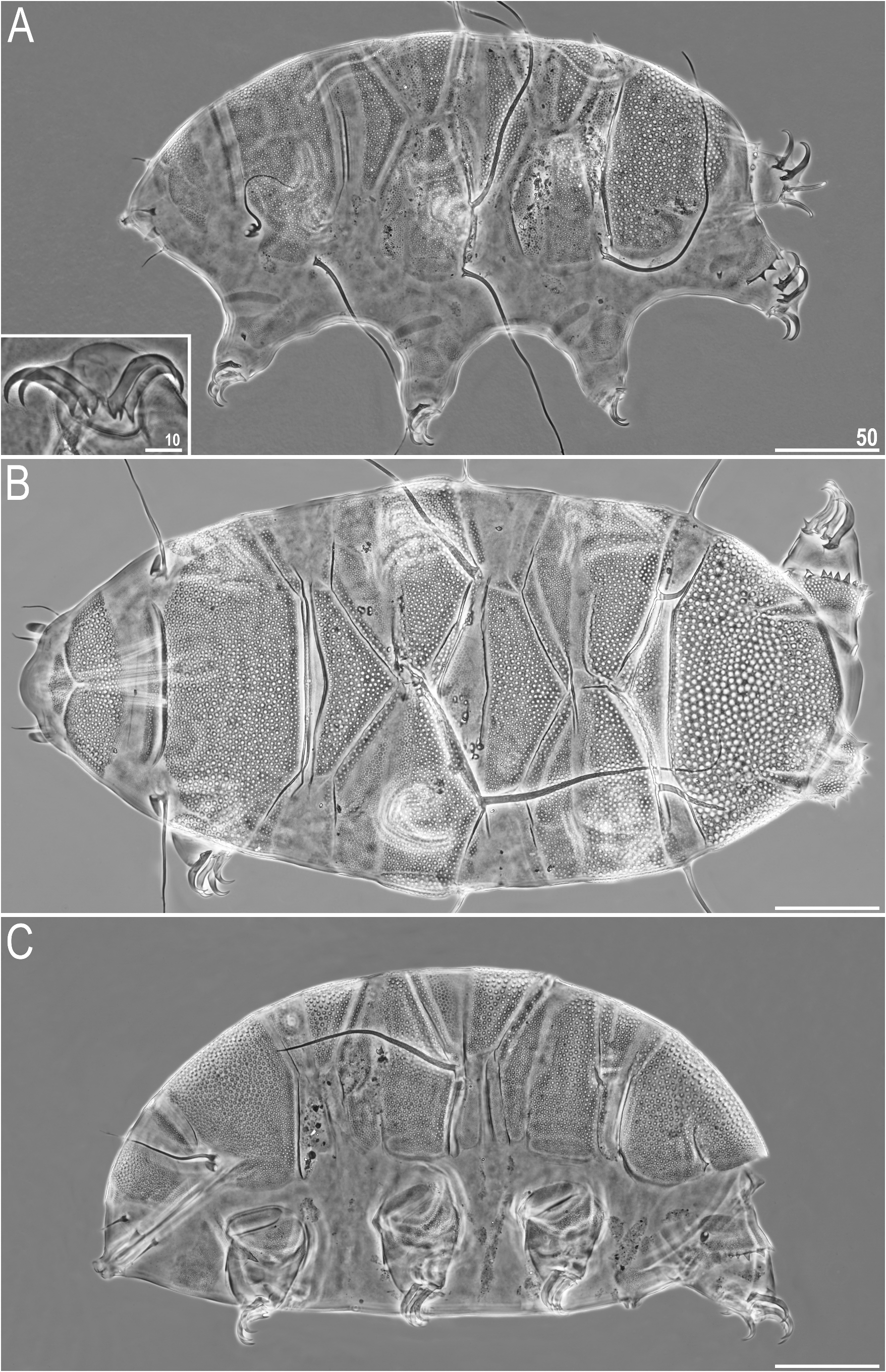
FIGURE 4. Echiniscus blumi-canadensis complex of species (PCM): A—E. blumi in dorsolateral view (insert shows claws III), B—E. trisetosus in dorsal view, C—E. canadensis in lateral view. Scale bars in μm.

FIGURE 5. Dorsal plate sculpturing of the blumi-canadensis complex (PCM): A—scapular plate of E. trisetosus, B—scapular plate of E. trojanus syn. nov. (paratype, upper magnification), C—scapular plate of E. trojanus syn. nov. (paratype, lower magnification), D—median plate 1 of E. trojanus syn. nov. (paratype), E—paired segmental plate I of E. trojanus syn. nov. (paratype), F—caudal (terminal) plate of E. trojanus syn. nov. (paratype), G—scapular plate of E. osellai syn. nov. (paratype), H—paired segmental plate I of E. osellai syn. nov. (paratype), I—caudal plate of E. osellai syn. nov. (paratype). Scale bars = 10 μm.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Echiniscus blumi Richters, 1903
| Gąsiorek, Piotr & Vončina, Katarzyna 2023 |
Echiniscus trojanus
| Maucci 1973 |
Echiniscus ramazzottii
| Binda & Pilato 1969 |
Echiniscus blumi schizofilus Bartoš, 1941
| Bartos 1941 |
Echiniscus mediantus
| Marcus 1930 |
Echiniscus bisetosus
| Heinis 1908 |