
Cophyla puellarum, Rakotoarison, Andolalao, Crottini, Angelica, Müller, Johannes, Rödel, Mark-Oliver, Glaw, Frank & Vences, Miguel, 2015
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3937.1.3 |
|
publication LSID |
lsid:zoobank.org:pub:5B7B0508-0A35-4566-B979-90FEBF12E2A1 |
|
DOI |
https://doi.org/10.5281/zenodo.5610940 |
|
persistent identifier |
https://treatment.plazi.org/id/782687BD-FFFA-0B69-1DDF-0255FECFFF7D |
|
treatment provided by |
Plazi |
|
scientific name |
Cophyla puellarum |
| status |
sp. nov. |
Cophyla puellarum View in CoL sp. nov.
( Figs. 7 View FIGURE 7 c, 12c & 16)
Remark. This species (ZFMK 57465) has been figured in Glaw & Vences (1994: Fig. 361) as Platypelis sp. and in Glaw & Vences (2007: 135, Fig. 5 View FIGURE 5 d) as unidentified Platypelis from Montagne d’Ambre. Referred to as Cophyla sp. Ca3 by Perl et al. (2014).
Holotype. ZSM 3249/2012 ( ZCMV 13521), adult male, collected at Point de Vue du Grand Moulin in Montagne d’Ambre National Park, -12.54077, 49.15424, 1246 m a.s.l., on 25 January 2012, by A. Rakotoarison and A. Razafimanantsoa.
Paratypes. ZSM 0782/2009 ( ZCMV 13005), male, and ZSM 0781/2009 ( ZCMV 13004) juvenile, both collected at Lac Maudit, Montagne d’Ambre National Park, -12.5856, 49.1457, 1306 m, on 19 and 20 November 2009 by A. Crottini, S. J. Hauswaldt, A. Lima, E. Rajeriarison, F. M. Ratsoavina and A. Razafimanantsoa; UADBA-A 60237 ( FGZC 4903), calling male, ZSM 3274/2012 ( FGZC 4904), calling male, ZSM 3275/2012 ( FGZC 4905), gravid female, UADBA-A 60238 ( FGZC 4906) male, all four collected at Point de Vue du Grand Moulin in Montagne d’Ambre National Park, 12.54077, 49.15424, 1246 m, on 10 December 2012, by A. Rakotoarison and A. Razafimanantsoa; ZFMK 57465, adult female (SVL 33.3 mm), collected at high altitude of Montagne d’Ambre National Park (no coordinates available) on 18–21 March 1994 by F. Glaw, N. Rabibisoa & O. Ramilison.
Diagnosis and comparisons. Assigned to the genus Cophyla in the microhylid subfamily Cophylinae based on enlarged terminal discs on fingers and toes, posterior vomer undivided and not overlapping with neopalatines, absence of nuptial pads and molecular phylogenetic relationships. Among other arboreal species of cophylines the species can be identified by combination of the following character states: body size comparatively large for Cophyla (SVL 27.3–33.6 mm); most of the specimens possess a uniformly yellow-greenish ventral colour; fifth toe slightly longer than third; males with prepollical tubercule but lacking a finger-like prepollex as typical for Anodonthyla ; posterior vomer with well developed and clearly recognizable vomerine teeth; clavicle present.
Distinguished from all other species of Cophyla by a unique advertisement call, characterized by a combination of short calls with long inter-call intervals; ( Table 2 View TABLE 2 ). Further distinguished from C. phyllodactyla , C. maharipeo and C. noromalalae by the presence of a clavicle (no osteological data available for C. berara and C. occultans ). Further distinguished from C. occultans and C. berara by larger body size (adult male SVL 27–29 mm vs. respectively 16–21 mm and 23–26 mm).
Description of the holotype. Adult male in good state of preservation, some muscle tissue removed from right thigh. Snout-vent length 27.3 mm; body slender; head wider than long, not wider than body; snout rounded dorsally and laterally; nostrils not protuberant, nearer to tip of snout than to eye; canthus rostralis almost indistinct, loreal region very slightly concave; tympanum slightly distinct, 46% of eye diameter; supratympanic fold running from posterior edge of eye to forearm; tongue broadly rounded, without papilla, not bifid or notched; weakly developed maxillary teeth present; vomerine teeth not visible, but palpable. Forelimbs slender; subarticular tubercles on all fingers recognizable (single and rounded); outer metacarpal tubercle relatively large and flat; inner metacarpal tubercle oblong and large, forming distinct protuberance at base of first finger; hand without webbing; fingers distinctly flattened and broad along entire length; relative length of fingers 1<2=4<3, fourth finger slightly longer than second; finger discs broadly rounded to slightly bilobate, with lateral fringes; nuptial pads absent. Hindlimbs slender; tibiotarsal articulation reaching between forelimb and tympanum when hindlimb adpressed along body; tibia length, 42% of SVL; inner metatarsal tubercle oblong; outer metatarsal tubercle absent; webbing between toes very weakly developed, with traces of webbing between toes 3–4 and 4–5; subarticular tubercles on toes hardly recognizable; toes flattened and their discs relatively broad, broadly rounded to slightly bilobate, with lateral fringes, smaller than discs of fingers; relative length of toes 1<2<3<5<4 left toes and 1<2<3=5<4 right toes; fifth toe slightly (left) to distinctly (right) longer than third. Dorsal skin smooth, without dorsolateral folds. Ventral skin smooth on throat and chest and heavily granular on belly.
Colour of holotype in preservation. After several years in 70% ethanol ( Fig. 7 View FIGURE 7 c), dorsum brownish with irregular markings, including a darker W-shaped patch extending over the posterior dorsum. Forelimbs and hindlimbs dorsally lighter with dark crossbands. Ventrally, uniformly beige, with some darker colour on venter caused by inner organs shining through the ventral skin. Some dark colour along lower jaws.
Colour in life. Dorsum brownish with yellowish patches that have a greenish shade ( Fig. 16 View FIGURE 16 b). Ventrally uniformly whitish ( Fig. 12 View FIGURE 12 c).
Variation. Most of the paratypes are morphologically similar to the holotype ( Table 3 View TABLE 3 ) except the females ZSM 3275/2012 (FGZC 4905) and ZFMK 57465 which are larger than the males. The third toe is slightly shorter than the fifth toe in all specimens. The female is larger than the males ( Table 3 View TABLE 3 ). There is large variation of coloration between the specimens. In life, dorsum is beige with yellow patches ( Fig. 16 View FIGURE 16 a), or beige and greenish with yellow patches ( Fig.16 View FIGURE 16 b), or brown with yellow patches ( Fig. 16 View FIGURE 16 c), or yellow with brown patches ( Fig. 16 View FIGURE 16 e) for adult and subadult specimens. The paler color pattern probably reflects coloration at night whereas the more contrasted pattern is probably typical for specimens during the day, but our fieldnotes do not allow to confirm unambiguously this hypothesized color change. Dorsum is dark brown with yellow patches and yellow vertebral line in a juvenile specimen ( Fig. 16 View FIGURE 16 d).
Etymology. The name is derived from Latin puellarum : girls, genitive plural of puella. This species is dedicated to the "girls team" of a joint TU Braunschweig and Université d'Antananarivo expedition who discovered the first specimen in 2009 at Lac Maudit.
Natural history. The holotype was found during the day with two metamorphosing tadpoles in a tree hole in highland forest. An ovigerous female (ZSM 3275/2012) contained 21 eggs. The recorded male (UADBA-A 60237) was found on a tree trunk in December 2012 in the same area as the holotype. A second calling male (ZSM 3274/ 2012) was found on a Pandanus leaf. More specimens were regularly observed calling. They were typically heard from low perches of ca. 100–150 cm on leaves, but in a few cases also from higher perches of above 2 m from the ground.
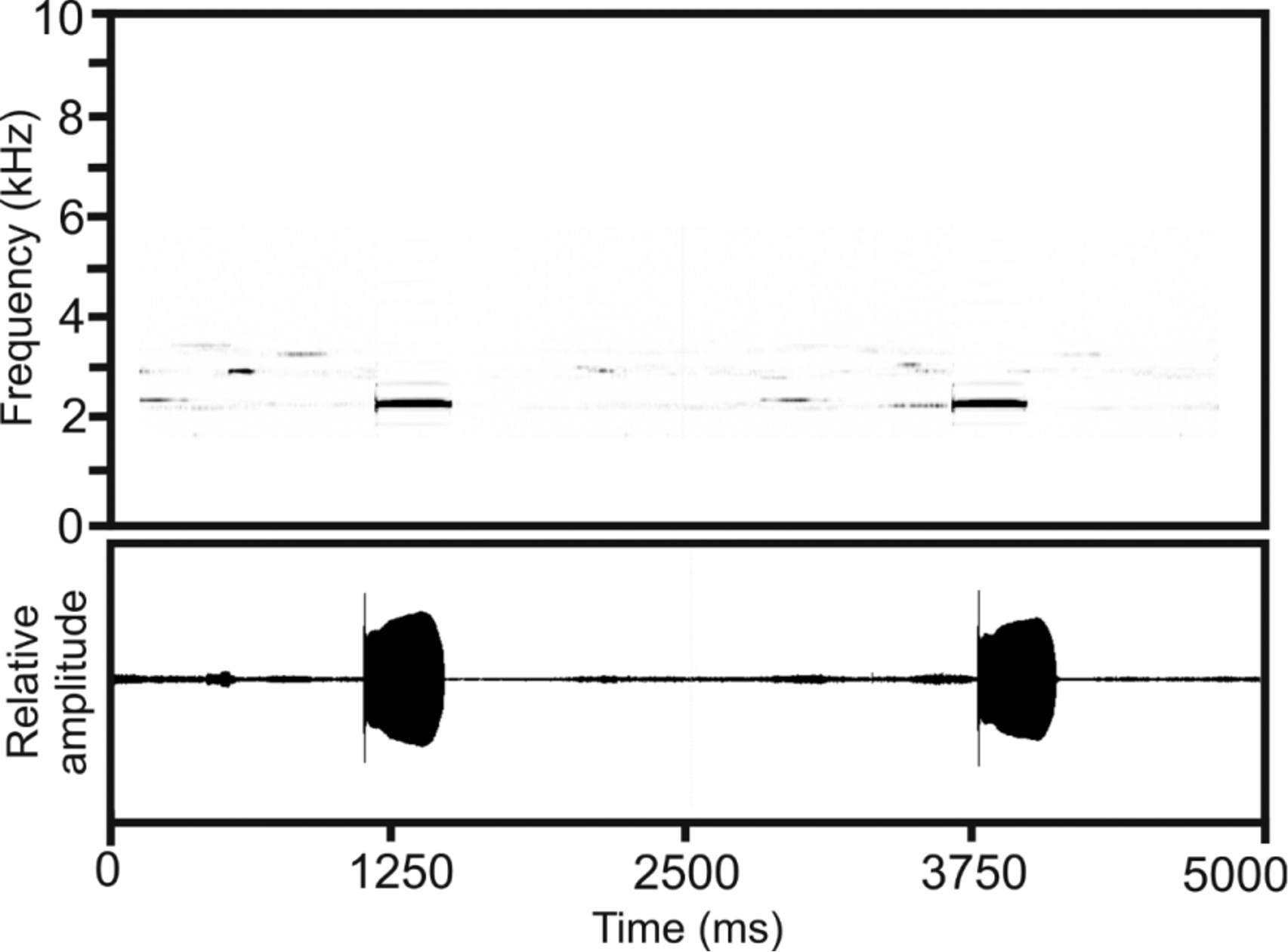
Advertisement call. Recorded from paratype UADBA-A 60237 (FGZC 4903), at Point de Vue du Grand Moulin in MANP, on 10 December 2012, at an air temperature of 16.6˚C ( Fig. 17 View FIGURE 17 ). The following call description refers to the calls of this single specimen. As with all other Cophyla and most cophylines, single tonal calls were repeated in long series (continued at least for several minutes but probably much longer in undisturbed specimens). The dominant spectral call frequency ranged from 2300–2400 Hz (N= 47). Temporal call parameters were as follows: call duration 326–390 ms (363±13 ms; N= 47); duration of inter-call intervals 1961–3996 ms (2640±594 ms; N= 46). As in C. maharipeo , each call in the recorded sequence starts with a very short intensity peak, and we hypothesize this represents an artefact of the recording equipment or an abnormal call feature (see call description of C. maharipeo ).




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.