
Amazophrynella bilinguis, Kaefer & Rojas & Ferrão & Farias & Lima, 2019
|
publication ID |
https://doi.org/10.11646/zootaxa.4577.2.5 |
|
publication LSID |
lsid:zoobank.org:pub:47EBC8B1-604C-4C81-942A-EAAD2BFA2225 |
|
DOI |
https://doi.org/10.5281/zenodo.5944212 |
|
persistent identifier |
https://treatment.plazi.org/id/6E3730AD-CC35-47B1-99F6-E7FEC5382F9F |
|
taxon LSID |
lsid:zoobank.org:act:6E3730AD-CC35-47B1-99F6-E7FEC5382F9F |
|
treatment provided by |
Plazi |
|
scientific name |
Amazophrynella bilinguis |
| status |
sp. nov. |
Amazophrynella bilinguis View in CoL sp. nov. Kaefer, Rojas, Ferrão & Lima
urn:lsid:zoobank.org:act:
Holotype. INPA-H 39784 (field number APL 18244), adult male ( Fig. 1 View FIGURE 1 A–B, 2A–C, 3A, 3C, 4A, 5B) collected on February 2012 by I.L. Kaefer, A.P. Lima and L. Vasconcelos in the farm Fazenda Taperinha ( 2°34’48.4” S; 54°22’16.6” W, 7 m a.s.l.) on the south margin of the Amazon River , Municipality of Santarém, Pará State, Brazil. GoogleMaps
Paratype s. Eleven specimens. Nine males: INPA-H 39782 , INPA-H 39780 , INPA-H 39774 , INPA-H 39776 , INPA-H 39783 , INPA-H 39778 , INPA-H 39781 , INPA-H 39775 , INPA-H 39785 (field numbers APL 18240, APL 18241, APL 18243, APL 18245, APL 18246, APL 18247, APL 18248, APL 18249, and APL 18250, respectively); two females: INPA-H 39779 and INPA-H 39777 (field numbers APL 18242, APL 18251, respectively); all collected by I.L. Kaefer, A.P. Lima and L. Vasconcelos between February 3–5 2012 at the same locality of holotype. Female INPA-H 39777 is designated as the allotype ( Fig. 1 View FIGURE 1 C–D, 2D–F, 3B, 3D, 4B, 5A) .
Diagnosis. A species of Amazophrynella characterized by: (1) SVL 13–14.5 mm in males and 19.6–20.4 mm in females; (2) Finger I Ż Finger II; (3) palmar tubercle rounded; (4) dorsal surfaces highly granular; (5) whitish belly with black tiny spots in life; (6) call type 1 composed by one note with duration of 0.206– 0.292 s and 69–77 pulses; (7) call type 2 with duration of 0.779– 3.191 s and constituted by 6–22 multipulsed notes (9–14 pulses/note).
Comparison with other species. We compared the new species with all currently recognized species of Amazophrynella : A. amazonicola ; A. bokermanni ; A. javierbustamantei Rojas, Chaparro, Carvalho, Ávila, Farias, Hrbek, and Gordo, 2016 ; A. manaos Rojas, Carvalho, Ávila, Farias, and Hrbek, 2014 ; A. matses Rojas, Carvalho, Ávila, Farias, Gordo, and Hrbek, 2015 ; A. minuta ; A. moisesii Rojas, Fouquet, Ron, Hernández-Ruz , Melo- Sampaio, Chaparro, Vogt, Carvalho, Pinheiro, Ávila, Farias, Gordo, and Hrbek, 2018; A. siona ; A. teko Rojas, Fouquet, Ron, Hernández-Ruz, Melo-Sampaio, Chaparro, Vogt, Carvalho, Pinheiro, Ávila, Farias, Gordo, and Hrbek, 2018 ; A. vote Ávila, Carvalho, Gordo, Kawashita-Ribeiro, and Morais, 2012 ; A. xinguensis Rojas, Fouquet, Ron, Hernández-Ruz, Melo-Sampaio, Chaparro, Vogt, Carvalho, Pinheiro, Ávila, Farias, Gordo, and Hrbek, 2018 . Amazophrynella bilinguis sp. nov. can be distinguished from all other Amazophrynella species by the combination of morphological and bioacoustic characters. In addition, the new species differs genetically from all other species in the genus. Characteristics of compared species are presented in parentheses.
