
Chauhanellus hamatopeduncularoideum, Domingues, Marcus V., Soares, Geusivam B. & Watanabe, Alana, 2016
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4127.2.4 |
|
publication LSID |
lsid:zoobank.org:pub:D5AEC22D-65C0-49B6-92C5-B8D1BCBBF91F |
|
DOI |
https://doi.org/10.5281/zenodo.5699400 |
|
persistent identifier |
https://treatment.plazi.org/id/7A05A870-3750-FFD7-E3B5-FB70FD82196A |
|
treatment provided by |
Plazi |
|
scientific name |
Chauhanellus hamatopeduncularoideum |
| status |
sp. nov. |
Chauhanellus hamatopeduncularoideum n. sp.
( Figs. 1–7 View FIGURES 1 – 7 )
Type-host: Amphiarius rugispinis (Valenciennes)
Site of infection: Secondary lamellae of the gills.
Type locality: Fishing village of Ajuruteua, Municipality of Bragança, State of Pará, Brazil (0°49’31”N, 46°36'29"W).
Other records: Amphiarius rugispinis, Bombom Village, Municipality of Viseu, State of Pará, Brazil (01 ° 11’48’’N, 46°08’24’’W); A. rugispinis, Fish market, Municipality of Bragança, State of Pará, Brazil. Sciades couma (Valenciennes) , Fish market, Municipality of Bragança, State of Pará, Brazil.
Prevalence of infection: 23 of 29 Amphiarius rugispinis (79%); 13 of 14 Sciades couma (92%).
Specimens deposited: Holotype, CHIOC 38240a; 16 paratypes, CHIOC 38240b–h, INPA 666, MPEG 044– 047; 35 vouchers, CHIOC 38241a–f, 38242, 38243a–c, 38244a–b, 38245a–b, 38246a–c, INPA 667–669, MPEG 048–056.
Etymology: The specific name refers to the superficial resemblance of the species of Hamatopeduncularia , mainly by possessing hooks located on haptoral digits and absence of expanded deep roots on the ventral anchors.
Comparative measurements: Table 2 View TABLE 2 .
Description: Based on 25 adult specimens; 10 mounted in Gomori’s trichrome, 15 mounted in Hoyer’ medium. Body fusiform ( Fig. 1 View FIGURES 1 – 7 ), total length excluding haptor 754 (710–810; n=5) long, 121 (90–160; n=7) wide at level of germarium. Tegument smooth with a tapered cephalic margin with moderately developed terminal lobes. Three bilateral pairs of head organs with rod-shaped secretion; unicellular cephalic glands lying lateral to pharynx. Equidistant eyes present (4); accessory granules absent. Mouth subterminal, midventral; pharynx ovate, 84 (64– 110; n=5) long, 81 (73–88; n=5) wide; oesophagus, intestinal caeca obscured by vitellaria. Genital pore opening midventral, anterior to copulatory complex; muscular genital atrium. Testis post-germarial, dorsal to germarium, bacilliform, 143 (n=1) long, 71(n=1) wide. Vas deferens apparently looping left of intestinal cecum; seminal vesicle a dilatation of vas deferens, sigmoid. Prostatic reservoir not observed. Copulatory complex comprising male copulatory organ, accessory piece; male copulatory organ, coiled sclerotized tube 206 (185–227; n=11) long, with two clockwise rings, distal aperture acute; base with sclerotized margin. Accessory piece sclerotized, nonarticulated with male copulatory organ comprising sheath along last ring and medial portion of male copulatory organ; distal portion of accessory piece bifurcated, which serves as a guide to male copulatory organ ( Fig. 2 View FIGURES 1 – 7 ). Germarium 123 (119–127; n=3) long, 57 (50–63; n=3) wide, saccate. Vagina comprises vaginal aperture with dextroventral opening, submarginal: vaginal vestibule with soft tissue; vaginal canal elongate, sclerotized, cupshaped. Seminal receptacle, ootype, Mehlis’ glands not observed. Vitellaria dense throughout trunk except in region of reproductive organs. Eggs not observed. Peduncle elongate. Haptor digitate, 170 (n=2) long, 90 (80–100; n=2) wide. Anchors dissimilar. Ventral anchor ( Fig. 7 View FIGURES 1 – 7 ), base 10 (8–13; n=10) long, with divergent roots; superficial root 33 (31–37; n=12) long, deep root 28 (24–30; n=12) long, not expanded; with knob shaped expansion for articulation with ventral bar; slightly curved shaft, elongate point extending well past level of tip of superficial root. Dorsal anchor ( Fig. 6 View FIGURES 1 – 7 ) 41 (38–43; n=6) long, base 14 (11–16; n=7) long, robust, with inconspicuous roots, slightly expanded outer base, slightly curved shaft, elongated point extending well past level of tip of inner base; anchor spine blunt. Ventral bar ( Fig. 4 View FIGURES 1 – 7 ) 6 (4–10; n=9) long, 48(38–52; n=7) wide, dumbbell shaped, with protuberances at each end for articulation with ventral anchor. Dorsal bar ( Fig. 5 View FIGURES 1 – 7 ) 7 (5–8; n=8) long, 50 (43–65; n=7) wide, Vshaped, with bifurcation on both ends, elongated posteromedial process. Hooks similar in shape ( Fig. 3 View FIGURES 1 – 7 ), 14 (11– 16; n = 12) long, shank without inflation, depressed thumb, lightly curved long shaft, and delicate point. Filamentous hook loop not observed.
Remarks: Chauhanellus hamatopeduncularoideum n. sp. is characterized by having a combination of Hamatopeduncularia -like features (i.e., haptoral digits, absence of expanded deep roots on the ventral anchors) and Chauhanellus -like features (i.e., spine on dorsal anchor, dumbbell-shaped ventral bar, dorsal bar with posteromedial process). Lim (1996) reported eight species of monogenoids (4 of Hamatopeduncularia and 4 of Chauhanellus ) that possess a combination of features of both genera, and although there are strictly no unique characteristics (synapomorphies), each species could be assigned to either of the genera based on those combinations (see Lim 1994, 1996). Although the presence of haptoral digits is a diagnostic feature for Hamatopeduncularia ( Yamaguti 1953, 1963; Lim 1996), it is also reported for some species of Chauhanellus : C. australis (Young, 1967) , C. pedunculatus Paperna, 1977 , C. digitalis Lim, 1994 , and the two new species described herein (i.e., C. hamatopeduncularoideum n. sp. and C. susamlimae n. sp.). Kearn & Whittington (1994) considered that the presence of extensible haptoral digits observed in Hamatopeduncularia might be a valid distinction between this genus and Chauhanellus . However, Lim (1996) suggests that haptoral digits should not be considered as a distinguishing feature between Hamatopeduncularia and Chauhanellus . The presence of haptoral digits is also reported for species of Trinigyrus Hanek, Molnar & Fernando 1974 from loricariids ( Siluriformes , Loricariidae ) from Central America and South America. Kritsky et al. (1986) suggested that Hamatopeduncularia and Trinigyrus are closed related mainly by sharing some haptoral elements (i.e., haptoral digits, posteromedial projections in the bar and conspicuous glandular reservoirs), and by parasitizing Siluriformes fishes. However, only a cladistic analysis will indicate the real relationship between these genera with other dactylogyrids, and whether those shared characters are phylogenetically informative. Other important diagnostic characteristic used to discriminate both genera is the presence of expanded deep roots on the ventral anchors, as well as, the presence of ventral bar with bifurcated ends (dumbbell shaped) which occurs in all species of Chauhanellus ( Bychowsky & Nagibina 1969; Lim 1994). Kearn & Whittington (1994) considered the flange on the shaft of the ventral anchor of Hamatopeduncularia pearsoni Kearn & Whittington, 1994 similar to the expanded deep roots on the ventral anchors found on species of Chauhanellus . However, these authors considered that this species belongs to Hamatopeduncularia because it possesses, among other features, extensible haptoral digits. With the exception of Chauhanellus intermedius Lim, 1994 and C. aspinous Lim, 1994 , the presence of a spine on the dorsal anchor is the common characteristic for species of Chauhanellus . We decided to accommodate the new species in Chauhanellus as C. hamatopeduncularoideum n. sp. mainly because it possesses a spine on the inner base of the dorsal anchor, as well as a dumbbell-shaped ventral bar and dorsal bar with spines at both ends and a posteromedial process.
Type-host: Sciades proops (Valenciennes) .
Site of infection: Secondary lamellae of the gills.
Type locality: Fishing village of Ajuruteua, Municipality of Bragança, State of Pará, Brazil (0°49’31”N, 46°36'29"W).
Prevalence of infection: 11 of 11 (100%).
Specimens deposited: Holotype, CHIOC 38247; 13 paratypes, CHIOC 38248a–b, 38249a–b, 38250a–b, INPA 678, MPEG 057–058.
Etymology: The specific name is from Greek (hypen = moustache; kleithron = bar) and refers to the shape of the posterior protuberances of the dorsal bar.
Description: Based on 12 adult specimens; 3 mounted in Gomori’s trichrome, 9 mounted in Hoyer’ medium. Body fusiform ( Fig. 8 View FIGURES 8 – 14 ), total length excluding haptor 339 (250–435; n=4) long, 63 (30–95; n=4) wide at level of germarium. Tegument smooth. Cephalic margin tapered; poorly developed terminal lobes; three bilateral pairs of head organs with rod-shaped secretion; cephalic glands unicellular, posterolateral to pharynx. Eyes 4, posterior pair larger than anterior pair; accessory granules absent or few in cephalic area, spherical. Mouth subterminal, midventral; pharynx subovate, 37(35–40; n=4) long, 32 (27–38; n=4) wide; oesophagus short, intestinal caeca two, non-confluent posteriorly, lacking diverticula. Genital pore opening mid-ventral; genital atrium muscular. Gonads overlapping; testis dorsal to germarium. Testis elongate, 43(n=1) long, 30 (n=1) wide. Vas deferens looping left intestinal cecum; seminal vesicle a dilatation of vas deferens, sigmoid. One prostatic reservoir, pyriform, lying ventral lateral to copulatory complex. Copulatory complex comprising male copulatory organ, accessory piece; male copulatory organ, sclerotized, a coiled tube with 2 counterclockwise rings, base with an expanded sclerotized cap, 188 (148–199; n=7) long, distal aperture acute. Accessory piece sclerotized, non-articulated with male copulatory organ comprising an elongated sheath with bifurcated distal portion ( Fig. 9 View FIGURES 8 – 14 ). Germarium 64 (63–65; n=2) long, 25 (24–27; n=2) wide, elongated. Vagina comprising vaginal aperture with dextroventral opening, marginal; vaginal vestibule, vaginal canal; vaginal canal sclerotized, elongated, sigmoid, with distal portion expanded before entering into seminal receptacle. Seminal receptacle ovate; Mehlis’ glands, ootype not observed. Vitellaria dense throughout trunk, except in region of reproductive organs. Eggs not observed. Peduncle elongate. Haptor subhexagonal, 19 (14–30; n=4) long, 23 (14–40; n=4) wide. Anchors dissimilar. Ventral anchor ( Fig. 14 View FIGURES 8 – 14 ), base 33 (30–35; n=8) long, with divergent roots, truncate superficial root, 35 (33–40; n=8) long, expanded deep root, 46 (43–49; n=8) long; short shaft, elongated point; point extending beyond of tip of superficial root. Dorsal anchor ( Fig. 13 View FIGURES 8 – 14 ) 46 (45–48; n=8) long, base 17 (16–17; n=8) long, robust, with inconspicuous roots, evenly curved shaft, point; point extending well past level of tip of inner base; anchor spine blunt. Ventral bar ( Fig. 10 View FIGURES 8 – 14 ) 10 (8–14; n=7) long, 65 (57–75; n=7) wide, curved in posterior direction with longitudinal groove; protuberances at each end for articulation with ventral anchor, posterior protuberance elongated with hook shaped termination at distal portion. Dorsal bar ( Fig. 11 View FIGURES 8 – 14 ) 8 (6–10; n=6) long, 42 (38–48; n=8) wide, V-shaped, with rounded ends; donkey-ear like protuberance on both ends; short posteromedial process with medial constriction (moustache-like). Hooks similar in shape ( Fig. 12 View FIGURES 8 – 14 ), 12(10–13; n = 8) long, shank without inflation, depressed thumb, lightly curved long shaft, delicate point, filamentous hook loop about half of shank length.
