
Oligoryzomys victus, (THOMAS, 1898)
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2009.00628.x |
|
DOI |
https://doi.org/10.5281/zenodo.13887383 |
|
persistent identifier |
https://treatment.plazi.org/id/7C4E878A-AB78-FF9B-60AB-E79C0BA38FAB |
|
treatment provided by |
Valdenar |
|
scientific name |
Oligoryzomys victus |
| status |
|
OLIGORYZOMYS VICTUS ( THOMAS, 1898) View in CoL
Holotype: NHM 97.12.26.1 (skin and skull).
Examined material: Holotype only.
Diagnosis: Oligoryzomys victus can be diagnosed by its small size, absence of jugal, absence of cranial crests, presence of large stapedial foramen and posterior opening of alisphenoid, absence of squamosal– alisphenoid groove and sphenofrontal foramen, presence of large sphenopalatine vacuities, presence of divided anterocone on M1, presence of two mesofossetti on M2, and presence of ectolophid on m1. This diagnosis, however, is similar to that for South American species of Oligoryzomys , and differentiation between insular and continental species requires further study (see below).
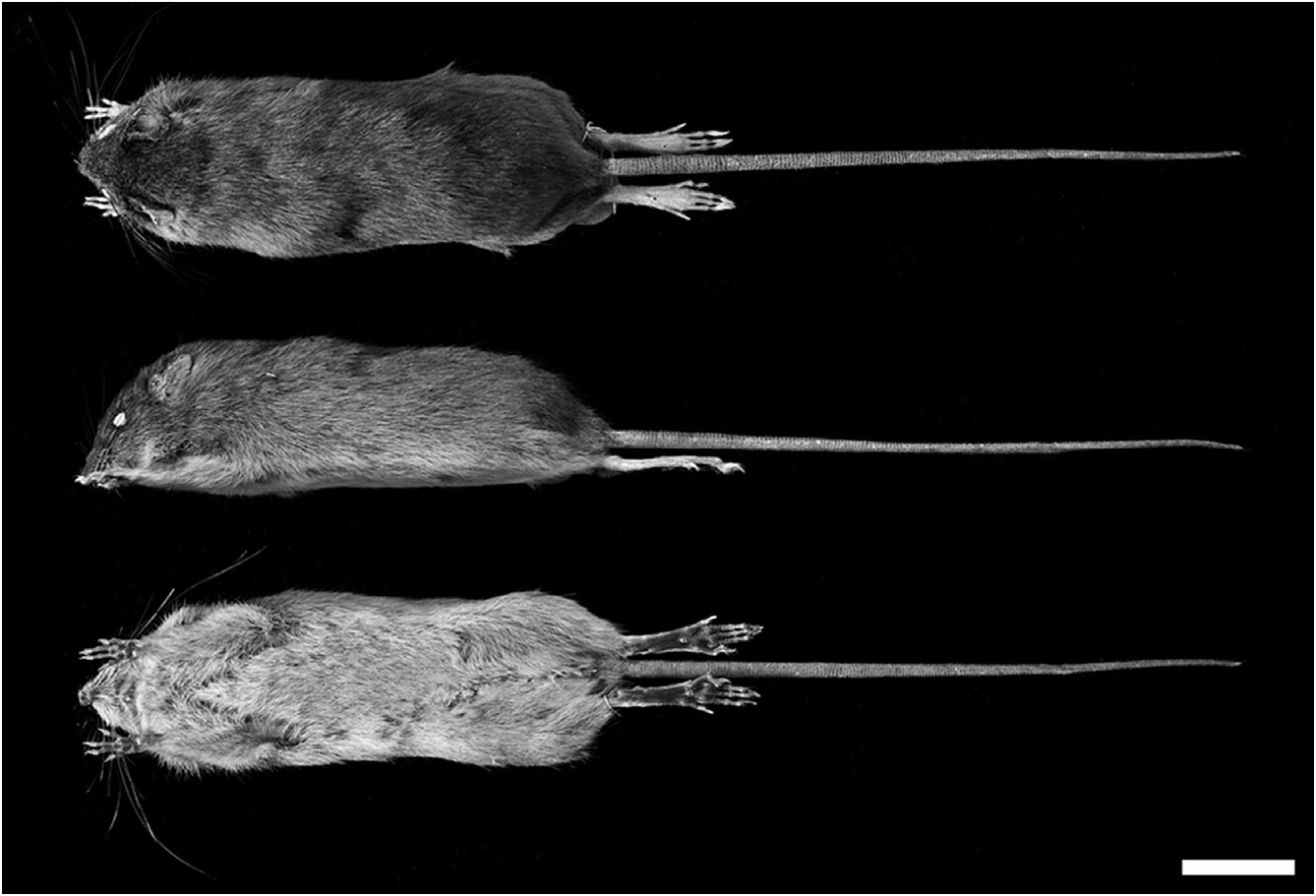
Description: Very small rodent ( Table 1 View Table 1 ) with brown/ ochre dorsal coloration, greyish white ventral pelage, and soft fur without subauricular patches ( Fig. 8 View Figure 8 ). Hindfeet without well-developed natatory fringes and interdigital webs, and with tufts of ungual hairs at bases of claws on dII–dV. Plantar surface densely covered with distinct squamae distal to thenar pad (smooth heel); hypothenar pad large, interdigital pads small and fleshy (pads 1 and 4 displaced proximally relative to 2 and 3). Claw of dI extends to first half of first phalange of dII; claw of dV extends to first interphalangeal joint of dIV. Tail longer than combined length of head and body, sparsely haired, and covered with more or less conspicuous epidermal scales; it lacks a long tuft of terminal hairs, and is weakly bicoloured (dark above, pale below). The mammae are inguinal, abdominal, postaxial, and pectoral, and number eight.
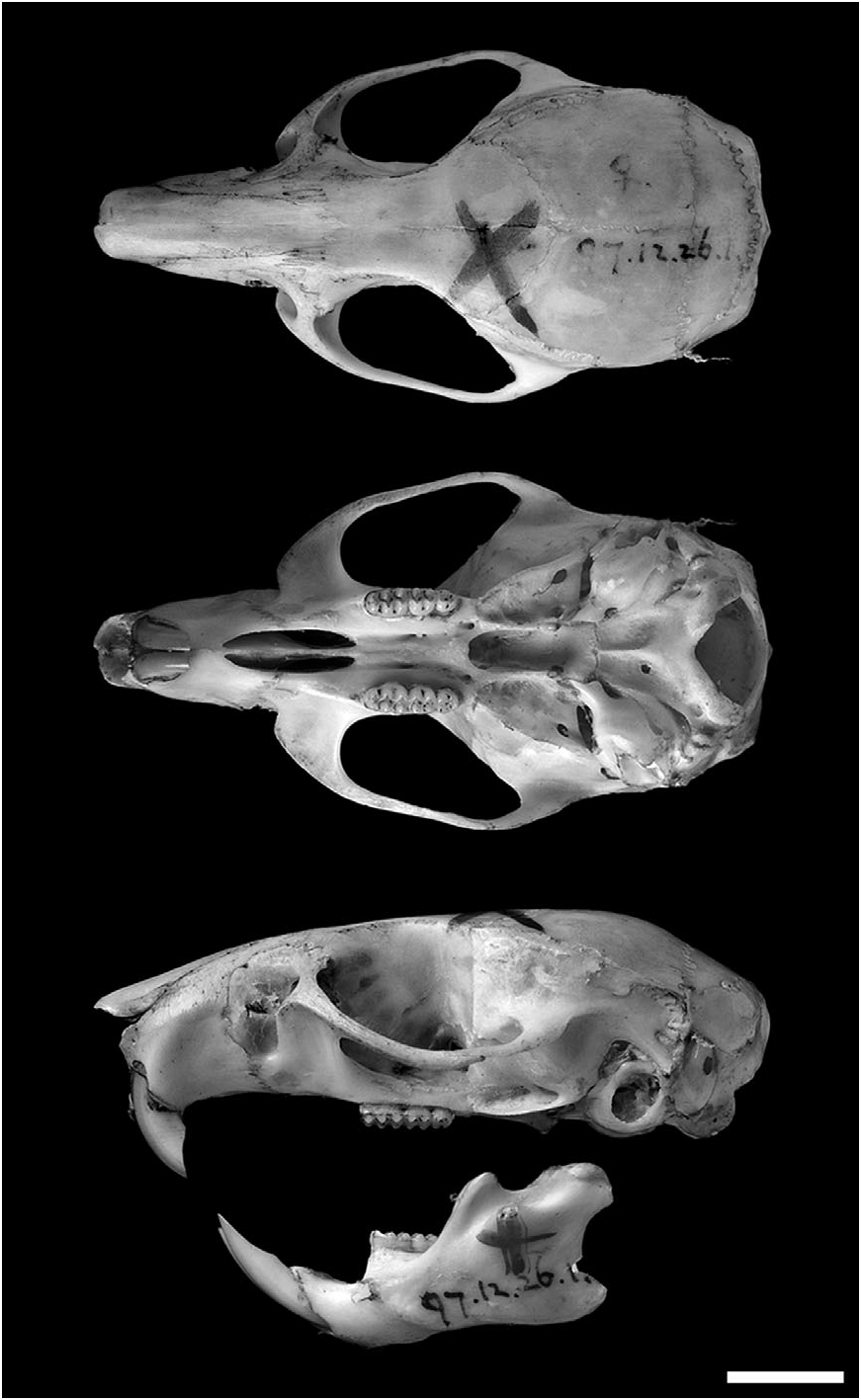
Skull with delicate rostrum flanked by moderate zygomatic notches; interorbital region symmetrically constricted (amphoral shape), with rounded, unbeaded supraorbital margins; braincase without temporal, lambdoidal, and nuchal crests. Zygomatic plate without anterodorsal spinous process, its posterior margin lying anterior to the M1 alveolus. Jugal absent (maxillary and squamosal processes in contact). Nasal bones with blunt posterior margins, not extending posteriorly beyond lacrimals, which are equally sutured to maxillary and frontal bones. Posterior wall of the orbit smooth, frontosquamosal suture colinear with the frontoparietal suture. Parietals without lateral process, interparietal strap shaped. Incisive foramina with posterior border anterior to molar alveoli, and with parallel lateral borders. Bony palate between molar rows smooth and long, mesopterygoid fossa not extending anteriorly between maxillae. Posterolateral palatal pits not recessed in fossae. Bony roof of mesopterygoid fossa perforated by very large sphenopalatine vacuities. The alisphenoid strut is absent (buccinator– masticatory foramen and accessory foramen ovale are confluent), and the alisphenoid canal has a large anterior opening. Stapedial foramen and posterior opening of the alisphenoid canal large, squamosal– alisphenoid groove and sphenofrontal foramen absent (= carotid circulatory pattern 2 of Voss, 1988). Posterior suspensory process of the squamosal absent, postglenoid foramen large and rounded, and subsquamosal fenestra patent but covered by translucid membrane. Periotic exposed posteromedially between the ectotympanic and the basioccipital, extending anteriorly to carotid canal. Mastoid perforated by conspicuous posterodorsal fenestra. In the mandible, the capsular process of the lower incisor alveolus is well developed, and the superior and inferior masseteric ridges converge anteriorly as an open chevron below m1 ( Fig. 9 View Figure 9 ).
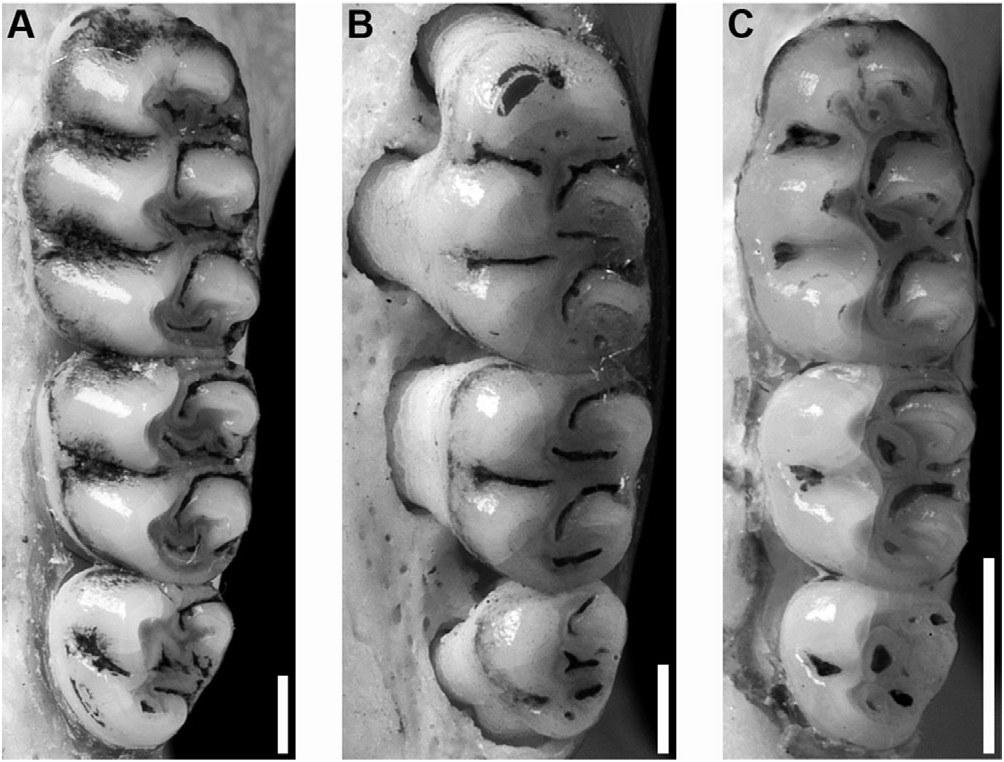
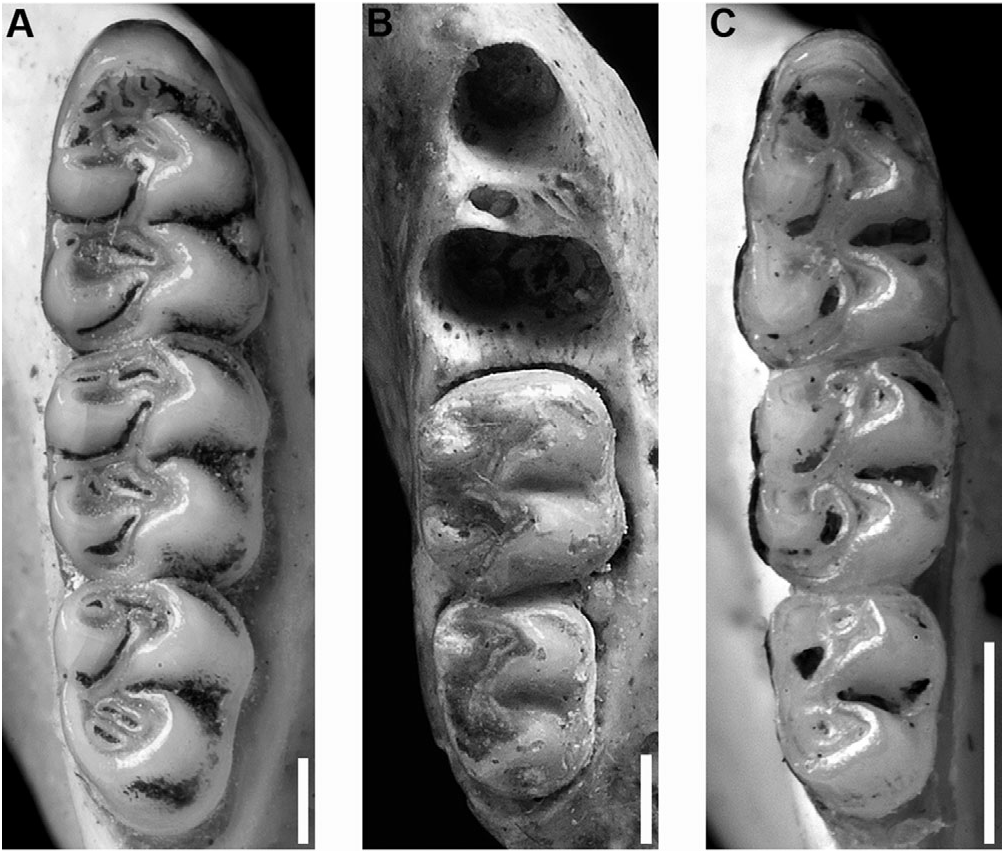
The upper incisors have smoothly rounded enamel bands, and the maxillary tooth rows are parallel. The molars are bunodont, with labial flexi enclosed by a cingulum, and longitudinal flexi not overlapping. M1 anterocone divided into anterolabial and anterolingual conules by deep anteromedian flexus; anteroloph well developed, separated from anterocone by persistent anteroflexus; protostyle absent; mesolophs present on all upper molars; paracone connected by enamel bridge to anterior moiety of protocone; median mure connected to protocone. M2 protoflexus absent; mesoflexus present as two internal fossetti; paracone without an accessory loph. M3 with posteroloph and large hypoflexus ( Fig. 6C View Figure 6 ). m1 anteroconid without anteromedian flexid, but with anteromedian fossettid; anterolabial cingulum present on all lower molars; anterolophid present on m1, but absent on m2 and m3; ectolophid present on m1 and m2; mesolophid distinct on unworn m1 and m2; posteroflexid present on m3 ( Fig. 7C View Figure 7 ).
Remarks: This species shares several characters with the other species of the genus Oligoryzomys , such as: small size; absence of jugal; smooth and amphoral supraorbital margins; absence of cranial crests; caudad-oriented foramen magnum; divided anterocone on M1; presence of posteroloph on M3; presence of large stapedial foramen and posterior opening of alisphenoid, and absence of squamosal–alisphenoid groove and sphenofrontal foramen (carotid circulation pattern 2 of Voss, 1988); and presence of large sphenopalatine vacuities. The last three characters are among the recovered synapomorphies for Oligoryzomys listed by Weksler (2006), and the present phylogenetic results (see below) provide the first cladistic corroboration of the placement of O. victus within that genus.
Oligoryzomys victus View in CoL consistently differs from other studied species of Oligoryzomys View in CoL in certain characters, such as the presence of two mesofossetti in M2, and presence of ectolophid on m1. However, the alpha taxonomy of the genus Oligoryzomys View in CoL is among the most poorly known within the Sigmodontinae View in CoL . Although a revision of the northern South American and Central American species (probable relatives of O. victus View in CoL ) is beyond the scope of this study, further morphological comparisons of O. victus View in CoL with species of continental Oligoryzomys View in CoL are still necessary to establish its status as a distinct species from mainland taxa. It is interesting to note that O. victus View in CoL has apparently not been recorded from pre-Columbian archaeological sites on St. Vincent, in marked contrast to rice rat taxa on other Lesser Antillean islands. This raises the possibility that the species may represent a relatively recent human-assisted translocation to the Windward Islands from an unknown source population in mainland South America. However, it is important to note that O. victus View in CoL is substantially smaller in body size than other described Lesser Antillean rice rats, and so may not have been perceived as a valuable food item by pre-Columbian Amerindians. In addition, a few undescribed small-bodied rice rat specimens that may represent this species have been reported from Amerindian middens on the neighbouring islands of Carriacou and Grenada ( Lippold, 1991; LeFebvre, 2007). Further investigation of archaeological sites in the southern Windward Islands, and detailed morphological studies of mainland Oligoryzomys species , are required to test these alternative hypotheses regarding the native distribution and status of O. victus View in CoL .
| NHM |
United Kingdom, London, The Natural History Museum [formerly British Museum (Natural History)] |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SuperFamily |
Muroidea |
|
Family |
|
|
SubFamily |
Sigmodontinae |
|
Tribe |
Oryzomyini |
|
Genus |