
Gomphos shevyrevae, Meng & Kraatz & Wang & Ni & Gebo & Beard, 2009
|
publication ID |
https://doi.org/ 10.1206/673.1 |
|
DOI |
https://doi.org/10.5281/zenodo.5062741 |
|
persistent identifier |
https://treatment.plazi.org/id/785587BB-DB23-FF9E-3798-3BA68104F925 |
|
treatment provided by |
Felipe |
|
scientific name |
Gomphos shevyrevae |
| status |
sp. nov. |
Gomphos shevyrevae , new species
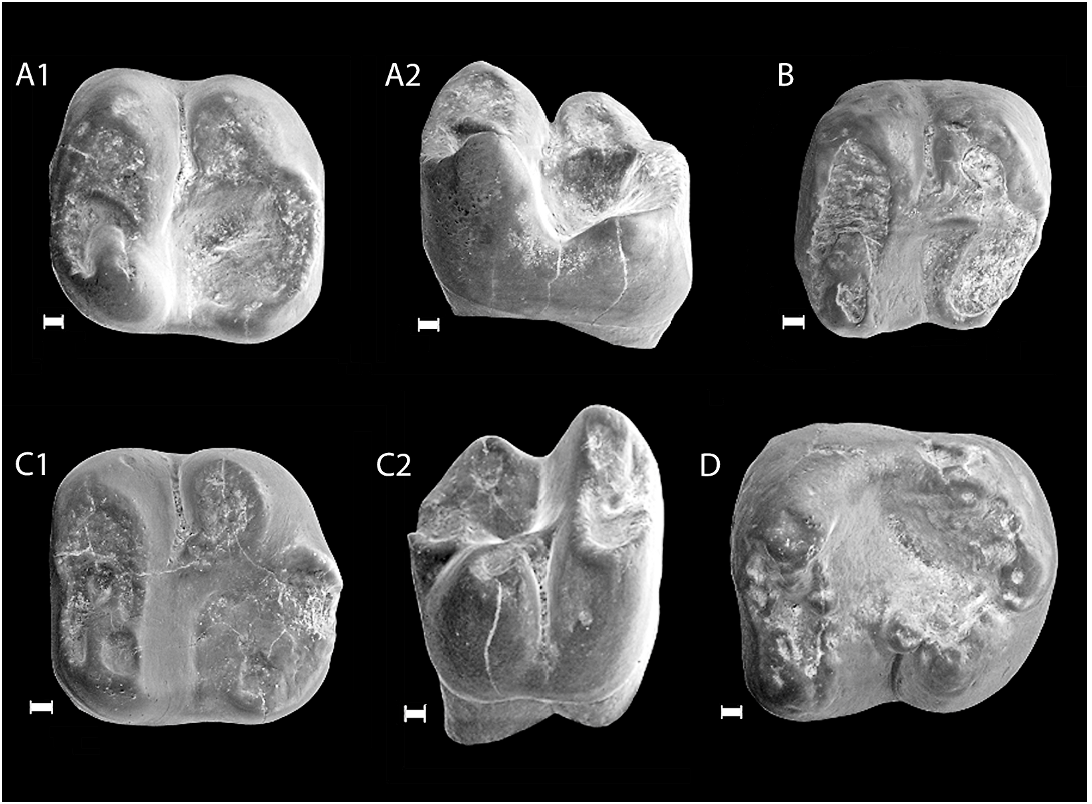
HOLOTYPE: A right M1 ( IVPP V 14669 View Materials ).
PARATYPE: A right m1 ( IVPP V 14670 View Materials ) .
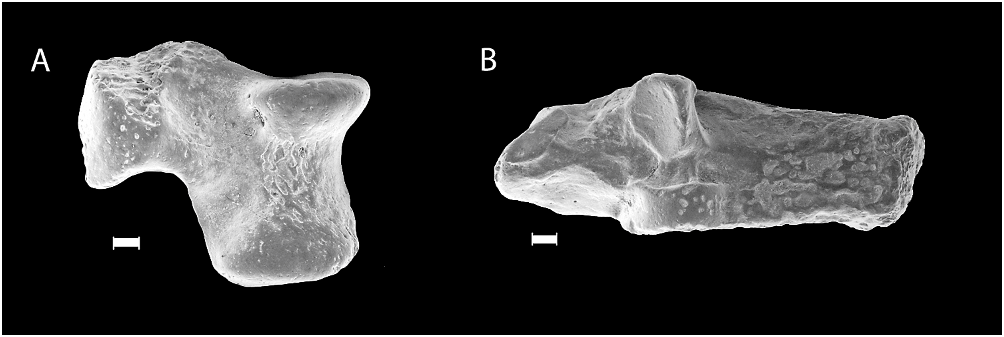
INCLUDED SPECIMENS: A right P4 (or P3) (V14671.1), a right M1 (V14671.2), a right M2 (V14671.3), a left M2 (V14671.4), a right m1 (V14672.1), a right m2 (V14672.2), a left m3 (V14672.3), a left calcaneus ( V14673 View Materials ), and a left astragalus ( V14674 View Materials ).
ETYMOLOGY: The species name is in honor of N.S. Shevyreva who named Gomphos elkema .
DIAGNOSIS: Differs from Gomphos elkema and G. ellae in having more robust teeth with higher crowns, inflated cusps; upper molars with more posteriorly expanded hypocone and shelf; absence of the ridge connecting the lingual and labial cusps on P4; lower molars with relatively long trigonid and short talonid, reduced mesoconid and hypoconulid, and absence of the mesostylid; further differs from G. elkema in having extra facets on the calcaneus for articulation with the astragalus and navicular (unknown for G. ellae ).
TYPE LOCALITY AND AGE: Huheboerhe escarpment, Erlian Basin of Nei-Mongol, Middle Eocene lower beds of the Irdin Manha Formation.
COMPARATIVE DESCRIPTION: Measurements of all teeth are in table 1. The tooth we identified as a P4 could also be a P3 ( fig. 2 View Fig ). However, the anterior border of P3 is usually narrower than the posterior border in Gomphos . This tooth has its anterior portion
TABLE 1 Tooth measurements of Gomphos shevyrevae (in mm)
as wide as the posterior and bears a tiny posterolingual cusp, suggesting that it is likely a right P4. The P4 is a recently erupted tooth that bears no wear. As is typical of Gomphos , the P4 is unilaterally hypsodont and consists of a lingual main cusp, presumably the protocone, and a labial cusp. The lingual cusp is crescent shaped, and its two lophs form the anterior and posterior edges of the tooth. The labial cusp is conical, having a steeper labial surface and a more rounded lingual surface. Unlike P4 of Gomphos elkema , in which P4 is known ( Meng et al., 2004; Asher et al., 2005), the ridge connecting the two cusps is absent in G. shevyrevae ; instead, there is a weak ridge extending from the labial cusp anterolingually to join the midpoint of the anterior loph. The absence of a connection between the lingual and labial cusps of P4 recalls the condition found in Mimotona ( Li, 1977; Li and Ting, 1993).
Upper molars have one major lingual and two minor labial roots. The enamel is thick. M1 is so assigned because its hypocone and shelf are less expanded posteriorly than those of M2, and M1 is usually proportionally shorter than M2 ( fig. 2 View Fig ). M1 of G. shevyrevae is similar to that of G. elkema in general morphology. However, the M1 and other upper cheek teeth of G. shevyrevae are more robust with more inflated cusps and stronger lophs. The protocone and its anterior and posterior lophs form a V-shaped structure with a slightly rounded apex (the lingual side of the protocone). The angle created by the lophs is about 45 °. The lingual portion of the anterior loph may be homologized with the preprotocrista and the labial portion, which is much thinner, with the preparaconule crista. A paraconule (protoconule) is not distinct, but is indicated by a slight inflation where the postparaconule crista joins the anterior loph. Absence of the paraconule is common in basal Glires , both in simplicidentates, such as Matutinia and Rhombomylus ( Ting et al., 2002; Meng et al., 2003), and duplicidentates, such as Mimotona and Gomphos ( Li, 1977; Li and Ting, 1993; Meng et al., 2004), but it is usually present in taxa that are more closely related to rodents and in basal rodents, such as Tribosphenomys ( Meng et al., 1994, 2007b; Meng and Wyss, 2001), Neimengomys ( Meng et al., 2007b) , Archetypomys ( Meng et al., 2007a) , and Cocomys ( Li et al., 1989) .
There is a weak ridge between the paracone and the anterior loph, which is probably the postparaconule crista. The metacone is small- er and more lingually positioned than the paracone. It is connected with the protocone by the posterior loph of the protocone. The latter loph is strong, almost as wide as the metacone. A mesostyle occurs at the posterior base of the paracone, which blocks the labial exit of the narrow trigon basin. The metaconule is present, but is difficult to distinguish in heavily worn specimens. In lightly worn specimens ( fig. 2E View Fig ) a broader wear facet indicates the metaconule that must be higher and broader than the crista in order for such a wear pattern to be created. In G. elkema the posterior loph of the protocone is relatively slimmer and there is usually a restriction labial to the protocone.
The hypocone and the postcingulum are well developed, and both structures are expanded posteriorly. The hypocone is separated from the protocone by a transverse groove, and a similar condition occurs on M2. This condition appears to differ from the morphology prevailing in G. elkema and G. ellae . In well-worn specimens ( fig. 2C View Fig ), however, the protocone and hypocone become confluent. On the lingual surface of the tooth, a vertical groove between the protocone and hypocone exists near the occlusal surface of the tooth, and this feature also seems more pronounced than on upper molars of G. elkema and G. ellae .
M2 is similar to M 1 in general morphology, but it is proportionally longer. The mesostyle is reduced, either being incorporated into the posterior base of the paracone or entirely absent. No M3 was collected.
The lower molars have an anterior and a posterior root. The tooth crown is somewhat higher than that of G. elkema and G. ellae . As in the upper molars, the enamel appears thicker than that of G. elkema and G. ellae . The m1 is identified as such because it has a transversely oriented, ridgelike hypoconulid ( fig 3 View Fig ). In Gomphos , the lower molar hypoconulid becomes more pronounced posteriorly. The m1 and m2 are very similar except that m2 has a more posteriorly projecting hypoconulid. The lower molars are proportionally wider, and therefore relatively shorter, than those of other species of the genus. Lower molar trigonids are proportionally longer in relation to their corresponding talonid than is the case in other species of the genus. As is typical of Gomphos , the protoconid and metaconid are transversely aligned, and the trigonid has nearly vertical anterior and posterior walls. The protoconid and the metaconid are subequal in size. The protoconid is worn more extensively and is therefore lower than the metaconid. The protoconid sends the paralophid (paracristid) to the anterolingual base of the metaconid and the protolophid (protocristid) to the posterior base of the metaconid. A crescentic trigonid basin is enclosed by the trigonid cusps and lophids.
Lower molar talonids are as wide as the trigonid and are longer than their corresponding trigonid. The hypoconid is as large as the protoconid and is very close to the posterior wall of the trigonid, so that the hypoflexid becomes a narrow groove. The cristid obliqua (ectolophid) is short and the mesoconid appears very weak relative to other mimotonids. The entoconid is low. Unlike the condition in G. elkema and G. ellae , in which a mesostylid is present ( Meng et al., 2004; Kraatz et al., 2009), the mesostylid is absent in the new species. Because of the reduction of the mesoconid and cristid obliqua, the talonid basin is a broad, featureless concavity. The hypoconulid is a transverse ridge at the rear of the tooth, being connected with the entoconid by a strong postcristid.
The m3 is little worn and displays several additional minor cuspules on the occlusal surface; these structures would be quickly removed with wear ( fig. 3 View Fig ). The m3 differs from m1 and m 2 in having a longer talonid and a more pronounced hypoconulid. However, compared to those of G. elkema and G. ellae , the talonid of G. shevyrevae is relatively shorter and the hypoconulid is reduced; it does not form a projection at the posterior end of the tooth.
The astragalus measures 11.34 mm long and 8.82 mm wide (maximum dimensions) and is partly broken at its head ( fig. 4A View Fig ). The morphology of the preserved portion is identical to that of Gomphos elkema ( Meng et al., 2004) . The bone is ventrodorsally (or anteroposteriorly) flat. The trochlea is transversely broad and shallow. In G. elkema , the lateral and medial rims of the astragalar trochlea appear to be more asymmetrical, with the lateral much larger than the medial ( Meng et al., 2004).
The calcaneus measures 20.8 mm long and 8.93 mm wide (maximum dimensions; fig. 4B View Fig ). It is typical of Gomphos in that the calcaneoastragalar and sustentacular facets are aligned at the same level, similar to that of lagomorphs ( Bleefeld and McKenna, 1985), but the bone lacks a calcaneal canal that is characteristic of lagomorphs ( Bleefeld and Bock, 2002). The calcaneoastragalar facet is a narrow, long and convex surface in a proximodistal orientation, nearly parallel to the long axis of the bone, whereas the sustentacular facet is rounded and concave. Although the general morphology of the calcaneus is similar to that of G. elkema , it differs from the latter in having a facet on the distal side of the bone, which probably articulates with the navicular and astragalus. This feature does not occur in known astragali of G. elkema ( Meng et al., 2004, 2005; Asher et al., 2005), and is most likely a derived feature of G. shevyrevae . However, in Mimolagus , a similar facet is present on the calcaneus, which is in articulation with the astragalus ( Bohlin, 1951).

| IVPP |
Institute of Vertebrate Paleontology and Paleoanthropology |
| V |
Royal British Columbia Museum - Herbarium |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |