
Megalomys, TROUESSART, 1881
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2009.00628.x |
|
DOI |
https://doi.org/10.5281/zenodo.13887379 |
|
persistent identifier |
https://treatment.plazi.org/id/7C4E878A-AB7C-FF96-6302-E44508EF8F12 |
|
treatment provided by |
Valdenar |
|
scientific name |
Megalomys |
| status |
|
GENUS MEGALOMYS TROUESSART, 1881 View in CoL
(TYPE SPECIES MUS PILORIDES SAY, 1822 )
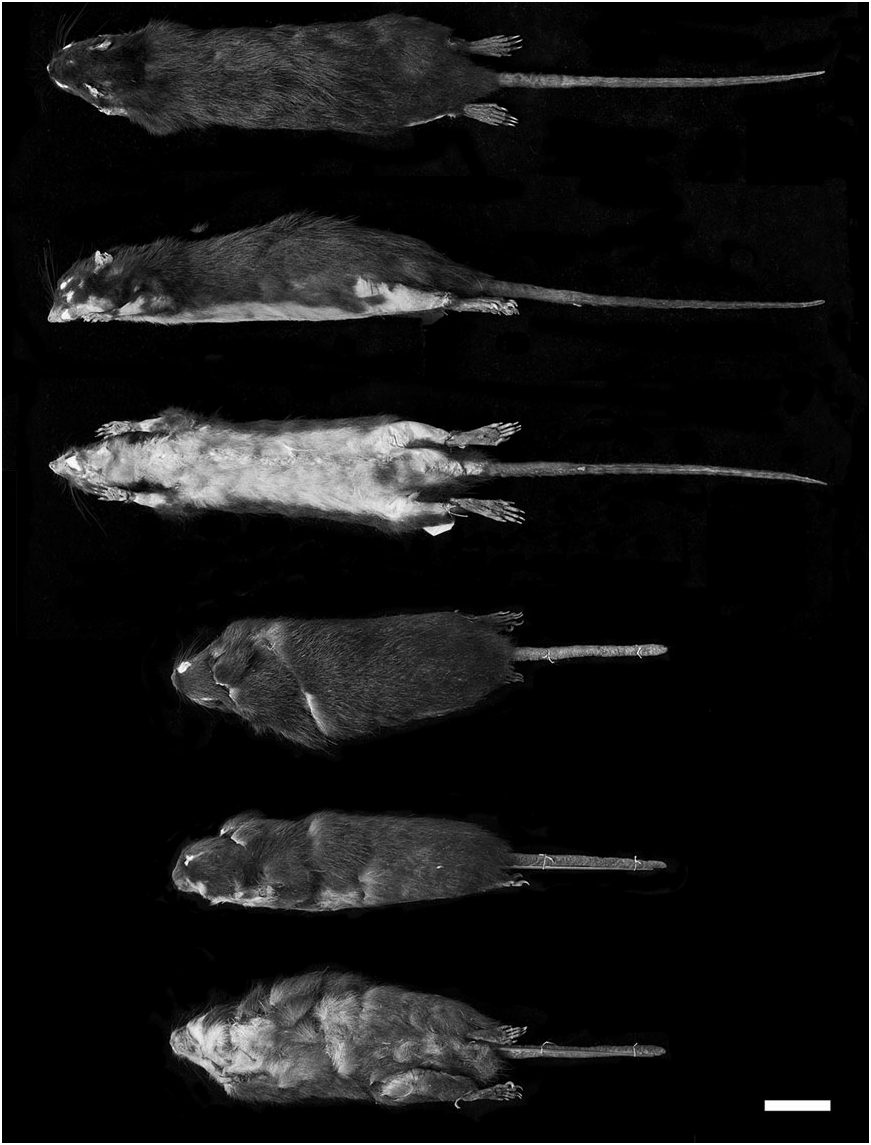
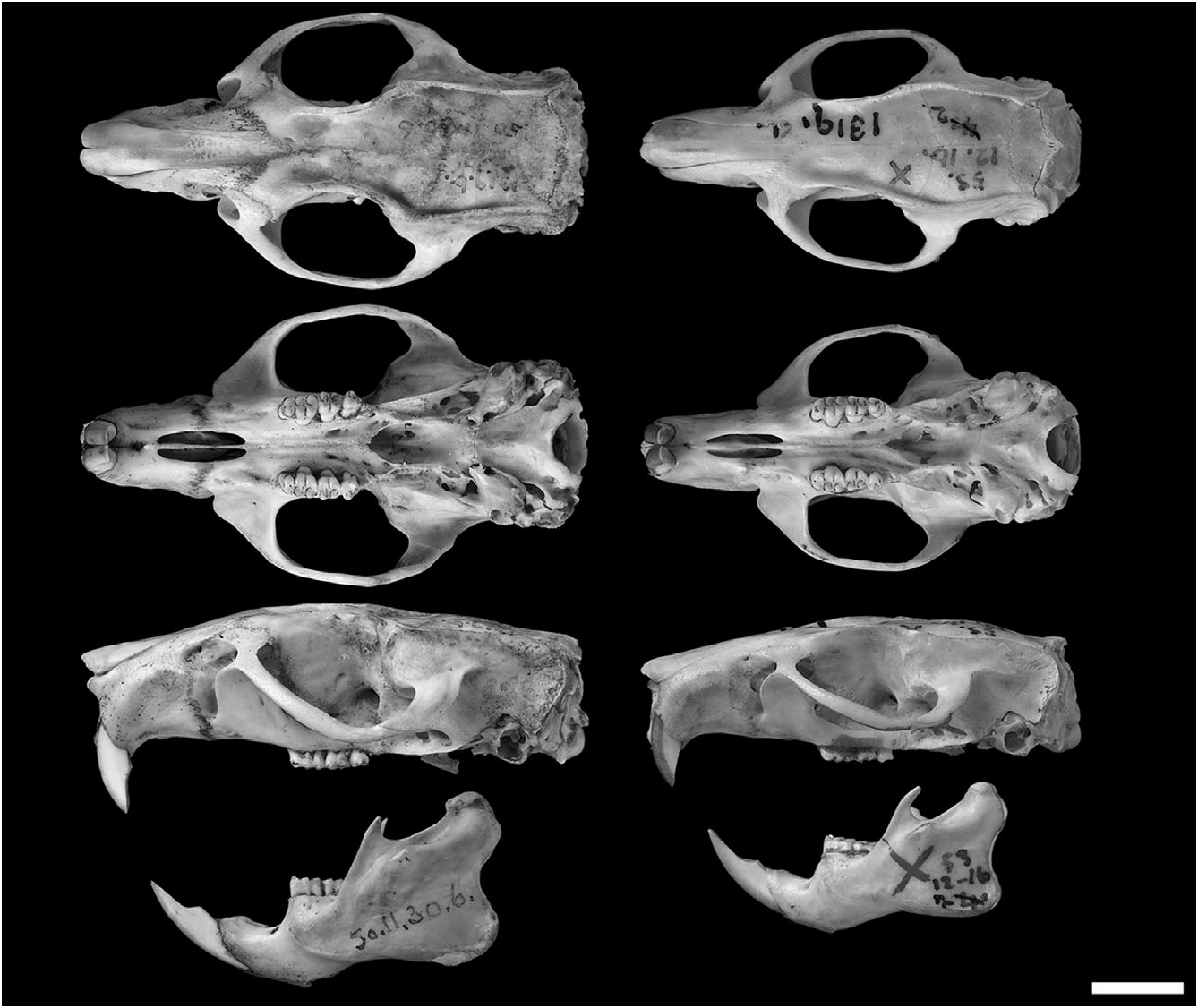
Examined material: Megalomys desmarestii : NHM 55.12.24.201 (skin and skull); NHM 50.11.30.5 (skin), and NHM 50.11.30.6 (skull and postcranial skeleton) – these represent the same individual.
