
Oviraptor, PHILOCERATOPS
|
publication ID |
https://doi.org/ 10.1206/0003-0082(2002)364<0001:CAOCOT>2.0.CO;2 |
|
DOI |
https://doi.org/10.5281/zenodo.5604439 |
|
persistent identifier |
https://treatment.plazi.org/id/64443907-2845-7E7B-FD70-9A62FC92ADA6 |
|
treatment provided by |
Felipe |
|
scientific name |
Oviraptor |
| status |
|
A REINTERPRETATION OF THE SKULL OF THE HOLOTYPE OF OVIRAPTOR PHILOCERATOPS
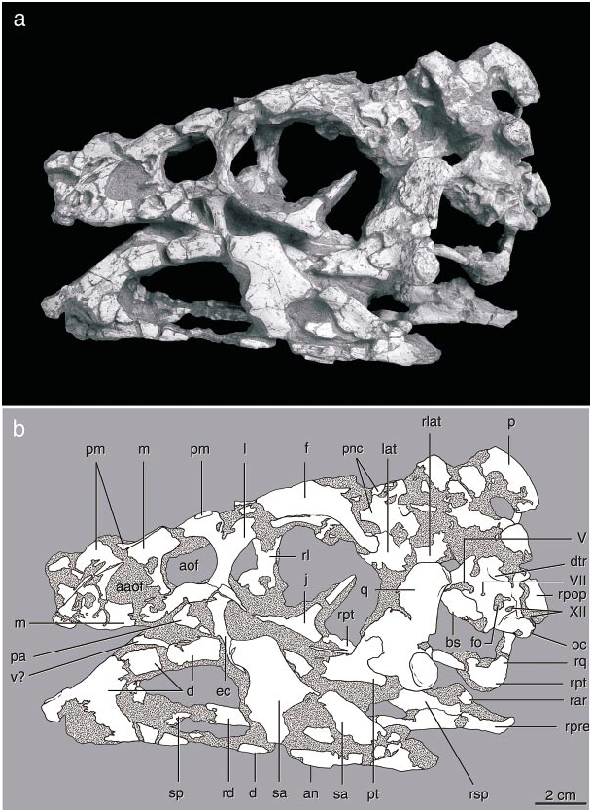
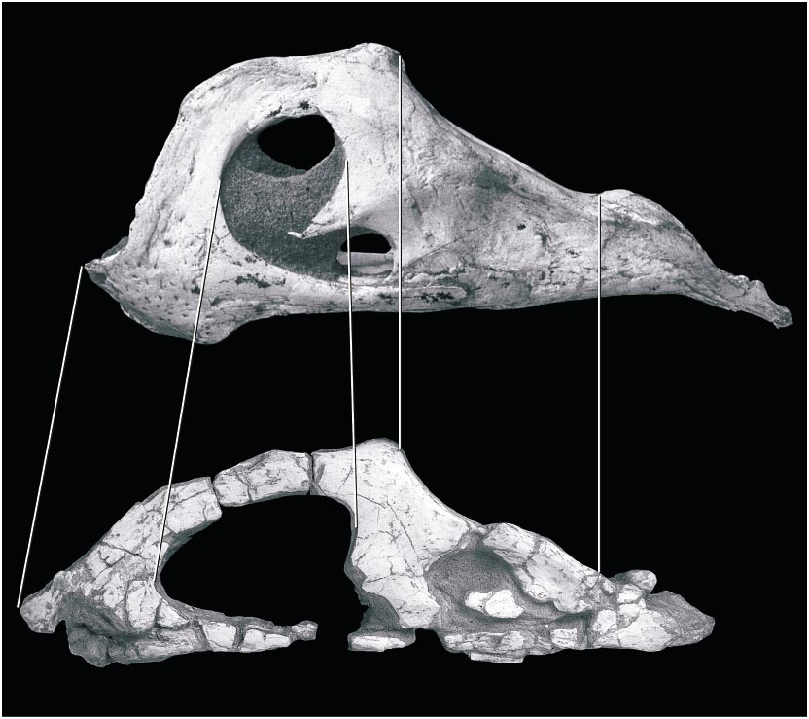
Beginning with its brief description by Osborn (1924), the morphology of the holotype of Oviraptor philoceratops, AMNH 6517, has been poorly understood. The skull ( fig. 11 View Fig ) is crushed and abraded, and initially was incompletely prepared, lying on the original slab adjacent to the skeleton. It was given its most detailed description by Smith (1993), who attempted a reconstruction of the skull and interpreted its functional morphology. Comparison between the new oviraptorid material described above and the holotype of O. philoceratops elucidates several features that previously were enigmatic.
The skull of AMNH 6517 has been crushed mediolaterally, compressing the temporal region. Thus, its breadth across the occiput is only about half that of the skull of the holotype of Citipati osmolskae , which is similar in length. The braincase and right temporal regions have been dislocated as a unit posteriorly. The broken pieces of the individual bones have been separated from one another to varying degrees along a longitudinal axis, to a greater degree on the right side than on the left. This is most evident in the mandible, especially a large gap within the dentary on the right side. The frontals are incomplete dorsally, exposing the roof of the braincase and the interorbital space. The right jugal is missing except for its postorbital process and its anterior tip, and the left jugal is missing posterior to the postorbital bar, as is the left quadratojugal.
Smith (1993) did not have the benefit of wellpreserved material for comparison, and the Ukhaa Tolgod material suggests that several identifications and interpretations of the distortion of the skull in that paper require modification. The dorsal side of the palatal complex is not ‘‘rotated as a unit and exposed on the left side of the skull’’ ( Smith, 1993: 367). Instead, the ectopterygoid and the anterior end of the palatine are in the typical position of oviraptorids in facing laterally rather than ventrally ( Osmólska, 1976; Elzanowski, 1999). Thus the dorsal edge of these bones as preserved is comparable to the lateral edge in other theropods, not the medial edge as Smith’s interpretation would suggest. Furthermore, a bone identified as the ectopterygoid is actually the quadrate (see below). The occiput has not been ‘‘rotated... so that the dorsal side faces the left side of the skull’’ ( Smith, 1993: 368). The occiput faces posteriorly in line with the remainder of the skull and is not rotated, although it is slightly compressed mediolaterally.
Smith described the skull as amphikinetic, but it is unclear whether the joints he identified were capable of movement. The ‘‘mesokinetic hinge between the frontal and parietal bones’’ ( Smith, 1993: 369) is not apparent, as the contact between frontal and parietal is not preserved and the edges of these two bones are broken. By comparison, this contact on the skull of Citipati is oblique on each side, with the parietal extending far forward along the midline, rather than transverse as in taxa with a hinge joint, such as most squamates. Whether or not movement was possible at the jugallacrimal or quadratesquamosal contacts is not readily apparent from the holotype of O. philoceratops , because the latter contact (preserved only on the right side) is concealed in matrix and the exoccipital contribution to the joint is not preserved. Furthermore, the force vectors reconstructed for the mandibular adductor muscles do not take into account the posterior displacement of the parietal, and thus should be more vertically oriented.
The left quadrate was identified as the ectopterygoid by Smith, from which he deduced a significant amount of distortion to the palate. (This bone is labeled ‘‘ec’’ in Smith’s [1993] figures 2 View Fig and 4 View Fig , but this label is defined as the ectopterygoid in figure 2 View Fig and the exoccipital in figure 4 View Fig of that paper; in the text [ Smith, 1993: 374] it is described as the ectopterygoid of the right side.) The unusually round shape of the skull in occipital view proposed by Smith (1993: fig. 3B View Fig ), with a ventrolaterally directed quadrate, is apparently due to this misidentification. However, the intact left quadrate is oriented vertically with a broad dorsal ramus, as in other oviraptorids. Furthermore, most of the ventral portions of what is figured as the right squamosal ( Smith, 1993: fig. 4H View Fig ) are instead parts of the quadrate. On the right side, the lateral surface of what was identified as the right quadrate is the quadratojugal.
The mandibular condyles of the quadrate on the left side are well preserved. The quadrate is similar to that of IGM 100/ 978 in having a welldeveloped lateral condyle and a smaller medial condyle. The surface generally is oriented transversely and convex ventrally, but the condyles are separated by a groove running slightly posterolaterally. A small process extends laterally from the lateral condyle, on which the quadratojugal would have articulated. Medially, the posterior end of the pterygoid is preserved slightly separated from the medial edge of the quadrate condyle.
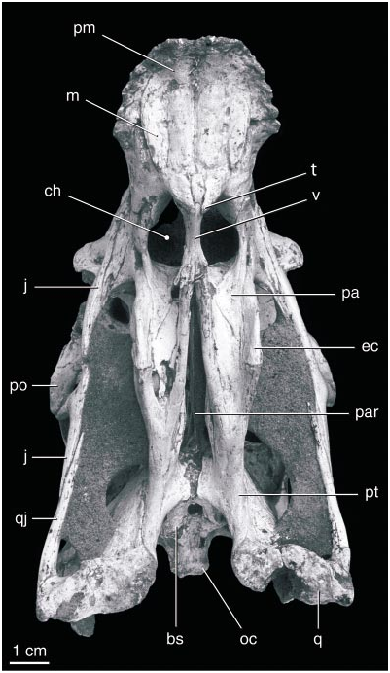
The body of the pterygoid is longitudinally oriented and convex dorsally. The two bones diverge gradually posteriorly, although the interpterygoid vacuity is not exposed. There is no evidence for an epipterygoid, as in Citipati , although this region has not been fully prepared. The ventral part of the basisphenoid is not preserved, including the region of the basipterygoid joint.
The suborbital process of the jugal is incomplete on both sides, and its contact with the quadratojugal is not preserved, so the length of both these bones is indeterminate. The anterior end of the jugal is apparent on both sides, and does not extend anteriorly beneath the antorbital fenestra as it does in Citipati , and is thus similar to Khaan . An intact quadrate foramen is preserved on the right side. It lies mainly in the quadrate, and the quadratojugal is a simple, vertical strap of bone. The foramen is moderately large and about twice as tall as it is wide, similar in size to that of IGM 100/978.
The postorbital is well preserved and similar to that of other oviraptorids. Anterodorsally, it curves medially around the anterior edge of the supratemporal fenestra. A portion of the right frontal adheres to the right postorbital, leading Smith (1993) to mistakenly suggest that the postorbital extended anteriorly over the orbit.
Most of the basioccipital is preserved in natural position, and the foramen magnum is intact dorsal to it. The foramen is narrower than in other oviraptorids, probably due to mediolateral compression postmortem. The bones figured and described by Smith (1993) as the exoccipital and supraoccipital correspond to this region of the skull if they are rotated 90 ° out of position. Thus, one of the ‘‘knoblike paroccipital processes’’ ( Smith, 1993: 374) corresponds to the occipital condyle. Most of the occiput dorsal to the foramen magnum is missing, but the position of the parietal suggests that this part of the occiput was vertical, as in Khaan , rather than facing posterodorsally as in C. osmolskae .
The paroccipital process is well preserved on the right side, but is not preserved on the left. It extends posterolaterally from the foramen magnum and appears to have been deflected slightly posteriorly, perhaps due to mediolateral compression. It is slightly pendant, but its distal end is missing so the extent of its descent cannot be ascertained.
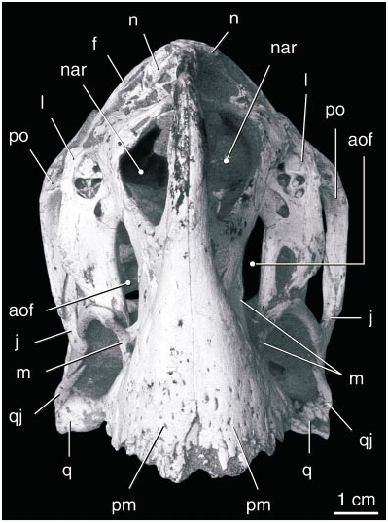
The shape of the narial opening is difficult to determine, because much of this region is not preserved. However, as pointed out by Smith (1993), the ventral edge of the narial opening is preserved where it is bordered by the premaxilla, and it is positioned above the anterior part of the antorbital fossa far from the anterior end of the skull. However, it is unclear that the maxilla bordered the naris because the posterior part of the nasal, which separates the maxilla from the naris in better preserved material, is absent. A slender splint of bone described and figured by Smith (1993), but since lost, probably represents the only part of the nasals preserved, but it does not appear to have been very informative. The premaxillae preserve an ascending process at the anterior end of the skull, but it is unclear how far it rose dorsally. The circumnarial region is quite variable between species of oviraptorids, so it is not possible to reconstruct most of this region in the holotype of O. philoceratops . The presence of a bony ‘‘tooth’’ formed by the maxilla and vomer, as in other oviraptorids, cannot be determined as this area has not been exposed.
The dorsal surface of both frontals is missing along the midline, and the missing portion may have been extensive. The fused parietals bear a tall midline crest, and the dorsomedial inclination of the dorsal surface of each frontal suggests that the crest may have continued anteriorly to above the orbits. This inclination is not due to mediolateral compression, as the ventral surface of the frontals is horizontal. Only the posterior part of the frontals is preserved along the midline, from the ventral part of the bone in the anterior part of the braincase. These fragments suggest that the posterior part of the frontals was filled by a large, presumably pneumatic cavity as in Citipati .
The crest on the parietal lies only along the midline, unlike the domed crest of O. mongoliensis ( Barsbold, 1986) , which occludes the supratemporal fenestrae. The height of the crest may have been accentuated by mediolateral crushing ventrally, but dorsally it appears to be solid and bears no indication of crushing. In lateral view, it forms a gentle arc interrupted by a missing portion anterodorsally. The anterior edge of the crest is preserved descending to the skull roof, but this area may have underlain the frontal and any crest that it bore.
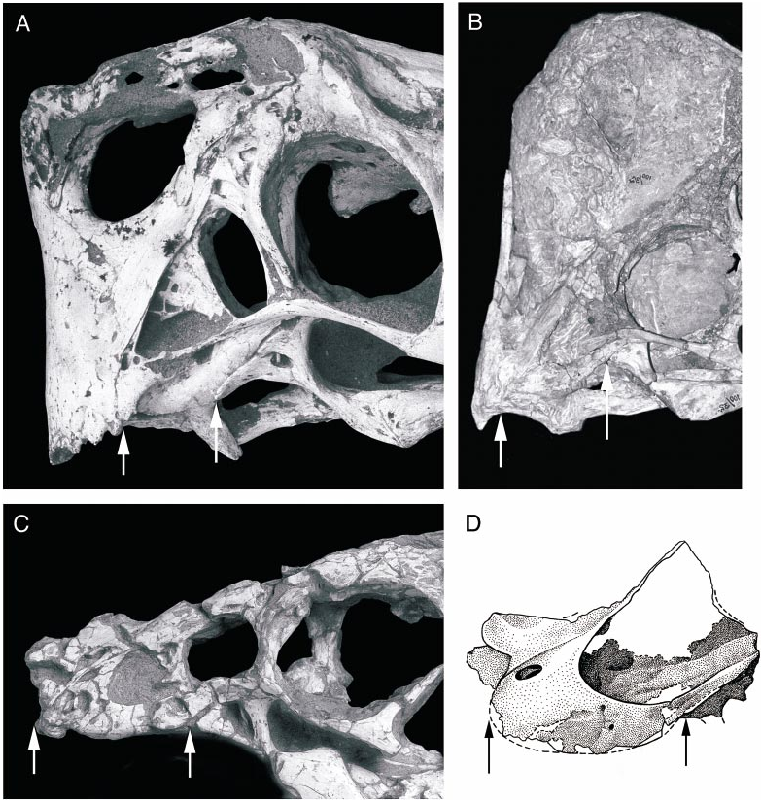
The oval antorbital fenestra is unlike that of other oviraptorids in being elongate longitudinally rather than vertically. A large accessory antorbital fenestra is present in the anterior part of the antorbital fossa (contra Smith, 1993). The entire border is preserved on the left side but is broken on the right. It is similar in size to that on the right side of IGM 100/978. A small opening anterior to the accessory opening on the left side near the premaxillary suture is not duplicated on the right side, but a broader excavation on the left side is in the same position as a small fossa on the right side; most likely this area has been eroded on both sides.
Below the antorbital fossa the suborbital fenestra is exposed between the vertical ectopterygoid and palatine. The fenestra is larger than in IGM 100/978 and other oviraptorids preserving this region. Much of the posterior part of the left ectopterygoid is visible medial to the mandibular ramus, indicating that this bone is similar to that of other oviraptorids. Thus the pterygoid flange broadens vertically rather than horizontally.
The lateral surface of the braincase is well preserved on the left side but has not been fully prepared. A shallow fossa is evident on the prootic dorsal to the fenestra ovale, in a position corresponding to the dorsal tympanic recess. The foramen ovale is exposed anteriorly on the braincase wall and is longer than high. The right laterosphenoid is preserved with the rest of the braincase, whereas the left adheres to the bottom of the left frontal.
The individual bones of the mandible are difficult to differentiate. The surangular is extensive, but almost certainly does not form the ventral edge of the posterior part of the mandibular ramus (cf. figs. 2 View Fig and 3 View Fig of Smith, 1993) or extend anteriorly beneath the mandibular fenestra. The articular probably does not extend anteriorly beyond the quadrate articulation on the lateral surface (the bone labeled by Smith as articular in this region is part of the surangular). The second mandibular foramen, in the posterior part of the ramus, may be an artifact, as this feature corresponds to an area of thin bone in the surangular of IGM 100/978. The spine on the surangular that extends anteriorly into the mandibular fenestra is horizontal rather than extending anteroventrally as in IGM 100/ 978.
The symphysis is poorly preserved and incompletely prepared. Mediolateral compression has dislocated the symphysial portion on the left side from the remainder of the dentary.
Smith (1993: 374) describes ‘‘a sliding articular surface on the quadrate, at its articulation with the articular, permitting an anterior/posterior shearing movement of the mandible.’’ However, in archosaurs, the quadrate condyle is convex and articulates with a concave surface on the articular, so a ‘‘sliding’’ joint can only be implied by an elongate surface on the articular (as it indeed is in the better preserved oviraptorid material; Osmólska, 1976). The articular is preserved well only on the right side, and it has not been prepared sufficiently to discern the length of the articulation surface, but a dorsal concavity evident in medial view suggests that the surface is not as elongate as in Citipati .
The rostrum and lower jaw of O. philoceratops were more elongate than in other oviraptorids, although the extent of this difference is difficult to determine in its poor state of preservation. Compared with other oviraptorids, the premaxilla appears to be incomplete anteriorly, and would have extended one or two centimeters farther than its preserved length. The region anterior to the antorbital fenestra is longer than in other oviraptorids even when the gaps between dislocated pieces of the rostrum are taken into account. This is evident from the greater length of the accessory antorbital fenestra in comparison with other oviraptorids ( fig. 12 View Fig ). If mandibular length is reconstructed by allowing for the large gap between pieces of the right dentary ( fig. 13 View Fig ), then the portion of the dentary anterior to the coronoid process is longer than this region in Citipati , which is typical of other oviraptorids. Furthermore, the descent of the dorsal edge of the dentary anterior to the mandibular fenestra is less pronounced than in other oviraptorids.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |