
Tytthoscincus leproauricularis, Karin, Benjamin R., Das, Indraneil & Bauer, Aaron M., 2016
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4093.3.7 |
|
publication LSID |
lsid:zoobank.org:pub:A7080821-87BE-48A3-96A4-D7C9F86F1766 |
|
DOI |
https://doi.org/10.5281/zenodo.6085914 |
|
persistent identifier |
https://treatment.plazi.org/id/79268628-9E0A-532C-FF26-FB66CCEA9F26 |
|
treatment provided by |
Plazi |
|
scientific name |
Tytthoscincus leproauricularis |
| status |
sp. nov. |
Tytthoscincus leproauricularis sp. nov.
( Figs. 3–4 View FIGURE 3 View FIGURE 4 )
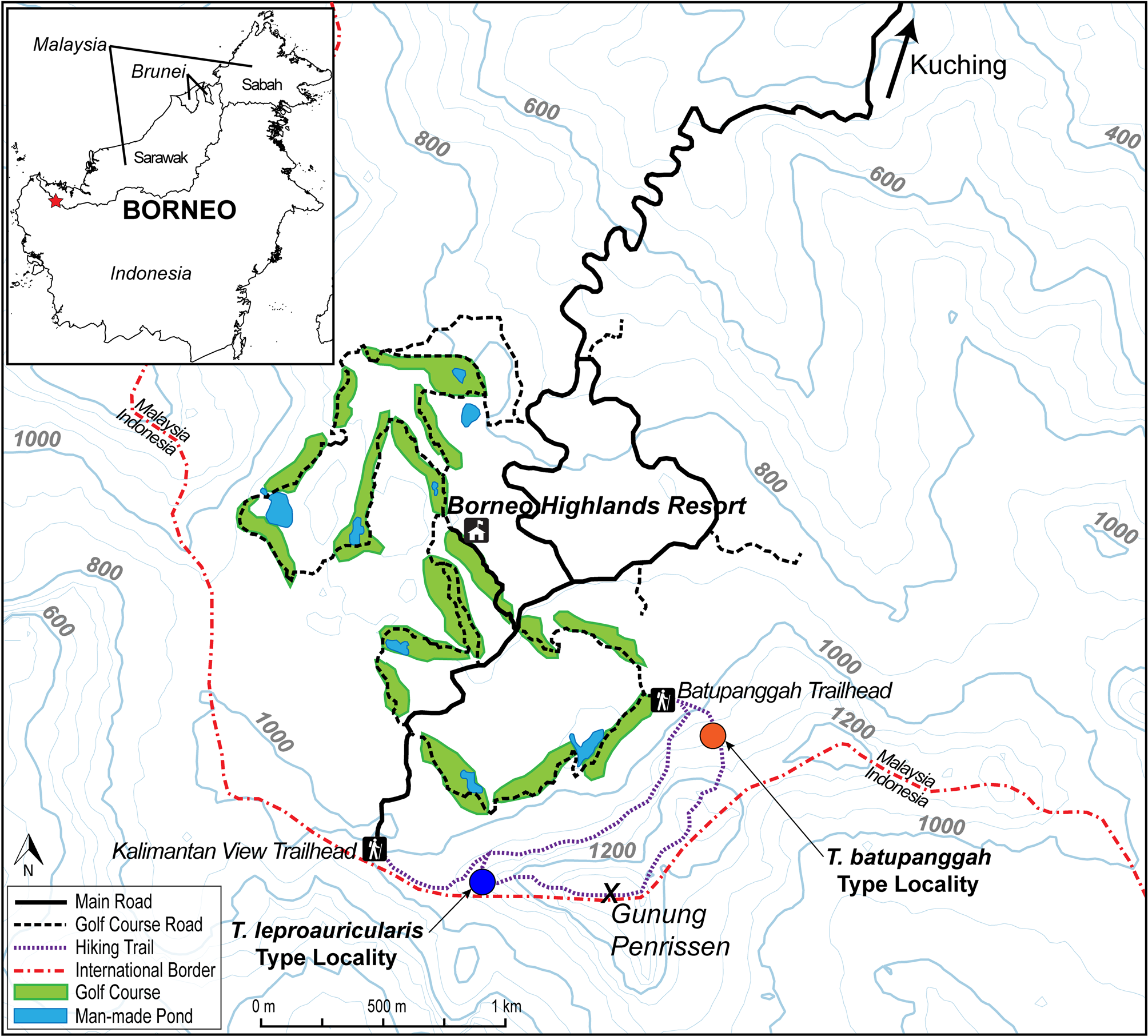
Holotype. A specimen (CAS 259190) from the West ridge of Gunung Penrissen, 1.115302º N 110.218629º E (WGS84), 1,100 m elevation, Kuching Division, Sarawak, Malaysia collected 14 August 2014 at approximately 13:00 h by Benjamin R. Karin, Pui Yong Min, Samuel Shonleben, and Adi Shabrani.
Paratype. A juvenile of the same species (ZRC 2.7122) was collected within 100 m of the holotype on 4 July 2014 by Benjamin R. Karin, Pui Yong Min, Samuel Shonleben, and Adi Shabrani.
Diagnosis. Tytthoscincus leproauricularis sp. nov. can be identified by the following combination of characters: (1) body size small (SVL = 33.5 mm); (2) MBSR = 35–37; (3) PVSR = 68–74; (4) VSR = 80–81; (5) four enlarged supraoculars; (6) parietals in contact with the last supraocular; (7) prefrontals separated; (8) two loreals; (9) six or seven supralabials; (10), five infralabials; (11) ten SDL on Toe IV; (12) smooth texture of the subdigital lamellae; (13) enlarged precloacal scales; (14) lacking a dark dorsolateral stripe; (15) lacking a light postorbital stripe; and (16) lacking a light dorsolateral stripe.
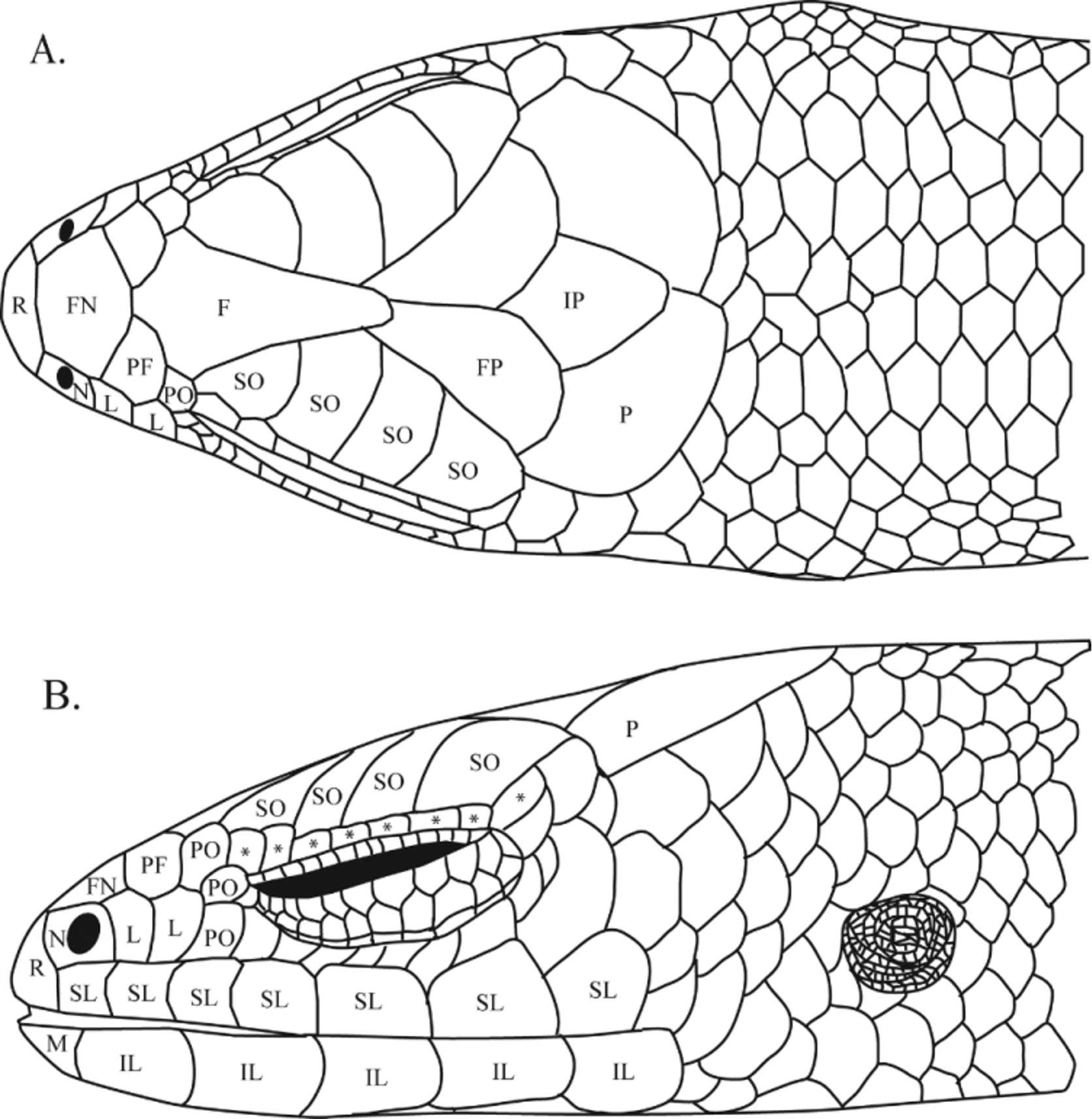
Description of Holotype. SVL = 33.5; TL = 5.5 (tail incomplete, used for additional genetic material); AGD = 19.1; HL = 7.3; HW = 4.4; SFL = 12.5; sex unknown; rostral wider than long, in broad contact with frontonasal; frontonasal wider than long; prefrontals large, widely separated; frontal elongate, triangular in shape, in contact with the first two supraoculars and one preocular; frontoparietals in broad contact, contacting second, third, and fourth supraoculars, parietals, interparietal, and frontal; right frontoparietal overlaps left; interparietal diamondshaped, large, posteriorly elongate, parietal eyespot absent; parietals large, in contact posterior to interparietal, in slight contact with posterior portion of fourth supraocular; left parietal overlaps right; enlarged nuchal scales absent; nasals small, widely separated, completely enclosing the nares, and contacting the rostral anteriorly, frontonasal dorsally, first loreal posteriorly, and first supralabial ventrally; seven supralabials; supranasals absent; two similarly sized loreals taller than wide; three similarly sized preoculars in contact with posterior margin of second loreal; eight supraciliaries; posterior supraciliary elongate and projecting medially; temporals enlarged relative to lateral body scales; lower eyelid scaly, without central window; mental twice as wide as long, in contact with postmental; single postmental, contacting first infralabial on each side; two enlarged pairs of genials following postmental, anterior pair in contact medially, posterior pair widely separated by a single ventral scale; genials contacting first, second, and third infralabials; five infralabials; ear opening scaly, circular, superficial, without visible tympanum, half the diameter of the eye.
Body scales smooth, hexagonal; ventral scales not enlarged relative to dorsal scales; 35 scale rows around the midbody; 74 paravertebral scale rows; 70 ventral scale rows; three enlarged, semi-transparent precloacal scales, center overlapping outer two; tail robust, cylindrical, incomplete.
Adpressed limbs widely separated; lamellae texture smooth; toe IV slightly longer than toe III; manus subdigital lamellae (L/R) in parentheses: I(2/2), II(3/3), III(4/4), IV(4/,(incomplete)), V(3/3); pes subdigital lamellae (L/R) in parentheses: I(4/4), II(6/6), III(9/9), IV(10/10), V(5/5).
Color in alcohol. Dorsal and lateral surfaces mostly brown with tiny beige spots dotting the entirety of the body; no dorsolateral stripe; ventral surface of body cream in color, chin and neck with small brown flecks of pigmentation.
Variation. The paratype (ZRC 2.7122) differs from the holotype in SVL = 18.2, TL = 9.0 (incomplete), AGD = 8.3, HL = 4.4, HW = 2.9, SFL = 7.8, MBSR =37, PVRS = 68, VSR = 80, six supralabials but is identical to the holotype in all other scale characters reported (four supraoculars, parietals contact supraocular, eight supraciliaries, prefrontals separated, two loreals, five infralabials, SDL on toe IV = 10, precloacal scales enlarged). Coloration of the juvenile paratype extremely similar to the holotype, though the paratype has slightly more intense beige spotting along the body. In life, the coloration of the holotype is nearly identical to the preserved specimen (see Fig. 3 View FIGURE 3 ).
Distribution. Tytthoscincus leproauricularis sp. nov. is only known from the type locality on the Western side of Gunung Penrissen, Kuching Division, Sarawak, Malaysia ( Fig. 6 View FIGURE 6 ). We expect that it exists over much of the high-elevation forests on these mountains, though it is possible that it is replaced in other parts of its range by Tytthoscincus batupanggah sp. nov., another similarly-sized, leaf-litter species. Unfortunately, much of the original forest on Gunung Penrissen has been clearcut to make way for a golf course, substantially limiting the suitable habitat for this species (see Fig. 6 View FIGURE 6 ).
Natural history. The holotype and paratype of Tytthoscincus leproauricularis sp. nov. were collected within 100 m of each other while walking on the summit trail up to the ridge on the west face of Gunung Penrissen. The collection location is composed of primary, highland, mixed-dipterocarp forest. Only 100–150 m elevation above the collection location the forest transitions to a localized area of tropical cloud forest with substantially shorter trees. The paratype was collected on 4 July 2014 and the holotype over a month later, on 14 August 2014. Both were encountered around midday, active in the leaf litter along the edge of the trail. Nothing more is known about this species, besides that it is likely a leaf-litter specialist.
Etymology. From the Greek lepros (scaly) and latin auricularis (pertaining to the ear) in reference to the scaly, superficial ear that is a distinctive character in the new species. Suggested common name: Scaly-eared Diminutive Leaf-Litter Skink.
TABLE 1. Summary οf diagnοstic characters (+ present; 0 absent; - missing) amοng species in the genus Tytthoscincus as well as phenοtypically οr genetically similar species οf the genus Sphenomorphus . Οrganized by type οf evidence used fοr inclusiοn as a cοmparisοn. Cοlumn headings are listed under the fοllοwing numbers∶ (1) adult SvL (mm), (2) MBSR, (3) PvSR, (4) vSR, (5) number οf supraοculars, (6) parietal in cοntact with supraοcular, (7) prefοntals in cοntact, (8) number οf lοreals, (9) number οf supralabials, (10) number οf infralabials,) SDL οn tοe Iv, (12) lamellae texture (k = keeled, s = smοοth), (13) enlarged preclοacal scales, (14) dark dοrsοlateral stripe, (15) light pοst-οrbital stripe, and (16) light dοrsοlateral stripe. Data taken frοm the literature is cited frοm∶ (A) (Grismer et al. 2009a); (B) (Steϳneger 1908); (C) (Τaylοr 1918); (D) (Bοulenger 1897); (E) (Kοpstein 1926); (F) (Muller 1895); (G) Dunn 1927); (H) (Grismer & Quah 2015); (I) (Brοngersma 1942); (J) (Datta-Rοy et al. 2013); (K) (Bοulenger 1898); (L) (Annandale 1912); (M) (Τaylοr 1962); (N) (Cοchran 1927); (Ο) Bοulenger 1894); (P) (Darevsky & Οrlοv 2005); (Q) (Quang Nguyen et al. 2011); and (R) (Greer et al. 2006).
Genetic evidence supports placement in Tytthoscincus (Linkem 2013) , but not formally recognized
……continued on the next page TABLE 1. (Cοntinued) Sοme genetic data exists, but phylοgenetic placement is nοt suppοrted.
Despite a SvL> 50 mm, this species is included because it has the same type lοcality as the new species.
Comparisons. Refer to the Introduction and previous Comparisons for background on the choice of species for which to compare to.
MBSR = 35–37 in T. leproauricularis sp. nov. distinguishes it from T. batupanggah sp. nov. (30–31), T. aesculeticola (26–32), T. atrigularis (30), T. biparietalis (32), T. parvus (30), S. bukitensis (31–33), S. butleri (31– 33), S. ishaki (30–32), S. perhentianensis (29), S. sanana (28), S. sibuensis (29), S. textus (30), S. buttikoferi (24), S. malayanus (32–33), S. necopinatus (28–32), S. schlegeli (22), S. senja (32), S. vanhuerni (31), S. apalpebratus (27–28), S. alfredi (26), S. cophias (24), S. courcyanum (24), S. grandisonae (34), S. helenae (30), S. mimicus (30), S. modigliani (32), S. puncticentrallis (29), S. sanctus (32–34), S. scotophilus (28–31), S. sheai (20), S. shelfordi (23–34), S. tanahtinggi (40–42), S. tenuiculus (26), S. tetradactylus (20), S. tonkinensis (32–34), and S. tridigitus (18). PVSR = 68–74 in T. leproauricularis sp. nov. distinguishes it from T. batupanggah sp. nov. (62–66), S. perhentianensis (65), S. sibuensis (58), S. vanhuerni (81), S. apalpebratus (62–64), S. cophias (56), S. macullicolus (79), S. mimicus (61), S. puncticentrallis (64), S. sheai (53), S. tanahtinggi (76–79), S. tenuiculus (57), S. tetradactylus (48), and S. tridigitus (50). VSR = 80–81 in T. leproauricularis sp. nov. distinguishes it from T. batupanggah sp. nov. (55–61), T. aesculeticola (60–68), T. hallieri (63–73), S. bukitensis (61–74), S. butleri (62– 63), S. ishaki (60–70), S. langkawiensis (70–72), S. perhentianensis (61), S. sibuensis (52), S. temengorensis (70,71), S. temmincki (62–72), S. malayanus (74), S. senja (68), S. apalpebratus (60), S. macullicolus (84), S. sheai (54), S. tenuiculus (68), and S. tonkinensis (61–67). The presence of 4 supraoculars distinguishes T. leproauricularis sp. nov. from S. textus (5), S. apalpebratus (5), S. macullicolus (7), S. puncticentrallis (5), S. sanctus (5), S. scotophilus (5), and S. tanahtinggi (5). The contact of the parietal with the last supraocular distinguishes T. leproauricularis sp. nov. from T. hallieri , and S. tetradactylus . The separation of the prefrontals distinguishes T. leproauricularis sp. nov. from all species with prefrontals in contact: T. batupanggah sp. nov., T. atrigularis , T. hallieri , S. bukitensis , S. butleri , S. ishaki , S. langkawiensis , S. perhentianensis , S. sibuensis , S. temengorensis , S. textus , S. buttikoferi , S. malayanus , S. schlegeli , S. courcyanum , S. grandisonae , S. mimicus , S. modigliani , S. puncticentrallis , S. sanctus , S. shelfordi , and S. tonkinensis . The presence of paired anterior loreals distinguishes T. leproauricularis sp. nov. from S. sibuensis , S. macullicolus , S. tanahtinggi , S. tetradactylus , and S. tridigitus . The presence of 6 or 7 supralabials distinguishes T. leproauricularis sp. nov. from S. sanana (5), and S. tanahtinggi (8–9). The presence of 5 infralabials distinguishes T. leproauricularis sp. nov. from T. biparietalis (6), S. perhentianensis (6), S. malayanus (7–8), S. senja (6), S. cophias (7), S. macullicolus (6), S. puncticentrallis (7), S. sanctus (6), S. scotophilus (6), S. tanahtinggi (7), S. tenuiculus (6), S. tetradactylus (7), and S. tonkinensis (6). SDL on Toe IV = 10 in T. leproauricularis sp. nov. distinguishes it from T. batupanggah sp. nov. (9), S. bukitensis (12–13), S. butleri (12–13), S. ishaki (11), S. langkawiensis (11,12), S. sanana (9), S. sibuensis (9), S. textus (15), S. buttikoferi (21–23), S. malayanus (12–13), S. necopinatus (11–15), S. senja (13–17), S. vanhuerni (14–15), S. apalpebratus (8–9), S. alfredi (12), S. cophias (9), S. courcyanum (11), S. grandisonae (12), S. helenae (17), S. macullicolus (18–23), S. mimicus (16), S. modigliani (15), S. puncticentrallis (25), S. sanctus (26–27), S. scotophilus (22–23), S. sheai (6), S. shelfordi (28–29), S. tanahtinggi (16–17), S. tenuiculus (21–24), S. tonkinensis (15–19), and S. tridigitus (7–8). The presence of smooth SDL distinguishes T. leproauricularis sp. nov. from all species with keeled SDL: S. bukitensis , S. ishaki , S. langkawiensis , S. sibuensis , S. temengorensis , S. malayanus , S. apalpebratus , and S. tanahtinggi . The presence of enlarged precloacal scales distinguishes T. leproauricularis sp. nov. from T. aesculeticola , T. biparietalis , T. hallieri , S. bukitensis , S. temmincki , S. apalpebratus , and S. alfredi . The absence of a dark dorsolateral stripe distinguishes T. leproauricularis sp. nov. from T. batupanggah sp. nov., T. atrigularis , T. biparietalis , T. hallieri , S. butleri , S. langkawiensis , S. sibuensis , S. temengorensis , S. temmincki , S. textus , S. buttikoferi , S. necopinatus , S. schlegeli , S. apalpebratus , S. alfredi , S. cophias , S. courcyanum , S. grandisonae , S. helenae , S. mimicus , S. puncticentrallis , S. scotophilus , S. sheai , S. shelfordi , S. tanahtinggi , S. tenuiculus , S. tetradactylus , S. tonkinensis , and S. tridigitus . The absence of a light postorbital stripe distinguishes T. leproauricularis sp. nov. from T. batupanggah sp. nov., T. biparietalis , S. ishaki , S. langkawiensis , and S. temengorensis . The absence of a light dorsolateral stripe distinguishes T. leproauricularis sp. nov. from T. batupanggah sp. nov., S. langkawiensis , S. sanana , S. sibuensis , S. temengorensis , S. schlegeli , S. apalpebratus , S. courcyanum , S. grandisonae , S. helenae , S. sheai , and S. tetradactylus .

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.