Remarks: Chauhanellus hypenocleithrum n. sp. is morphologically closer to C. hamatopeduculoideum n. sp. due to the presence of coiled male copulatory organ and also by having a sclerotized vaginal vestibule and canal; hooks with shank without inflation. It differs from C. hamatopeduculoideum n. sp. by possessing a haptor without digits, ventral anchor with expanded deep root, and ventral bar with an elongate posterior protuberance. The new species also differs from all other congeneric species by possessing a dorsal bar with a short posteromedial process with medial constriction (moustache-like).
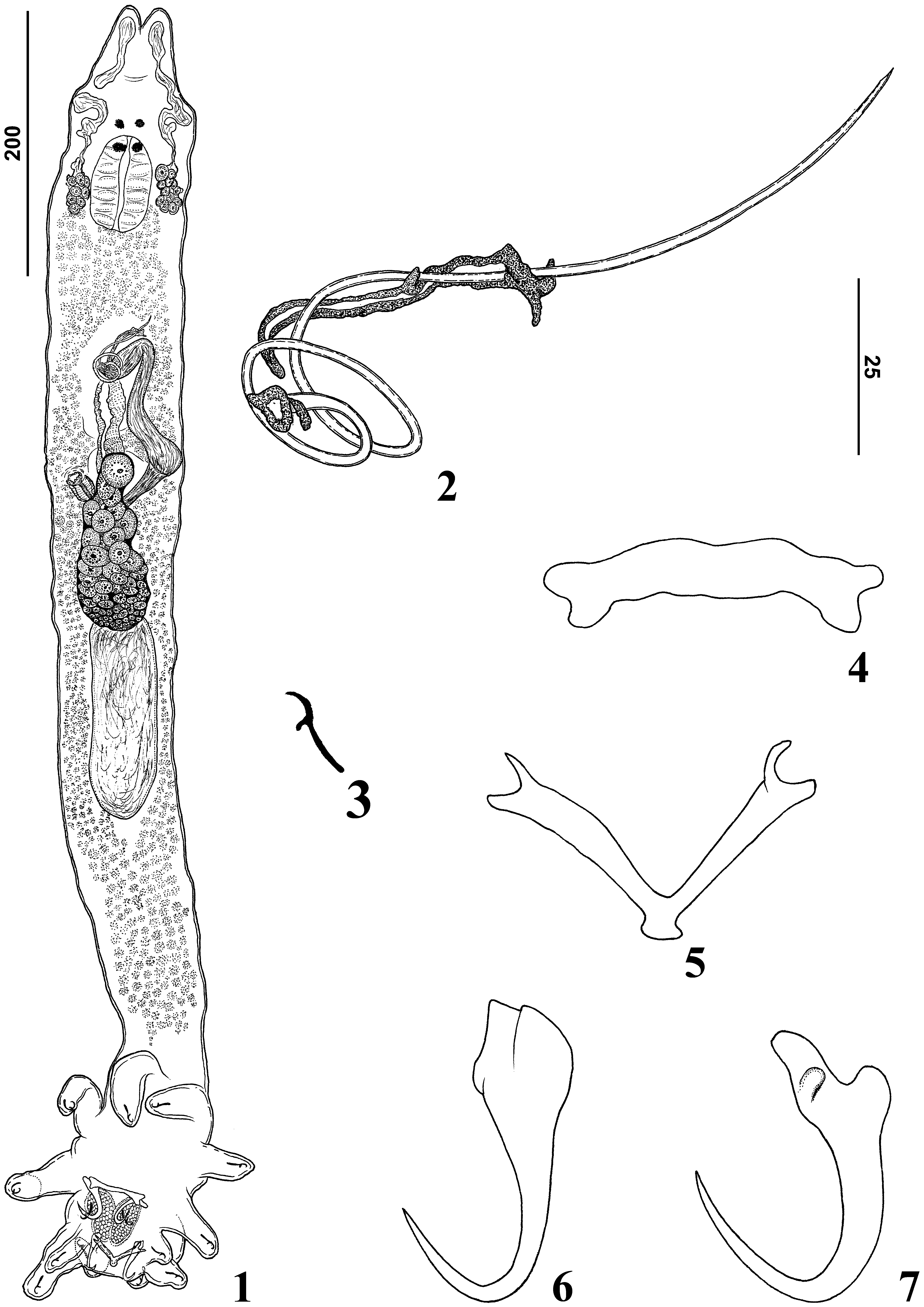

TABLE 2. Comparative measurements (in µm) of specimens of Chauhanellus hamatopeduncularoideum n. sp. from the gills of Amphiarius rugispinis and Sciades couma. MCO = male copulatory organ.
| A. rugispinis | N | S. couma | N |
|---|---|---|---|
| Body | |||
| Length 754 (710–810) | 5 | 527(420–600) | 3 |
| Width 121 (90–160) | 7 | 97(60–120) | 3 |
| Haptor | |||
| Length 170 (170–170) | 2 | 145(140–150) | 2 |
| Width 90 (80–100) | 2 | 155(140–170) | 2 |
| Pharynx | |||
| Length 84 (64–110) | 5 | 58(57–58) | 2 |
| Width 81 (73–88) | 5 | 51(50–53) | 2 |
| MCO length 206 (185–227) | 11 | 182(163–198) | 3 |
| MCO ring 20 (18–23) | 11 | 26(25–27) | 3 |
| Ventral Bar | |||
| Length 6 (4–10) | 9 | 7(6–8) | 3 |
| Width 48 (38–52) | 7 | 48(48–49) | 2 |
| Dorsal Bar | |||
| Length 7 (5–8) | 8 | 8(8–10) | 4 |
| Width 50 (43–65) | 7 | 51(48–54) | 2 |
| Ventral Anchor | |||
| Inner 28 (24–30) | 12 | 31 (29–32) | 4 |
| Outer 33 (31–37) | 12 | 33 (32–33) | 4 |
| Base 10 (8–13) | 10 | 13 (13–14) | 4 |
| Dorsal Anchor | |||
| Inner 40 (38–43) | 5 | 41 (40–41) | 5 |
| Outer 41 (38–43) | 6 | 43 (41–43) | 5 |
| Base 14 (11–16) | 7 | 14 (13–14) | 5 |
| Hooks 14 (11–16) | 12 | 15 | 2 |
| Germarium | |||
| Length 123 (119–127) | 3 | 74 (71–76) | 2 |
| Width 57 (49–63) | 3 | 40 (32–48) | 2 |
| Testis | |||
| Length 123 (119–127) | 3 | 74 (71–76) | 2 |
| Width 57 (49–63) | 3 | 40(32–48) | 2 |
| Chauhanellus hypenocleithrum n. sp. (Figs. 8–14) |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |