
Paratelmatobius segallai, Santos & Oliveira & Carvalho & Zaidan & Silva & Berneck & Garcia, 2019
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4648.3.4 |
|
publication LSID |
lsid:zoobank.org:pub:6EFC650A-E3DF-4DC0-932F-3226C476CB57 |
|
persistent identifier |
https://treatment.plazi.org/id/3E3050DE-37E1-463C-B961-1A3293007FBA |
|
taxon LSID |
lsid:zoobank.org:act:3E3050DE-37E1-463C-B961-1A3293007FBA |
|
treatment provided by |
Plazi |
|
scientific name |
Paratelmatobius segallai |
| status |
sp. nov. |
Paratelmatobius segallai sp. nov.
http://zoobank.org/ urn:lsid:zoobank.org:act:3E3050DE-37E1-463C-B961-1A3293007FBA
( Figs. 1–2 View FIGURE 1 View FIGURE 2 ; Table 2 View TABLE 2 )
Holotype. CFBH 43545 View Materials , adult male from Brazil: state of Paraná: municipality of Piraquara: Parque Estadual Pico do Marumbi: Mananciais da Serra : “Salto” trail (25°29′53.18′′S, 48°58′53.45′′W, 1040 m above sea level), collected on 18 January 2018 by M.T.T. Santos, and J.P.C. Monteiro. GoogleMaps
Paratypes. All adults (40 males, and 10 females), collected at the type locality, or in its surroundings . CFBH 43546–43549 View Materials (females) , CFBH 43550–43554 View Materials (males) , UFMG 20769–20776 View Materials (males) , MNRJ 92342–92345 View Materials (males) , MHNCI 10879–10882 View Materials (males), collected on 17–19 January 2018 by M.T.T. Santos, and J.P.C. Monteiro; same locality as the holotype GoogleMaps . CFBH 43555–43557 View Materials (males), collected on 17 January 2018 by M.T.T. Santos, and J.P.C. Monteiro, in the surroundings of the type locality (trail to the water tank “ Carambola ”, 25°29′20.09′′S, 48°58′34.71′′W, 1036 m above sea level) GoogleMaps . MZUSP 137887 View Materials , 137889 View Materials , 137894 View Materials , 137898 View Materials (females) , MZUSP 136882 View Materials , 137888 View Materials , 137890– 93 View Materials , 137895–97 View Materials (males) , UFMG 7335 View Materials (male) , UFMG 7336 View Materials (female) , MHNCI 2970–72 (males), collected on 31 October to 2 November 1999 by P.C.A. Garcia, M.V. Segalla, and R. Lingnau; same locality as the holotype GoogleMaps . CFBH 35790 View Materials (male) , CFBH 35791 View Materials (female) , UFMG 20586 View Materials (male), collected on 11–13 October 2002 by P.C.A. Garcia, and M.V. Segalla; same locality as the holotype GoogleMaps . MZUSP 137560 View Materials (male), collected on 8 October 2005 by M.V. Segalla; same locality as the holotype GoogleMaps .
Generic allocation. We assign this new species to the genus Paratelmatobius based on its morphological similarity to congeners (presence of well-developed fringes on toes, and belly with blotches of vivid coloration in life) and corroborate this inference with a phylogenetic hypothesis based on a mtDNA dataset (detailed below in the section “Molecular phylogenetics and genetic distances”).
Diagnosis. This new species of Paratelmatobius is diagnosable from all congeners by the following combination of characters: (1) advertisement call composed of two note types (A and B); (2) calls emitted in series of 2–4 notes; (3) call duration varying 26–64 ms; (4) note A with 4–10 pulses; (5) absence of an isolated pulse on the onset of notes; (6) dominant frequency (both note types) ranging 2455–3230 Hz; (7) snout not flattened; (8) tympanic annulus visible through the skin; (9) males with vocal slits; (10) presence of a well-developed mandibular tubercle; (11) finger I longer than II; (12) tip of finger III pointed; (13) medial margin of the first toe not fringed or webbed; (14) foot fringed, not webbed (15) presence of tubercles on upper eyelid; (16) nuptial pads with pigmented keratinized spicules; (17) nuptial pads continuous, not divided; (18) toe tips not expanded; (19) dorsolateral fold welldeveloped; (20) in life, belly and ventral surfaces of arms and forearms blotched with orange; (21) in life, external margins of throat lacking orange blotches; (22) adult male SVL 17.2– 23.1 .
Comparisons with congeners (characters of the comparative species in parentheses). Paratelmatobius segallai sp. nov. is distinguished from all congeners by its advertisement call being composed of two note types that are always emitted in a series of 2–4 notes; Fig. 4 View FIGURE 4 A–C (calls composed of a single note type emitted isolated or in intermittent series; Cardoso & Haddad 1990; Giaretta & Castanho 1990; Garcia et al. 2009; Giaretta & Magrini 2013). In addition, the advertisement call of Paratelmatobius sp. nov. distinguishes it from P. cardosoi and P. yepiranga by being shorter: both notes combined range of 26–64 ms (70–130 ms in P. cardosoi, Pombal & Haddad 1999 ; 490–660 ms in P. yepiranga, Garcia et al. 2009 ). The new species further differs from P. yepiranga by having note A composed of 4–10 pulses—note B is not clearly pulsed; Fig. 4 View FIGURE 4 B–C (16–22 pulses; Garcia et al. 2009). It differs from P. gaigeae by the absence of an isolated pulse on the onset of either note A or B (presence; Giaretta & Magrini 2013). It differs from P. poecilogaster by having a higher dominant frequency in both note types: combined range of 2455–3230 Hz (1800–2500 Hz; Giaretta & Castanho 1990). To date, the vocalizations of P. lutzii and P. mantiqueira are unknown.
Paratelmatobius segallai sp. nov. is distinguished from species of the P. lutzii group ( P. lutzii , P. gaigeae , and P. poecilogaster ) by having: snout not flattened (flattened); tympanic annulus present and visible through the skin (tympanic annulus absent in P. lutzii , and concealed by the skin in P. gaigeae and P. poecilogaster ); presence of vocal slits in males (absence); presence of mandibular tubercle (absence); finger I longer than II (finger I shorter than II in P. lutzii and P. poecilogaster , and finger I as long as finger II in P. gaigeae ); tip of finger III pointed (rounded); medial margin of the first toe not fringed or webbed (webbed in P. lutzii , and fringed in P. gaigeae and P. poecilogaster ); nuptial pads in males not divided (divided in two portions); ventral coloration in life with a gray background blotched with orange (black background with white and red stains in P. lutzii ; dark brown background with pinkish red blotches and white dots in P. gaigeae ; and orange background with black, red, and white blotches in P. poecilogaster ). The new species further differs from P. lutzii by the presence of tubercles on the upper eyelid (absence); presence of pigmented keratinized spicules on nuptial pads of males (absence); and foot fringed (foot webbed). The new species is also distinguished from P. poecilogaster by the tips of toes not expanded (expanded). Paratelmatobius segallai is distinguished from P. cardosoi by the presence of tubercles on the upper eyelid (absence); a well-developed mandibular tubercle (poorly developed); and a well-developed dorsolateral fold (poorly developed). It differs from P. mantiqueira by having larger males, SVL 17.2–23.1 mm ( SVL 14.4–16.7 mm); welldeveloped dorsolateral fold (moderately developed); and the tip of finger III pointed (rounded). The new species differs from P. yepiranga by the absence of orange blotches on external margins of throat in life (presence).
Description of holotype. Adult male, SVL 18.4 mm; body robust; head wider than long ( HW /HL 1.15); head width 0.36 SVL; head length 0.31 SVL; snout not flattened, rounded in dorsal and lateral views; END shorter than ED ( END / ED 0.82); canthus rostralis slightly curved in dorsal view and rounded in cross-section; loreal region slightly concave; nostrils slightly protuberant, oval, dorsolaterally directed; interorbital area more than twice as long as ED ( AMD / ED 2.41). Eyes large and protuberant ( ED /HL 0.31; ED / HW 0.27), laterally oriented; upper eyelid margin with small scattered tubercles and a prominent tubercle in its medial region. Tympanic membrane undifferentiated; tympanic annulus medium-sized ( TD / ED 0.55), visible through the skin; supratympanic fold poorly developed, covering the upper portion of tympanic annulus. Well-developed ventrolaterally-oriented mandibular tubercle present on the posterior third of lower jaw. Dorsolateral fold well-developed but fragmented, extending from the posterior corner of the eye to the inguinal region.
Choanae small, nearly rounded, spaced 1.8 mm from each other. Dentigerous processes of vomers poorly distinguishable, oblique, positioned posterior to the level of the choanae, broadly separated from each other. Tongue ovoid, with lateral constriction on its anterior third, free behind for about one third of its length. Vocal slits short, located at the level of the posterior third of tongue; vocal sac externally indistinct. Single, small tooth-like process present on front of lower jaw with a socket between premaxillae.
Forelimb robust, hypertrophied, lacking fold or fringe, well-developed tubercle present on the anterior surface of arm; fingers short, robust; tips of fingers I, II and IV rounded, tip of finger III slightly pointed; relative length of fingers II <I < IV < III; finger I broadly widened, with nuptial pad formed by numerous keratinized spicules covering most of its lateral margin and dorsal surface; subarticular tubercles large, flat, oval in ventral view on finger I, and round in ventral view on fingers II–IV; supernumerary tubercles absent; inner metacarpal tubercle flat, elliptical; outer metacarpal tubercle large, flat, and nearly round in ventral view.
Hindlimb medium-sized and moderately robust ( THL / SVL 0.47; TBL / SVL 0.45), bearing two transverse irregular dermal crests on dorsal surfaces of thigh and tibia; tarsal fold absent; toes medium-sized, robust, with narrow and rounded tips, similar in size to those on fingers; relative length of toes I < II <V < III < IV; toes not webbed, bearing developed and flexible lateral fringes, except in medial margin of toe I; fringes joined between toes II and III, III and IV, and IV and V; subarticular tubercles small, flat, oval in ventral view on toe I, and rounded in ventral view on toes II–V; supernumerary tubercles absent; inner metatarsal tubercle large, flat, and elliptical in ventral view; outer metatarsal tubercle small, nearly rounded in ventral view, conical in profile.
Skin shagreen except on belly (smooth) and on posterior surface of thigh around the cloaca (coarsely granular). Cloacal opening directed posteriorly, at upper level of thighs, covered above by a poorly developed cloacal flap.
Measurements (mm). SVL 18.4, HL 5.8, HW 6.6, ED 1.8, END 1.5, NSD 1.5, IND 2.1, AMD 4.4, TD 1.2, AL 4.6, AW 2.3, FAL 4.5, FAW 2.7, HAL 4.3, THL 8.6, TBL 8.2, TAL 4.3, FL 8.4.
Coloration ( Figs. 1–2 View FIGURE 1 View FIGURE 2 ). In life, dorsum brown with scattered whitish flecks. Dorsal surface between the orbits dark brown. A light brown V-shaped blotch present on posterior third of head delimited anteriorly by whitish flecks, and posteriorly by a thin whitish line. A grayish brown, somewhat quadrangular blotch is present in the scapular region; an oblique band of the same color, connected to this blotch, extends into the inguinal region from each of the posterior corners. A pale yellow middorsal stripe extends from the medial third of the body to cloacal region, which is fragmented anteriorly and continuous posteriorly. Loreal region light brown. A whitish dorsolateral fold extends from behind the eye to inguinal region. Flank with a dark-brown band dorsally delimited by the whitish dorsolateral fold. This band possesses two expansions anteriorly: one triangle-shaped, between eye and arm insertion; the other a vertical bar just posterior to the arm insertion. Ventral to the dark-brown band the flank is brown with several dark brown and blueish flecks and blotches. Tubercles on eyelids, posterior third of mandible, and proximal third of arm whitish. Limbs brown dorsally with scattered whitish flecks and tubercles. Spicules on nuptial pad black. Toes, and dermal crests on thighs and tibia pale yellow. Anterior surface of thigh dark brown. Anterior and posterior surfaces of arm, and inner and outer surfaces of tibia with dark brown transverse bars. An inverted V-shaped thin dark-brown mark extends from the cloacal region to the posterior surfaces of thighs. Ventral surfaces of hand and toes light brown. Ventral surfaces of throat, chest, hindlimbs, and distal third of forearms grayish brown with scattered whitish and blueish blotches and spots. Ventral surfaces of arms and proximal half of forearms orange. Belly with large orange blotches covering almost its entire surface, over a grayish brown background. Ventral surface of inguinal region with small orange blotches. Iris copper-colored with dark brown reticulations.
In preservative, the colors fade, turning into pale tones with scattered melanophores. Dorsal blotches become less distinct. The pale-yellow colors of the middorsal stripe, toes, and dermal crests of the thighs and tibia become whitish. The blueish blotches on flanks and venter become whitish. The orange colors of the ventral surfaces of the forelimbs, belly, and inguinal region become paler ( Fig. 2 View FIGURE 2 ).
Variation. Males are distinguished from females by the presence of nuptial pads, and hypertrophy of arms and forearms (males AW/AL 0.44–0.59, mean 0.52; females AW/AL 0.31–0.39, mean 0.36; and males FAW/ FAL 0.50–0.66, mean 0.58; females FAW/ FAL 0.35–0.42, mean 0.39; Table 2 View TABLE 2 ). In addition, males possess wider thumbs than females. In general females are larger than males (females SVL mean = 20.6 mm, n = 9; males SVL mean = 18.5 mm, n = 37), but a male specimen ( CFBH 43552) is the largest individual of the type series (maximum values of all measurements taken correspond to this individual; Table 2 View TABLE 2 ). The tubercles on posterior third of the mandible and on proximal third of arm vary in degree of development. The middorsal stripe varies in size, width, and degree of fragmentation. In some specimens the stripe is large and reaches the quadrangular blotch in the scapular region ( MHNCI 10881, MNRJ 92343, 92345, UFMG 20770, 20775), while in others it is very small and covers just the posterior third of the body, reaching the cloacal region ( CFBH 43551, 43554, MHNCI 10880, 10882, UFMG 20769, 20774). Some specimens possess an orange blotch on the posterior third of the tarsus ( CFBH 43555, 43549, MHN- CI 10880, 10882). Some specimens possess few ( CFBH 43549, MHNCI 10880, MNRJ 92343) or several ( UFMG 20773) white spots in the pectoral region. One specimen ( CFBH 43552) possesses a lighter coloration of the dorsum and the blotches of the belly, which are more yellowish. The mandibular tubercle of one specimen ( MHNCI 10881) is orange. One specimen ( MNRJ 92342) has dark brown labial bars and possesses a greater number of dark brown bars and blotches on the limbs, and a darker band on the flanks. The coloration of the specimens that were collected between 1999 and 2005 are more faded (dorsum light brown, blotches of belly cream and less visible markings).
Tadpole. Maximum total length 19.95 mm (at stage 34; Table 3 View TABLE 3 ). Body 0.37–0.40 times TL, depressed (BH/ BW = 0.70–0.83), elliptical in dorsal view, rounded depressed in lateral view, with peribranchial region slightly convex, and abdominal region convex ( Fig. 3A, B View FIGURE 3 ). Snout rounded in dorsal ( BWN / BWE = 0.66–0.73) and lateral views. Eyes 0.15–0.19 times BW, dorsally located ( IOD / BWE = 0.49–0.57), dorsolaterally directed. Nostrils elliptical, dorsal, located halfway between eyes and tip of snout ( NSD / ESD = 0.43–0.53), anterolaterally directed and with a continuous and elevated rim, without fleshy projection. Spiracle sinistral, lateral ( SDEH /BH = 0.38–0.43), posterodorsally directed, 0.10–0.13 times BL, opening between the middle and posterior thirds of body ( SSD / BL = 0.65–0.72); inner wall fused to body with a small free edge. Lateral line system indistinct. Intestinal switchback point located at the center of abdominal region or dislocated to the left ( Fig. 3C View FIGURE 3 ). Vent tube dextral, posteriorly directed, small, fused to ventral fin and positioned at its ventral margin ( Fig. 3A, C View FIGURE 3 ). Tail with medium height (TH/ TAL = 0.2 9–0.33), moderately slender musculature ( TMH /BH = 0.40–0.46), not reaching the rounded tail tip; dorsal fin height approximately the same as that of ventral fin ( AND / ANV = 1.02–1.10). Dorsal fin height 0.09–0.13 times TAL, with slightly convex external margin; emerging on the posterior third of body; maximum height at the posterior third of tail. Ventral fin height 0.08–0.12 times TAL, with slightly convex external margin; originates at base of tail, concealed by vent tube ( Fig. 3A View FIGURE 3 ). Oral disc 0.38–0.45 times BW, ventral, laterally emarginated; a single row of marginal papillae aligned on the anterior labium and alternated on the posterior labium, with a wide dorsal gap; papillae reduced or absent on lateral emarginations; one submarginal papilla anteriorly on each side; labial tooth row formula ( LTRF) 2(2)/3(1), A1=A2; P1=P2=P3; jaw sheaths narrow, finely serrated; upper jaw sheath arc-shaped and lower sheath “V”-shaped ( Fig. 3D View FIGURE 3 ).
Coloration of tadpole in preservative. Body brown with scattered dark brown dots; caudal musculature beige with scattered dark brown dots, mainly on its anterior third; dorsal and ventral fins, spiracle and myomeres on medial and posterior thirds of tail musculature translucent; few to numerous small white dots scattered along the body, dorsal and ventral fins and caudal musculature. The intestinal mass is visible both ventrally and laterally.
Comparison with congeners’ tadpoles. Tadpole morphology is known for five species of Paratelmatobius : P. cardosoi ( Cardoso & Haddad 1990; Pombal & Haddad 1999), P. poecilogaster ( Giaretta & Castanho 1990; Pombal & Haddad 1999), P. yepiranga ( Garcia et al. 2009) , P. gaigeae ( Domenico et al. 2014) , and P. segallai (present study). Tadpoles of P. lutzii and P. mantiqueira remain undescribed. The tadpole of P. segallai is distinguished from P. poecilogaster by possessing a single row of alternated marginal papillae on the posterior labium (three rows; Pombal & Haddad 1999), and shorter total length— 19.9 mm at Stage 34 (25.8 mm at Stage 37; Pombal & Haddad 1999). It differs from P. gaigeae by possessing an elliptical body in dorsal view (ovoid), a single row of marginal papillae aligned on the anterior labium and alternated on the posterior labium (single row, alternated on anterior and posterior labia), a wider dorsal gap of marginal papillae (less wide), and narrow jaw sheaths (wide). It differs from P. yepiranga by the absence of submarginal papillae on the posterior labium (presence) and by possessing narrow jaw sheaths (wide). It differs from P. cardosoi by possessing a single row of marginal papillae aligned on the anterior labium and alternated on the posterior labium (single row, alternated on anterior and posterior labia).
Advertisement call. The vocalization of P. segallai ( Fig. 4 View FIGURE 4 ) consists of a series of 2–4 notes (n = 30; calls of two males). Notes comprising the advertisement call (= note series) are classified into two distinct types: notes A and B. Note A is defined by having well-marked pulses throughout the note and a higher amplitude in comparison with note B. On the other hand, note B possesses varying degrees of amplitude modulations (AMs), from a pattern of no modulation throughout the note, to an intermediate pattern of AMs, but irregularly distributed and/or weakly modulated in these notes, to a pattern of several AMs throughout the note, with variable modulation rates and depths between peaks and valleys ( Fig. 4 View FIGURE 4 ). Because of this variation we chose not to apply the term “pulse” to the highly variable AM patterns present in notes B.
The two note types are arranged in different combinations, with two-note calls “B + A” and three-note calls “B + A + B” being the most frequent emission patterns of one of the recorded males (57% and 36% of calls, respectively; recording MTTS 001, voucher CFBH 43545, holotype), whereas two-note calls “B + A” and three-note calls “B + A + A” were equally frequent in the other recorded male (50% of calls each; recording MTTS 002, unvouchered). The emission of isolated notes (either A or B) was rarely observed in call recordings and only on occasions when two males were calling in antiphony, which we assume to be a variant of the most frequent calling emission patterns described earlier related to male-male acoustic interactions. Calling rate ranged 33–69 calls per minute (50.9 ± 25.3; n = 2 males), and the interval between notes (in a note series) lasted 74–177 (1 11.7 ± 26.8; n = 44) ms.
Note A lasted 27–61 (44.9 ± 5.7; n = 40) ms, with rise time ranging 2–65 (32.3 ± 5.9; n = 40) %. Pulse num- ber ranged 4–10 (6.6 ± 0.6, n = 41), and was emitted at a rate of 125–222 (147.1 ± 4.7; n = 41) pulses per second. The linear frequency modulation throughout the note was very subtle. The dominant frequency ranged 2756–3230 (2966 ± 140; n = 40) Hz, the minimum frequency 2412–2885 (2657 ± 96; n = 40) Hz and the maximum frequency 3058–4091 (3365 ± 223; n = 40) Hz. Note B lasted 26–64 (45.0 ± 9.0; n = 33) ms, with rise time ranging 4–83 (43.6 ± 10.8; n = 33) %. The linear frequency modulation throughout the notes was very subtle. The dominant frequency ranged 2455–3187 (2764 ± 86; n = 33) Hz, the minimum frequency 1852–2541 (2210 ± 96; n = 33) Hz, and the maximum frequency 2842–4433 (3400 ± 588; n = 33) Hz.
Geographic distribution. Paratelmatobius segallai is known to occur only at its type locality (municipality of Piraquara, state of Paraná, in the Serra do Mar mountain range, southern Brazil), which represents the southernmost record of the genus ( Fig. 5 View FIGURE 5 ). For geographic coordinates, see the sections “ Holotype ” and “ Paratypes ”.
Natural history. The new species occurs in remnants of high montane dense ombrophilous Atlantic Forest, in well-preserved fragments. We visited the type locality (Piraquara, state of Paraná) in October-November 1999, October 2002, October 2005, January 2006, November 2017, and January 2018. Despite drizzly weather in November 2017, we did not observe any vocalizing males of Paratelmatobius segallai nor did we find any individuals. On all other occasions we visited the area during and after long periods of heavy rains (at least a week) and found many individuals near temporary pools, some of which were calling. In January 2018, we observed many individuals of P. segallai (males, females and amplectant pairs) exposed and/or walking on the leaf litter shortly after a heavy rain ceased. According to our observations, P. segallai can be classified as an explosive breeder ( Wells 1977), with its reproductive activity being strongly associated with heavy rainfall.
Adult males of Paratelmatobius segallai call at night and occasionally during the day from within or on the edge of small, shallow muddy-bottom pools, usually positioned with their body partially submerged. When disturbed, individuals dive quickly to the bottom of the pools. These pools were usually next to rivulets. Amplexus is axillary. Eggs are submerged and attached to leaves or to the margin of pools. Clutches are composed of few eggs (less than 30) that are relatively large and uniformly dark-pigmented. Larvae of P. segallai are exotrophic, lentic and benthic (guild IIA 1; McDiarmid & Altig 1999), completing their development in temporary pools inside the forest.
Another species of Paratelmatobiinae, Scythrophrys sawayae , was syntopic with Paratelmatobius segallai at the type locality. Despite males of both species calling simultaneously, their advertisement calls were easily distinguishable by temporal (e.g., pulse number and rate) and spectral (dominant frequency) traits. Individuals of S. sawayae were more abundant and shared the calling and breeding microhabitat with P. segallai .
Molecular phylogenetics and genetic distances. The optimal MAFFT similarity-alignment comprised 2440 characters. The BIC in Partition Finder identified one subset, GTR + I + G, for 12S, tRNAvaline, and 16S, BIC: 29876.8386125. The Bayesian phylogenetic inference analysis recovered a monophyletic and well-supported Paratelmatobius ( BPP = 100), and placed P. segallai in the P. cardosoi group ( Fig. 6 View FIGURE 6 ). Within this group, P. segallai was recovered as the sister taxon of Paratelmatobius cf. segallai , a morphologically related lineage for which more data are still needed for a specific assignment (see the section Remarks below). The clade comprising these taxa formed the sister group of P. cardosoi + Paratelmatobius cf. cardosoi . These four taxa formed the sister group of the sampled species of the P. lutzii group ( P. gaigeae and P. poecilogaster ). All relationships among recognized species and lineages with uncertain taxonomic status of Paratelmatobius were well-supported ( BPP = 100). The uncorrected p-distances of H1 fragment between recognized species of Paratelmatobius varied from 6.4% to 11.9% (Table 4).
Etymology. The specific epithet honors the Brazilian herpetologist Magno V. Segalla for having collected the first specimens of the new species, and for his substantial contributions to the study and conservation of amphibians in southern Brazil.
Remarks. We carefully examined the external morphology of specimens from the municipalities of Capão Bonito, Guapiara, Iporanga, Piedade, Pilar do Sul, São Miguel Arcanjo, and Tapiraí, in the state of São Paulo (Appendix I), including the vouchers CFBH 8365, 16536, and 43420, which were included in our phylogenetic analysis ( Paratelmatobius cf. segallai ; Fig. 6 View FIGURE 6 ). The morphology of these specimens is largely congruent with that described for the type series of P. segallai , considering all the morphological diagnostic characters that were possible to evaluate (characters # 7–19 and 22). We only found some variation in the arrangement of blotches on belly. In some specimens the blotches are similar to those of the type series of P. segallai (e.g., CFBH 8365, 8370, 16535–16536, 40684, 40686), whereas in other specimens the blotches may be small and sparse (e.g., CFBH 3724–3726, 8366, 43420–43421, 40681, 40789, 40794), or medium-sized and slightly merged (e.g., CFBH 3722–3723, 8367–8368, 16537, 23302, 36008–36011, 38426–38427). A few specimens possess a midventral line crossing the throat and the pectoral region ( CFBH 16536, 38427) or only the pectoral region ( CFBH 8366).
In addition, we accessed the only available call recording for the populations of São Paulo State ( APA 001, housed at CFBH from the municipality of Pilar do Sul). Because of the low sound input of calls recorded, we were able to analyze only some qualitative features. These features are in accordance with the diagnosis of P. segallai (characters # 1–2, 5; note A is pulsed).
However, karyotypic differences between specimens from the type locality of Paratelmatobius segallai (Piraquara) and specimens of Paratelmatobius cf. segallai from São Miguel Arcanjo were reported by Lourenço et al. (2008): e.g., the nucleolar organizer region ( NOR) site is adjacent to a large heterochromatic block of the short arm of the chromosome 9 in specimens from Piraquara, whereas it is in the long arm of chromosome 8 in specimens from São Miguel Arcanjo.
The uncorrected p-distance between Paratelmatobius segallai and Paratelmatobius cf. segallai (2.9–3.1%) is similar to the distances between P. cardosoi and Paratelmatobius cf. cardosoi (2.5%) and between the two lineages of P. poecilogaster (4.0%); Table 4. These distances might indicate that there are structured intraspecific lineages within these taxa; alternatively, this might indicate the presence of cryptic distinct evolving lineages corresponding to undescribed species. In the case of P. segallai and Paratelmatobius cf. segallai , the genetic distances can also be attributed to the sampling gap (i.e., 135 km) between the single occurrence locality for P. segallai and the southernmost sampled locality for Paratelmatobius cf. segallai (see distribution point 6 in Fig. 5 View FIGURE 5 ). Therefore, a more comprehensive study, including specimens from other localities and with the addition of nuclear DNA fragments and other mitochondrial fragments, is required for a better understanding of the diversification processes that occurred in Paratelmatobius .
We used an integrative approach to describe the new species, with evidence from adult and tadpole morphology, advertisement call, and DNA sequences. For this reason and considering the karyotypic differences reported by Lourenço et al. (2008), we believe that additional data are necessary to an accurate assignment of the populations in São Paulo State.



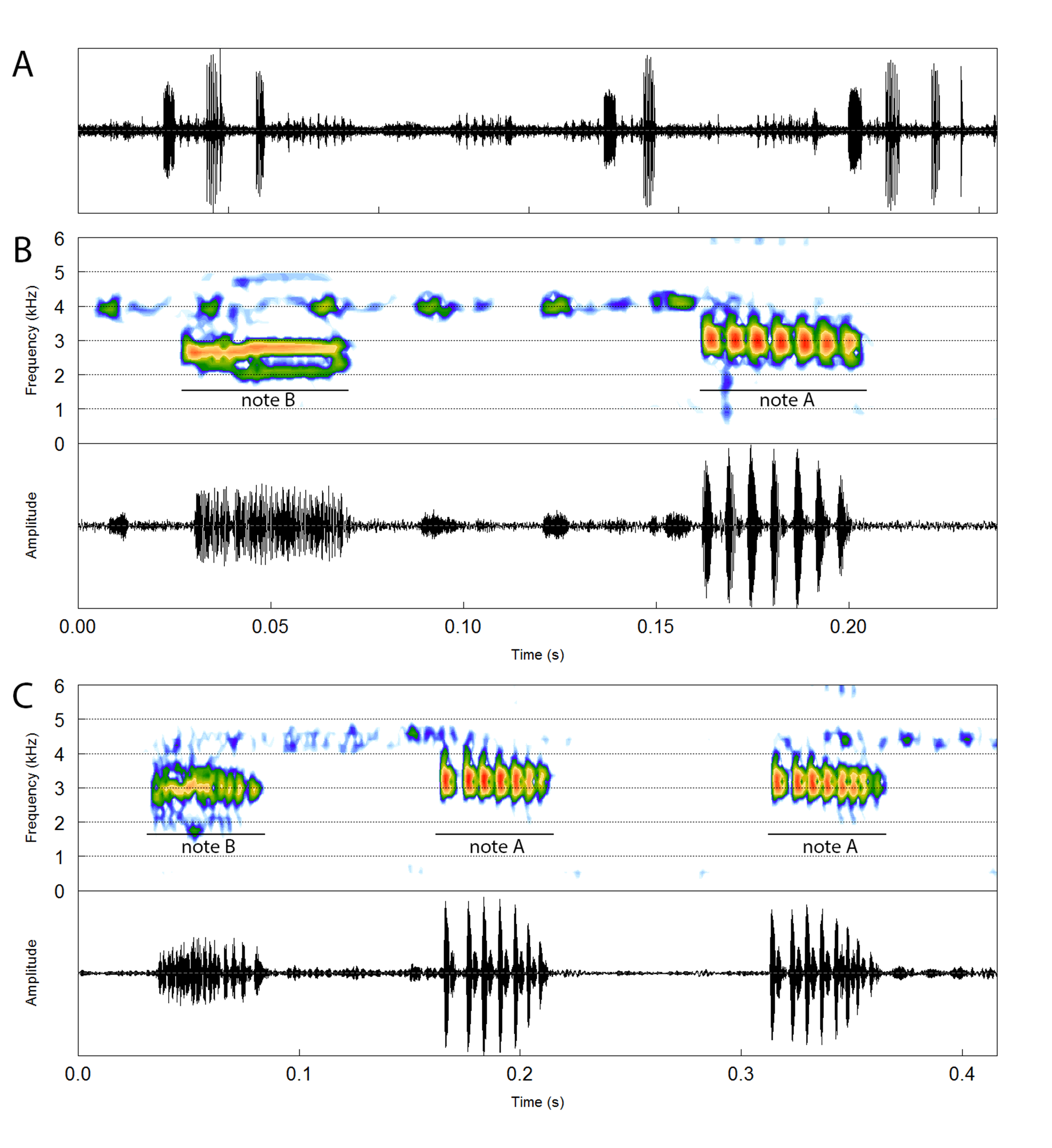
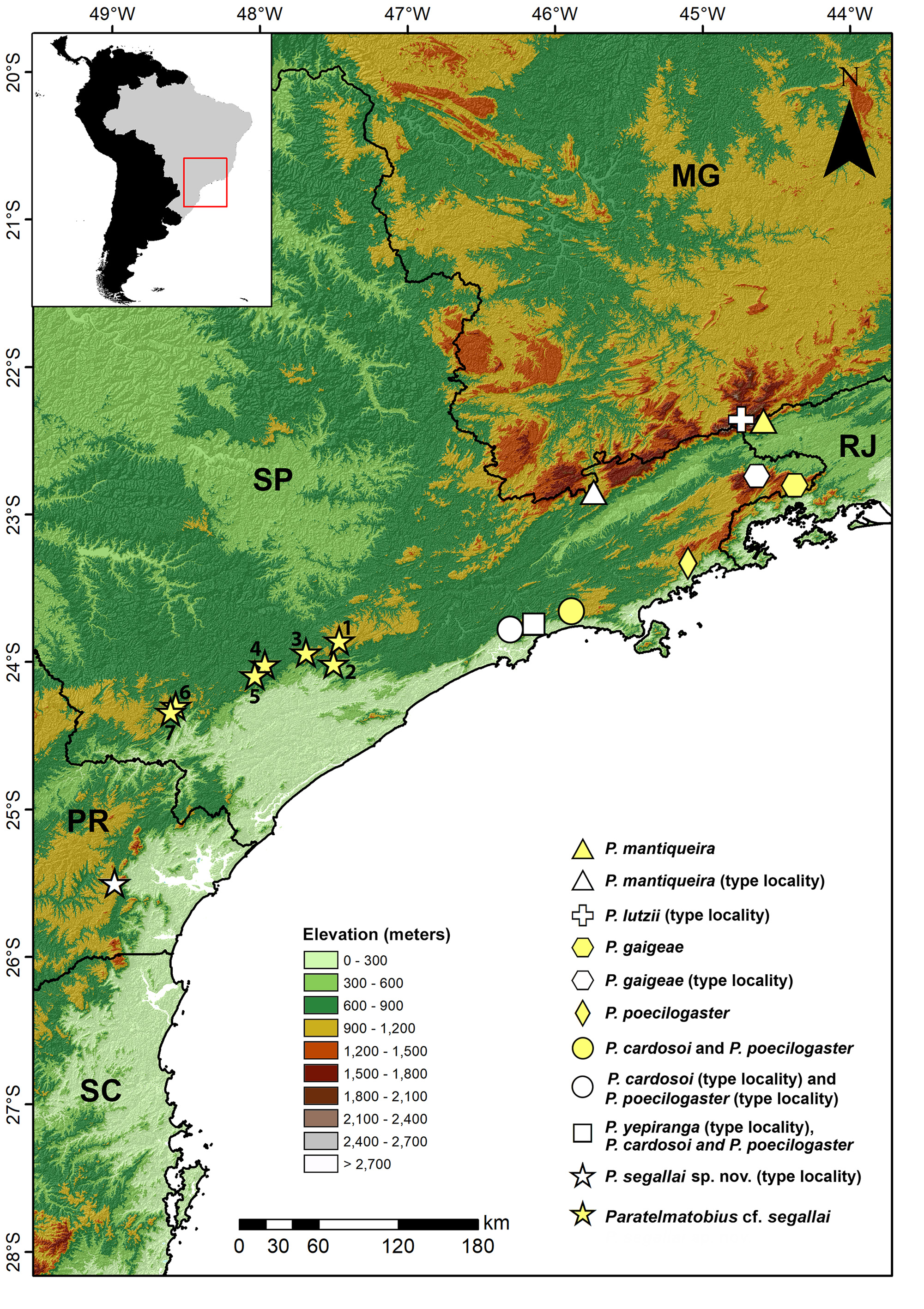

TABLE 2. Measurements and proportions for the type series of Paratelmatobius segallai. Values (mm) are reported as ranges (mean ± SD). See text for explanation of measurement abbreviations.
| Measurement | Body ratios | ||||
|---|---|---|---|---|---|
| Males (n = 37, | Females (n = 9, | Males (n = 37) | Females (n = 9) | ||
| SVL | except for SVL, n = 41) 17.2–23.1 (18.6 ± 1.0) | except for SVL, n = 10) 19.5–22.0 (20.6 ± 0.8) | HW / HL | 1.13–1.18 | 1.12–1.18 |
| HL | 5.5–7.1 (5.9 ± 0.2) | 6.3–6.8 (6.6 ± 0.2) | HL / SVL | 0.30–0.33 | 0.31–0.34 |
| HW | 6.4–8.3 (6.8 ± 0.3) | 7.4–8.0 (7.6 ± 0.2) | HW / SVL | 0.38–0.37 | 0.36–0.38 |
| ED | 1.8–2.2 (1.9 ± 0.1) | 1.9–2.1 (1.9 ± 0.1) | END / ED | 0.74–0.85 | 0.77–0.86 |
| END | 1.4–1.8 (1.5 ± 0.1) | 1.5–1.7 (1.6 ± 0.1) | AMD / ED | 2.24–2.52 | 2.35–2.65 |
| NSD | 1.4–2.0 (1.6 ± 0.1) | 1.5–1.8 (1.7 ± 0.1) | AMD /HL | 0.72–0.80 | 0.69–0.77 |
| IND | 2.0–2.7 (2.1 ± 0.1) | 2.3–2.5 (2.4 ± 0.1) | AMD / HW | 0.62–0.70 | 0.61–0.67 |
| AMD | 4.2–5.4 (4.5 ± 0.2) | 4.7–5.0 (4.8 ± 0.1) | ED / HL | 0.29–0.35 | 0.28–0.30 |
| TD | 1.1–1.5 (1.1 ± 0.1) | 1.2–1.5 (1.4 ± 0.1) | ED / HW | 0.25–0.31 | 0.24–0.27 |
| AL | 4.3–6.1 (4.7 ± 0.3) | 4.3–4.9 (4.6 ± 0.2) | TD / ED | 0.56–0.66 | 0.62–0.74 |
| AW | 2.0–3.4 (2.4 ± 0.2) | 1.5–1.9 (1.7 ± 0.1) | TD / HL | 0.18–0.21 | 0.18–0.22 |
| FAL | 4.1–5.9 (4.5 ± 0.3) | 4.7–5.6 (5.0 ± 0.2) | AW / AL | 0.44–0.59 | 0.31–0.39 |
| FAW | 2.1–3.8 (2.6 ± 0.3) | 1.7–2.3 (2.0 ± 0.2) | FAW / FAL | 0.50–0.66 | 0.35–0.42 |
| HAL | 4.0–5.6 (4.4 ± 0.3) | 4.2–4.9 (4.6 ± 0.2) | FAL / SVL | 0.22–0.27 | 0.22–0.27 |
| THL | 8.1–10.8 (8.6 ± 0.5) | 8.7–9.5 (9.1 ± 0.3) | HAL / SVL | 0.22–0.25 | 0.21–0.25 |
| TBL | 7.7–10.5 (8.4 ± 0.4) | 8.3–9.1 (8.7 ± 0.2) | THL / SVL | 0.44–0.48 | 0.43–0.46 |
| TAL | 4.3–5.6 (4.6 ± 0.3) | 4.7–5.3 (4.9 ± 0.2) | TBL / SVL | 0.42–0.48 | 0.41–0.44 |
| FL | 7.5–10.2 (8.5 ± 0.4) | 8.3–9.6 (8.9 ± 0.4) | FL / SVL | 0.43–0.49 | 0.39–0.46 |
| MHNCI |
Museu de Historia Natural Capao de Imbuia (Brazil) |
| AMD |
National Herbarium of the Netherlands, Hugo de Vries-Laboratory |
| THL |
Grierson Museum |
| CFBH |
Universidade Estadual Paulista |
| MNRJ |
Museu Nacional/Universidade Federal de Rio de Janeiro |
| UFMG |
Universidade Federal de Minas Gerais |
| SSD |
Sammlung Simon des Stattlichen Museum fur Mineralogie und Geologie Dresden |
| TAL |
Jardin botanique de Talence |
| TMH |
Tasmanian Museum and Art Gallery |
| AM |
Australian Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Paratelmatobiinae |
|
Genus |