
Tytthoscincus batupanggah, Karin, Benjamin R., Das, Indraneil & Bauer, Aaron M., 2016
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4093.3.7 |
|
publication LSID |
lsid:zoobank.org:pub:A7080821-87BE-48A3-96A4-D7C9F86F1766 |
|
DOI |
https://doi.org/10.5281/zenodo.6085912 |
|
persistent identifier |
https://treatment.plazi.org/id/79268628-9E0E-5321-FF26-F8ADCB1F9C16 |
|
treatment provided by |
Plazi |
|
scientific name |
Tytthoscincus batupanggah |
| status |
sp. nov. |
Tytthoscincus batupanggah sp. nov.
( Figs. 1–2 View FIGURE 1 View FIGURE 2 )
Holotype. A specimen (CAS 259189) from Batu Panggah, 1.120666º N 110.226922º E (WGS84), 1,050 m elevation, on the north face of Gunung Penrissen, Kuching Division, Sarawak, Malaysia collected 4 July 2014 at 20:00 h by Benjamin R. Karin, Pui Yong Min, Samuel Shonleben, and Adi Shabrani.
Paratype. One additional specimen (ZRC 2.7123) was collected at the same locality at the same time as the holotype.
Diagnosis. Tytthoscincus batupanggah sp. nov. can be identified by the following combination of characters: (1) body size small (SVL = 26.8–33.2 mm); (2) MBSR = 30–31; (3) PVSR = 62–66; (4) VSR = 55–61; (5) four enlarged supraoculars; (6) parietals in contact with the last supraocular; (7) prefrontals in contact with one another; (8) two loreals; (9) six supralabials; (10), five infralabials; (11) nine SDL on Toe IV; (12) smooth texture of the subdigital lamellae; (13) enlarged precloacal scales; (14) a dark dorsolateral stripe; (15) a light postorbital stripe; and (16) a light dorsolateral stripe.
Description of Holotype. All measurements in mm. SVL = 33.2; TL = 28.4 (tail incomplete, used for additional genetic material); AGD = 17.5; HL = 6.9; HW = 4.5; SFL = 12.8; sex unknown; rostral wider than long, in broad contact with frontonasal; frontonasal wider than long; prefrontals large, in broad contact, left prefrontal overlapping right; frontal elongate, triangular in shape, in contact with the first two supraoculars and one preocular; frontoparietals in broad contact, contacting second, third, and fourth supraoculars, parietals, interparietal, and frontal; right frontoparietal overlaps left; interparietal diamond-shaped, large, elongate posteriorly, with parietal eyespot in posterior projection; parietals large, in contact posterior to interparietal, in slight contact with posterior portion of fourth interparietal; left parietal overlaps right; enlarged nuchal scales absent; nasals small, widely separated, completely enclosing the nares, and contacting the rostral anteriorly, frontonasal dorsally, first loreal posteriorly, and first supralabial ventrally; six supralabials; supranasals absent; two similarly sized loreals taller than wide; three similarly sized preoculars in contact with posterior margin of second loreal; nine supraciliaries; posterior supraciliary elongate, projecting medially; temporals not distinctly enlarged in comparison to lateral body scales; lower eyelid transparent, scaly, without central window; mental twice as wide as long; single postmental, contacting first infralabial on each side; two enlarged pairs of genials following postmental, anterior pair in contact medially, posterior pair widely separated by a single ventral scale; genials contacting first, second, and third infralabials; five infralabials; external ear opening two-thirds the diameter of the eye, circular, with an auricular component projecting from the posterior margin; tympanum sunk.
Body scales smooth, hexagonal; ventral scales not enlarged; 30 longitudinal scale rows around the midbody; 66 paravertebral scale rows; 61 ventral scale rows; two enlarged, semi-transparent precloacal scales, left overlapping right, and both overlapping outer scales; tail robust, cylindrical, incomplete; anterior subcaudals equal in size to dorsal caudals; posterior subcaudals enlarged.
Adpressed limbs widely separated; lamellae texture smooth; manus subdigital lamellae (L/R) in parentheses: I(2/2), II(4/4), III(5/5), IV((incomplete)/6), V(4/4); pes subdigital lamellae (L/R) in parentheses: I(5/4), II((incomplete)/5), III(8/8), IV(9/9), V(6/6);
Color in alcohol. Dorsal and lateral surfaces of holotype mostly brown with tiny cream-yellow spots dotting the entirety of the body; dorsolateral stripe extending from the orbit to midway along the body, cream in color, underlined by a solid-colored, dark-brown lateral stripe, both stripes fading away posterior to the forelimbs; a second lateral stripe extends from the orbit to the ear opening, cream in color; ventral surface of body and ventral surface of base of tail cream in color, posterior ventral surface of tail punctuated with tiny brown dots of pigmentation.
Variation. The paratype (ZRC 2.7123) differs from the holotype in SVL = 26.8, TL = 11.0 (incomplete), AGD = 13.7, HL = 6.1, HW = 3.9, SFL = 10.2, MBSR = 31, and VSR = 55 but is identical to the holotype in all other scale characters reported (four supraoculars, parietals contact supraocular, nine supraciliaries, prefrontals in contact, two loreals, six supralabials, five infralabials, SDL on toe IV = 9, precloacal scales enlarged). The coloration of the paratype is extremely similar to the holotype, but differs in the thickness of the cream-colored dorsolateral stripe, which in the paratype is more distinct nearer to the orbit than the stripe of the holotype. In life, the coloration of both the holotype and paratype is more orange, with a highlight of orange near the axilla ( Fig. 1 View FIGURE 1 ).
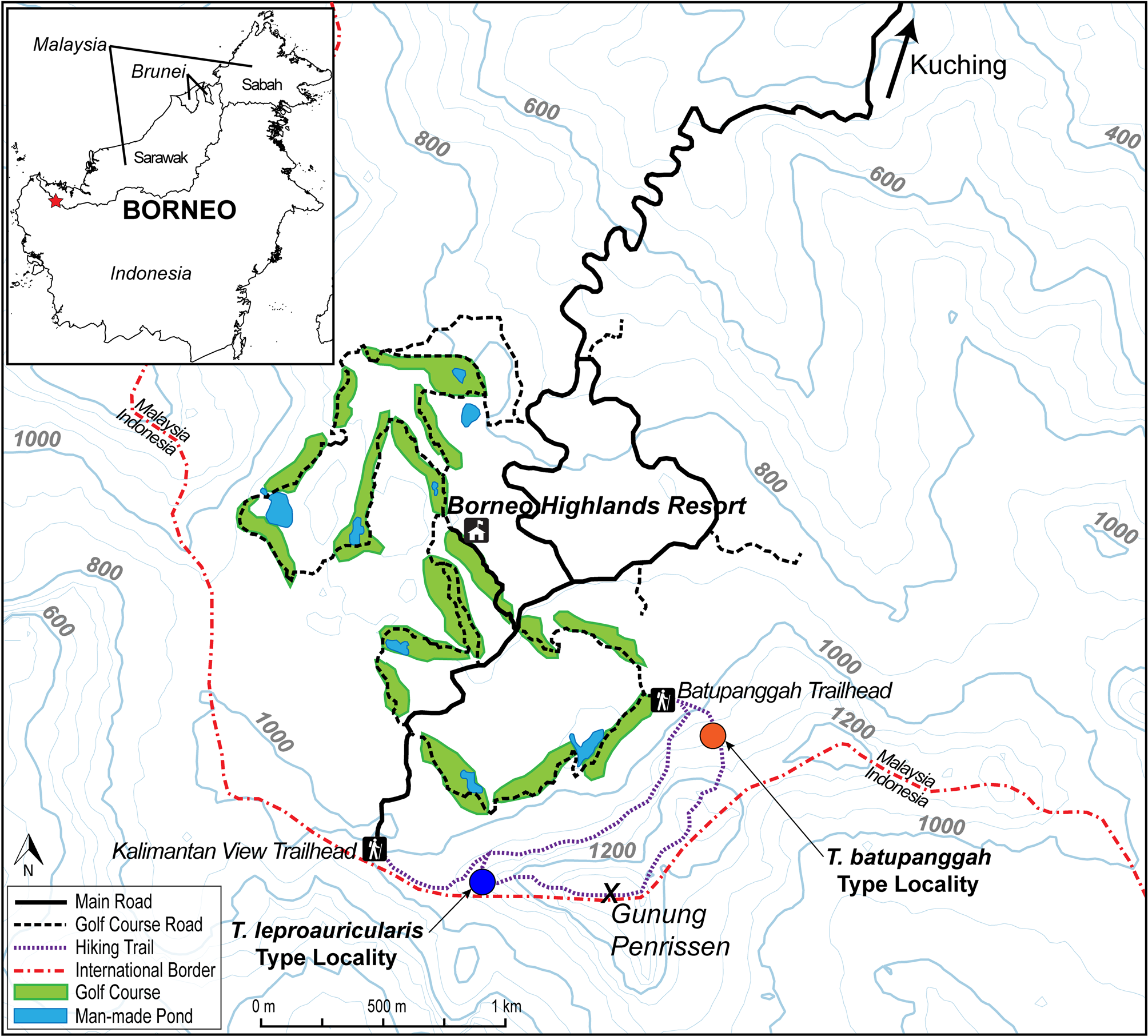
Distribution. Tytthoscincus batupanggah is known only from the type locality from Batu Panggah on the North side of Gunung Penrissen, Kuching Division, Sarawak, Malaysia ( Fig. 6 View FIGURE 6 ). We expect that it exists in much of the high-elevation forest on the mountain, though it is possible that it is replaced in other parts of its range by Tytthoscincus leproauricularis sp. nov., another similarly-sized, leaf-litter species. Unfortunately, much of the original forest on Gunung Penrissen has been clear-cut to make way for a golf course, substantially limiting the suitable habitat for this species (see Fig. 6 View FIGURE 6 ).
Natural history. The two known specimens of Tytthoscincus batupanggah sp. nov. were collected at the same time, date, and location from leaf-litter surrounding Batu Panggah on Gunung Penrissen. The area is primary, highland, mixed-dipterocarp forest. When collected, the forest understory around Batu Panggah had been recently cleared to improve the view of the large sandstone rock, consequently making it easier for herpetologists to inspect the leaf-litter for lizards. This species appears to be a leaf-litter specialist and was seen running and jumping through this microhabitat (BRK, pers. obs.). The specimens were found after dark, at approximately 20:00 h. It is difficult to determine whether this species was actively foraging during this time (as was the initial speculation by BRK) and therefore semi-nocturnal, or if resting individuals were simply disturbed by nearby human footsteps, though we expect the latter to be true.
Etymology. The species epithet batupanggah is in reference to the type locality at Batu Panggah on Gunung Penrissen. From Malay, Batu Panggah translates to “Stone of the Head House” or “Cursed Stone” to refer to the large sandstone rock that is believed by local peoples to be cursed. The story goes that a young couple from different longhouses fell in love, but when their respective longhouses found out about their romance they sent out the headhunters to hunt them down. As they fled into the jungle it began to pour rain with thunder and lightning, they found a house in the woods, a head house, where human heads were traditionally kept as a symbol of victory, and ran inside to take shelter. However, women are not traditionally allowed in the head house and when they ran inside the house turned to stone, leaving behind a large rock that is now known as Batu Panggah. Suggested common name: Cursed-stone Diminutive Leaf-Litter Skink.
Comparisons. In order to be conservative in differentiating the new species from all others, we compare Tytthoscincus batupanggah sp. nov. to all currently recognized Tytthoscincus and all candidate species of the genus Sphenomorphus that have genetic or phenotypic similarity or which are geographically proximate and have an adult SVL less than or equal to 50 mm (see Introduction; Table 1). Though we cannot be completely sure that other uncollected individuals of T. batupanggah sp. nov. (SVL = 33.2 mm) may have a maximum SVL> 50 mm, since the Holotype lacks an umbilical scar it is likely that it represents an adult size. Therefore, all Sphenomorphus with an adult SVL greater than 50 mm can be differentiated from T. batupanggah sp. nov. (SVL = 26.8–33.2 mm) by body size, however we do not directly list them here. Though we consider Sphenomorphus shelfordi to be too large to be a member of Tytthoscincus (see Introduction), nevertheless we conservatively choose to compare directly with it as it was described from the same type locality as the new species.
MBSR = 30–31 in T. batupanggah sp. nov. distinguishes it from T. leproauricularis sp. nov. (35–37), T. hallieri (33–41), S. langkawiensis (34–37), S. sanana (28), S. buttikoferi (24), S. schlegeli (22), S. apalpebratus (27–28), S. alfredi (26), S. cophias (24), S. courcyanum (24), S. grandisonae (34), S. macullicolus (35–36), S. sheai (20), S. tanahtinggi (40–42), S. tenuiculus (26), S. tetradactylus (20), and S. tridigitus (18). PVSR = 62–66 in T. batupanggah sp. nov. distinguishes it from T. leproauricularis sp. nov. (68–74), S. bukitensis (73–74), S. ishaki (68–73), S. sibuensis (58), S. temengorensis (68–70), S. temmincki (68–80), S. malayanus (76–80), S. senja (72– 73), S. vanhuerni (81), S. cophias (56), S. grandisonae (71), S. macullicolus (79), S. mimicus (61), S. sanctus (71), S. scotophilus (67–74), S. sheai (53), S. tanahtinggi (76–79), S. tenuiculus (57), S. tetradactylus (48), and S. tridigitus (50). VSR = 55–61 in T. batupanggah sp. nov. distinguishes it from T. leproauricularis sp. nov. (80–81), T. hallieri (63–73), S. butleri (62–63), S. langkawiensis (70–72), S. sibuensis (52), S. temengorensis (70,71), S. temmincki (62–72), S. malayanus (74), S. senja (68), S. macullicolus (84), S. sheai (54), and S. tenuiculus (68). The presence of 4 supraoculars distinguishes T. batupanggah sp. nov. from S. textus (5), S. apalpebratus (5), S. macullicolus (7), S. puncticentrallis (5), S. sanctus (5), S. scotophilus (5), and S. tanahtinggi (5). The contact of the parietal with the last supraocular distinguishes T. batupanggah sp. nov. from T. hallieri , and S. tetradactylus . The contact of the prefrontals distinguishes T. batupanggah sp. nov. from all species with prefrontals separated: T. leproauricularis sp. nov., T. aesculeticola , T. biparietalis , T. parvus , S. sanana , S. senja , S. vanhuerni , S. apalpebratus , S. alfredi , S. cophias , S. helenae , S. macullicolus , S. sheai , S. tenuiculus , S. tetradactylus , and S. tridigitus . The presence of a paired anterior loreals distinguishes T. batupanggah sp. nov. from S. sibuensis , S. macullicolus , S. tanahtinggi , S. tetradactylus , and S. tridigitus . The presence of 6 supralabials distinguishes T.
batupanggah sp. nov. from S. sanana (5), S. malayanus (7), S. senja (7), S. cophias (7), S. mimicus (7), S. puncticentrallis (7), S. sanctus (7), S. scotophilus (7), S. tanahtinggi (8–9), S. tenuiculus (7), and S. tonkinensis (7). The presence of 5 infralabials distinguishes T. batupanggah sp. nov. from T. biparietalis (6), S. perhentianensis (6), S. malayanus (7–8), S. senja (6), S. cophias (7), S. macullicolus (6), S. puncticentrallis (7), S. sanctus (6), S. scotophilus (6), S. tanahtinggi (7), S. tenuiculus (6), S. tetradactylus (7), and S. tonkinensis (6). SDL on Toe IV = 9 in T. batupanggah sp. nov. distinguishes it from T. leproauricularis sp. nov. (10), T. atrigularis (10), T. biparietalis (10), T. hallieri (10–15), T. parvus (10), S. bukitensis (12–13), S. butleri (12–13), S. ishaki (11), S. langkawiensis (11,12), S. perhentianensis (10), S. temengorensis (10,11), S. textus (15), S. buttikoferi (21–23), S. malayanus (12– 13), S. necopinatus (11–15), S. schlegeli (10), S. senja (13–17), S. vanhuerni (14–15), S. alfredi (12), S. courcyanum (11), S. grandisonae (12), S. helenae (17), S. macullicolus (18–23), S. mimicus (16), S. modigliani (15), S. puncticentrallis (25), S. sanctus (26–27), S. scotophilus (22–23), S. sheai (6), S. shelfordi (28–29), S. tanahtinggi (16–17), S. tenuiculus (21–24), S. tetradactylus (10), S. tonkinensis (15–19), and S. tridigitus (7–8). The presence of smooth SDL distinguishes T. batupanggah sp. nov. from all species with keeled SDL: S. bukitensis , S. ishaki , S. langkawiensis , S. sibuensis , S. temengorensis , S. malayanus , S. apalpebratus , and S. tanahtinggi . The presence of enlarged precloacal scales distinguishes T. batupanggah sp. nov. from T. aesculeticola , T. biparietalis , T. hallieri , S. bukitensis , S. temmincki , S. apalpebratus , and S. alfredi . The presence of a dark dorsolateral stripe distinguishes T. batupanggah sp. nov. from T. leproauricularis sp. nov., T. aesculeticola , T. parvus , S. perhentianensis , S. sanana , S. senja , S. vanhuerni , S. macullicolus , S. modigliani , and S. sanctus . The presence of a light postorbital stripe distinguishes T. batupanggah sp. nov. from T. leproauricularis sp. nov., T. aesculeticola , T. atrigularis , T. hallieri , T. parvus , S. bukitensis , S. butleri , S. perhentianensis , S. sanana , S. sibuensis , S. temmincki , S. textus , S. necopinatus , S. schlegeli , S. senja , S. vanhuerni , S. apalpebratus , S. alfredi , S. cophias , S. courcyanum , S. grandisonae , S. helenae , S. macullicolus , S. mimicus , S. modigliani , S. puncticentrallis , S. sanctus , S. scotophilus , S. sheai , S. shelfordi , S. tanahtinggi , S. tenuiculus , S. tetradactylus , S. tonkinensis , and S. tridigitus . The presence of a light dorsolateral stripe distinguishes T. batupanggah sp. nov. from T. leproauricularis sp. nov., T. aesculeticola , T. atrigularis , T. biparietalis , T. hallieri , T. parvus , S. bukitensis , S. butleri , S. ishaki , S. perhentianensis , S. temmincki , S. textus , S. buttikoferi , S. malayanus , S. necopinatus , S. senja , S. vanhuerni , S. alfredi , S. cophias , S. macullicolus , S. mimicus , S. modigliani , S. puncticentrallis , S. sanctus , S. scotophilus , S. shelfordi , S. tanahtinggi , S. tenuiculus , S. tonkinensis , and S. tridigitus .


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.