Megalomys luciae : NHM 53.12.16.2 (holotype; damaged skin and skull) .
Megalomys audreyae : NHM M7406a (holotype; left mandibular ramus lacking m1); NHM M7406b (upper incisor).
Diagnosis: Megalomys can be differentiated from all other sigmodontine rodents by the combination of its extreme body size, oryzomyine synapomorphies (e.g. long palate, absence of both suspensory process of squamosal and alisphenoid struts), and several integumental, cranial, and dental traits, including nine synapomorphies (see below). Important characters include: pes without well-developed natatory fringes and interdigital webs, and with interdigital pads large and fleshy; robust skull with interorbital region convergent anteriorly, with well-developed supraorbital crests and squared braincase; zygomatic plate lacks an anterodorsal spinous process; small incisive foramina, not extending posteriorly between M1 alveoli; large posterolateral palatal pits recessed in deep fossae; stapedial foramen and posterior opening of alisphenoid canal small, and squamosal– alisphenoid groove and sphenofrontal foramen absent. Well-developed capsular process of lower incisor alveolus; and superior and inferior masseteric ridges joined anteriorly as an open chevron. Molars are bunodont, with M1 anterocone divided by a shallow anteromedian flexus; accessory rootlets are present on M1 and m1; m2 and m3 have three roots.
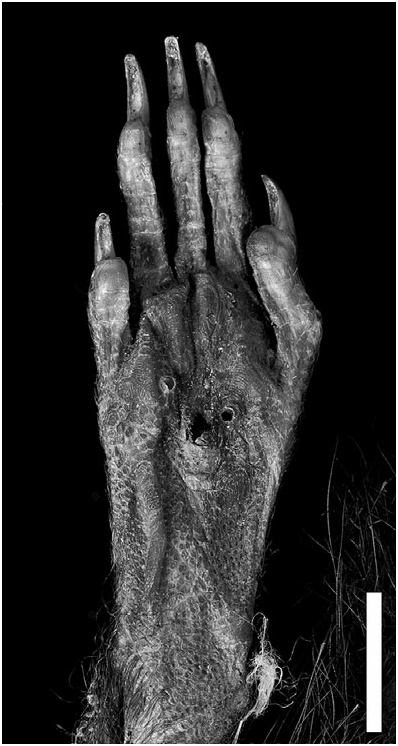
Description (based on M. desmarestii and M. luciae only): Very large rats, as large or larger than any extant oryzomyines ( Table 1 View Table 1 ). Dorsal pelage dark, blackish brown (almost black) (tan brown in one specimen in which the colour might have faded); ventral pelage abruptly paler in M. desmarestii , all white, except for a few darker guard hairs; in M. luciae , general ventral colour is dark, with plumbeous base, but there are distinct paler areas under the chin and throat and under the lower thorax, the two regions being separated by a streak of darker fur ( Fig. 3 View Figure 3 ). Soft fur, subauricular patches absent. Pinnae small, not reaching eye when laid forward. Mystacial and superciliary vibrissae not extending posteriorly beyond pinnae when laid back. Manual claws small and unkeeled. Hindfoot without well-developed natatory fringes and interdigital webs; tufts of ungual hairs at bases of claws on dII–dV vestigial or absent; plantar surface covered with distinct squamae distal to thenar pad; hypothenar pad absent or vestigial in M. luciae , present but reduced in M. desmarestii ; interdigital pads large and fleshy, pads 1 and 4 contiguous with 2 and 3; claw of digit I (dI) extending just beyond first interphalangeal joint of dII; claw of dV extending to middle of second phalange of dIV ( Fig. 4 View Figure 4 ). Tail about same length as head and body, sparsely haired and covered with conspicuous epidermal scales; without tuft of long terminal hairs, and unicoloured (dark). Mammary complement unknown.
Skull with stout rostrum flanked by moderate or deep zygomatic notches; interorbital region convergent anteriorly with well-developed supraorbital crests (symmetrically constricted in an older adult specimen of M. desmarestii ); braincase squared, with well-developed temporal crests; lambdoidal and nuchal crests well developed in older adults. Posterior margin of zygomatic plate dorsal to M1 alveolus; zygomatic plate lacks an anterodorsal spinous process. Jugal present and small (the maxillary and squamosal zygomatic processes overlap). Nasal bones with acutely angled posterior nasal margins; extending posteriorly behind lacrimals; lacrimals sutured mainly to maxillary. Posterior wall of the orbit with postorbital ridge (faint in M. luciae ). Frontosquamosal suture anterior to frontoparietal suture. Parietals with broad lateral expansions. Incisive foramina small, not extending posteriorly between M1 alveoli, spindle-shaped. Bony palate between the molar rows is smooth or weakly sculpted. Posterolateral palatal pits large, complex, and recessed in deep fossae; mesopterygoid fossa penetrating anteriorly between maxillae; bony roof of mesopterygoid fossa completely ossified in an older adult specimen of M. desmarestii , perforated by small (slit-like) sphenopalatine vacuities in a younger specimen of M. desmarestii and in M. luciae . Alisphenoid strut absent (buccinator– masticatory foramen and accessory foramen ovale confluent). Alisphenoid canal with large anterior opening. Stapedial foramen and posterior opening of alisphenoid canal small; squamosal–alisphenoid groove and sphenofrontal foramen absent; secondary anastomosis of internal carotid crosses dorsal surface of pterygoid plate (= carotid circulatory pattern 3 of Voss, 1988). Posterior suspensory process of the squamosal absent. Postglenoid foramen small and dorsoventrally compressed, covered by tegmen tympani; subsquamosal fenestra vestigial or absent. Periotic exposed posteromedially, between ectotympanic and basioccipital, extending anteriorly to carotid canal; mastoid unfenestrated. Capsular process of lower incisor alveolus well developed in adult specimens (smaller in M. luciae ); superior and inferior masseteric ridges joined anteriorly as open chevron below m1 (slightly anterior in M. desmarestii ) ( Fig. 5 View Figure 5 ).
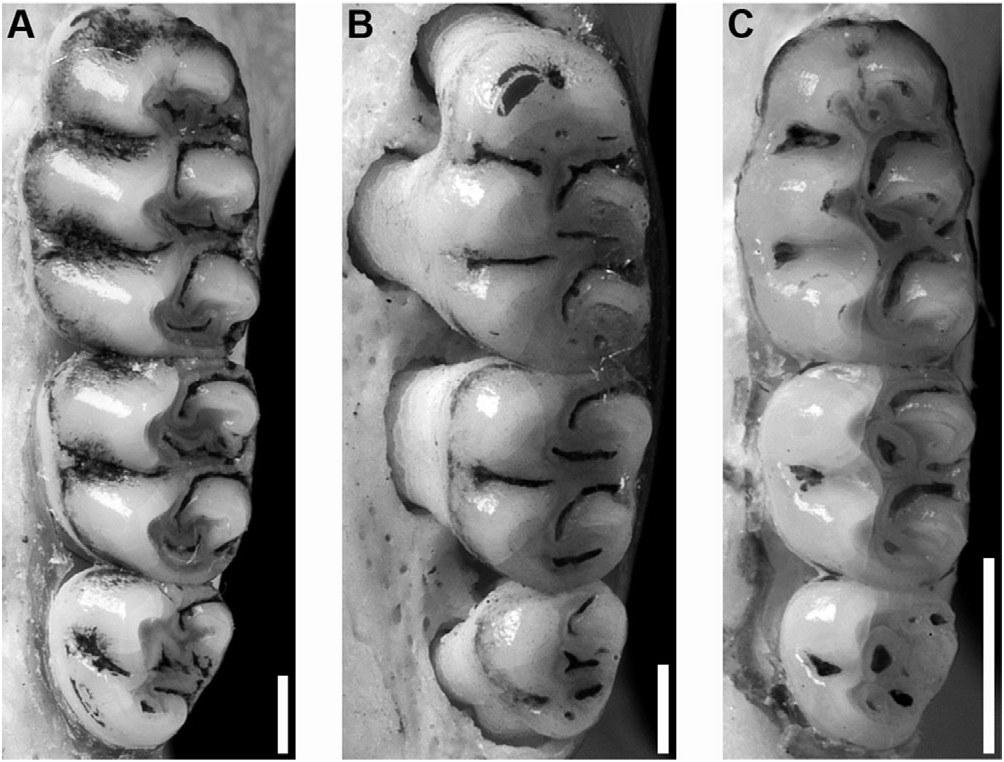
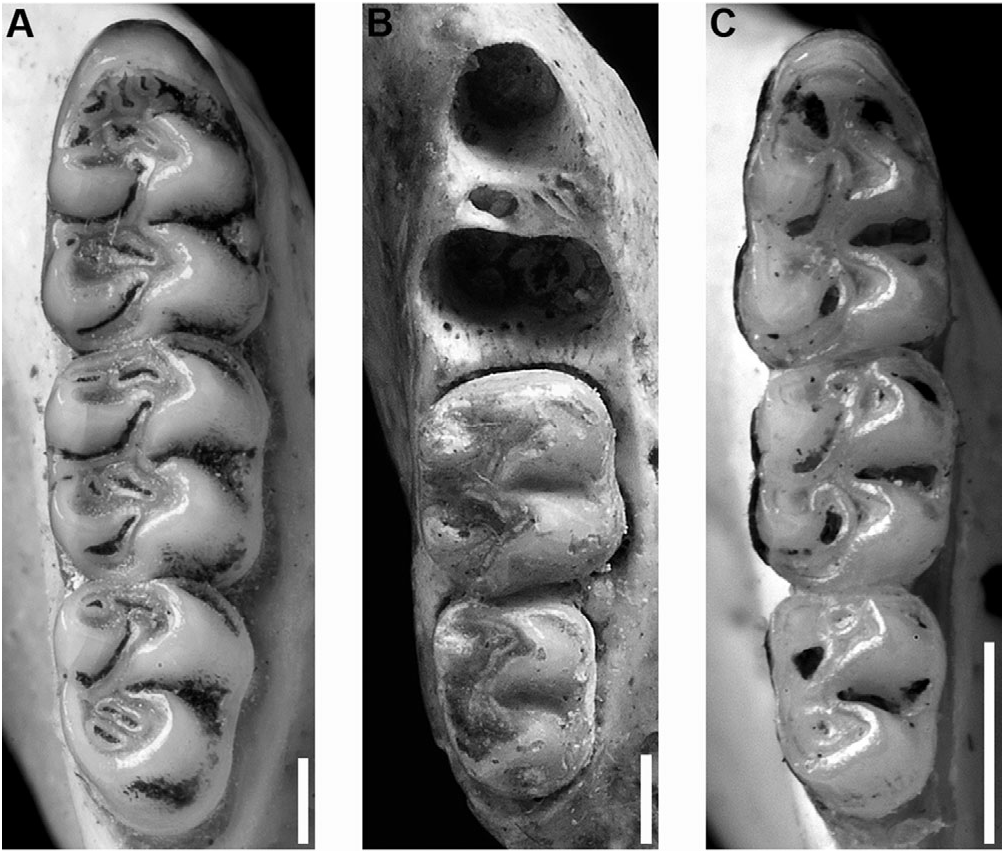
Upper incisors with smoothly rounded enamel bands. Maxillary tooth rows parallel. Molars bunodont and with labial flexi enclosed by a cingulum; labial and lingual flexi of M1 and M2 meet at midline, enamel overlaps. M1 anterocone divided into anterolabial and anterolingual conules by shallow anteromedian flexus; anteroloph well developed and fused with anterostyle on labial cingulum, separated from anterocone by persistent anteroflexus; protostyle absent; mesolophs present on all upper molars; paracone connected by enamel bridge to anterior or middle moiety of protocone; median mure connected to protocone. M2 protoflexus absent; mesoflexus present as single internal fossette; paracone without an accessory loph. M3 with posteroloph and developed hypoflexus (the latter remaining excavated with moderate to heavy wear) ( Fig 6A, B View Figure 6 ). Accessory labial root of M1 present.
m1 anteroconid without an anteromedian flexid; anterolabial cingulum present on all lower molars; ectolophid absent on m1 and m2; anterolophid present on m2 and m3; mesolophid distinct on unworn m1 and m2; m2 hypoflexid short; posteroflexid present on m3 ( Fig. 7A View Figure 7 ). Accessory lingual and labial roots of m1 present; m2 and m3 each with two small anterior roots and one large posterior root.
First rib with double articulation. Humerus without entepicondylar and supratrochlear foramina. Twelve ribs (false thirteenth rib present in older adult specimen). Fifth lumbar (seventeenth thoracicolumbar) vertebra with well-developed anapophysis. Hemal arch between second and third caudal vertebrae, with posterior spinous process.
Conditions of soft anatomy (stomach, liver, and male accessory reproductive glands) and glans penis characters are unknown.
Remarks: The complex early taxonomic history of Megalomys was described by Tate (1932a). Although historical museum specimens of M. desmarestii and M. luciae have permitted redescription of the genus and comparison of Megalomys with other extinct rice rats from the Windward and Leeward Islands, the status of other species that have been assigned to the genus requires further clarification through future research. The limited type material of M. audreyae ( Fig. 7B View Figure 7 ) is difficult to evaluate, and the genus-level identity of this species remains unclear. Abundant fossil and zooarchaeological rice rat material is known from the Antigua – Barbuda Bank ( Wing et al., 1968; Steadman et al., 1984a; Jones, 1985; Pregill et al., 1994; MacPhee & Flemming, 1999; Steadman & Hilgartner, 1999), and fossil material from Barbuda has been named ‘ Ekbletomys hypenemus ’ by Ray (1962); however, this taxon remains a nomen nudum because Ray’s work has never been formally published. Although further material apparently referable to M. audreyae has been reported and radiometrically dated by MacPhee & Flemming (1999), the identity of rice rats from the Antigua – Barbuda Bank cannot be determined in the absence of further investigation of this material, which is beyond the scope of the present study.
Two other large-bodied extinct insular cricetid rodents, Megalomys curioi Niethammer, 1964 from the Galápagos Islands and Megalomys curazensis Hooijer, 1959 from Curaçao, were also originally referred to the genus. Megalomys curioi has since been transferred to the new genus Megaoryzomys ( Lenglet & Coppois, 1979) and reinterpreted as a thomasomyine ( Steadman & Ray, 1982). The southern Netherlands Antilles show little biogeographical affinity with the Windward and Leeward Islands, and have close faunal and floral similarities to mainland South America ( Hooijer, 1959, 1966, 1967; Trejo-Torres & Ackerman, 2001; Vázquez-Miranda et al., 2007; Voss & Weksler, 2009), and it has been hypothesized that M. curazensis is derived from a different mainland oryzomyine ancestor ( McFarlane & Lundberg, 2002). A redescription of M. curazensis will form the basis for a future study in this series, and it is not considered further herein.
| NHM |
United Kingdom, London, The Natural History Museum [formerly British Museum (Natural History)] |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SuperFamily |
Muroidea |
|
Family |
|
|
SubFamily |
Sigmodontinae |
|
Tribe |
Oryzomyini |