The whitish belly in life distinguishes Amazophrynella bilinguis sp. nov. from A. minuta (yellow to orange: Rojas et al. 2018c), A. amazonicola (yellow to reddish orange: Rojas et al. 2015), A. matses (yellow to yellowish orange: Rojas et al. 2015), A. siona (yellow to reddish brown: Rojas et al. 2018a), A. javierbustamantei (pale yellow: Rojas et al. 2016), A. moisesi (pale yellow: Rojas et al. 2018a), A. teko (pale yellow to cream: Rojas et al. 2018a). The Finger I Ż Finger II in A. bilinguis sp. nov. differs from those of all above cited species (Finger I smaller than Finger II: Ávila et al. 2012, Rojas et al. 2014, 2015, 2016, 2018a, 2018c). Additionally, by emitting call type 1 with duration of 0.206– 0.292 s and 69–77 pulses, the new species can be distinguished from A. minuta (0.132– 0.143 s and 43–48 pulses: present study), and A. teko (0.150– 0.190 s and 10–30 pulses: Rojas et al. 2018a). In addition, A. bilinguis sp. nov. emits call type 2 with multipulsed notes that differs from those of A. amazonicola , A. minuta , and A. siona (non-pulsed: Rojas et al. 2018a, b, present study; see Discussion section).
The whitish belly of the new species is similar to A. manaos , A. vote , A. xinguensis and A. bokermanni . The new species differs from A. manaos by having Finger I Ż Finger II (Finger I shorter than Finger II: Rojas et al. 2014), whitish belly with black tiny spots (whitish belly with black blotches: Rojas et al. 2014), and advertisement call type 1 with duration of 0.206– 0.292 s (0.133– 0.156 s: Rojas et al. 2018b). Amazophrynella bilinguis sp. nov. differs from A. vote by reaching 14.5 mm SVL in males and 20.4 mm in females (maximum SVL 19.3 mm in males, and 25.7 mm in females: Ávila et al. 2012) and Finger I Ż Finger II (Finger I <Finger II: Ávila et al. 2012). Moreover, A. bilinguis sp. nov. differs of A. vote by having call type 1 with duration of 0.206– 0.292 s and 69–77 pulses (0.098– 0.150 s and 41–60 pulses: Rojas et al. 2018b).
According to the genetic analyses (see section Phylogenetic relationships), A. xinguensis and A. bokermanni are the most closely-related nominal species in relation to Amazophrynella bilinguis sp. nov. The new species can be distinguished from A. xinguensis by having SVL 13–14.5 mm in males and 19.6–20.4 mm in females (SVL males 17.7–20.0 mm; SVL females 22.4–26.3 mm: Rojas et al. 2018a), palmar tubercle rounded (elliptical: Rojas et al. 2018a). The advertisement call of A. xinguensis is unknown. Amazophrynella bilinguis sp. nov. is differentiated from A. bokermanni by SVL 13–14.5 mm in males (SVL = 15.9–16.5 mm: Rojas et al. 2018a), dorsal surfaces highly granular (granular), black tiny spots on venter (small black dots). Moreover, the advertisement call type 1 of A. bilinguis sp. nov. can be distinguished from those of A. bokermanni by duration of notes 0.206– 0.292 s (0.125– 0.163 s: Rojas et al. 2018b).
Description of holotype. Body small, elongate. Head triangular in dorsal and lateral view. Head longer than wide. HL 38.04% and HW 30.43% of SVL. Snout acuminate in lateral view and triangular in dorsal view. SL 44.4% of HL. Nostrils protuberant, closer to snout than eyes. Cantus rostralis straight in dorsal view. Internarinal distance smaller than eye diameter. IND 28.6% of HW. Upper eyelid covered by small granules. Eye prominent, 29.0% of HL. Tympanum not visible through the skin. Texture of skin on tympanum covered with granules. Texture of dorsal skin highly granular. Texture of dorsolateral skin granular. Abundance of granules on arms insertion. Forelimbs thick. Upper arms robust. UAL 39.1% of SVL. Presence of granules on upper arm and arm. HAL 20.9% of UAL. Fingers slender. Tips unexpanded. Fingers basally webbed. Relative length of Fingers: I Ż II <IV <III. Thumb larger and robust. Supernumerary tubercle rounded: two on Finger I and II and IV and three in Finger III. Palmar tubercle rounded, approximately ¼ of hand. Gular region granular. Texture of ventral skin granular. Cloacal opening slightly above midlevel of thighs. Hind limbs slender. Thigh to tarsus covered by spiny protuberances. THL 58.2% of SVL. TAL 50.6% of SVL. Presence of small granules on tibia. Tarsus 36.8% of SVL. FL 60.5% of THL. Relative length of toes: I <II <III <V <IV. Inner metatarsal tubercle rounded. Outer metatarsal tubercle small and oval. Subarticular tubercles rounded: one on toes I, II and V, three on toes III and IV; foot with slender, basally webbed toes.
Measurement of the holotype ( in mm). SVL: 13.8; HW: 4.2; HL: 5.2; SL: 2.3; ED: 1.5; IND: 1.2; UAL: 5.4; HAL: 2.9; FI: 1.5; FII:1.5; THL: 8.1; TAL: 7.5; TL: 5.1; FL: 4.9.
Coloration of the holotype. In life, head brown in dorsal view. Dorsum brown with brown chevrons. Flanks light brown. Dorsal surfaces of upper arm, arm and hand light brown. Dorsal surfaces of thighs, tibia, tarsus and foot light brown. Gular region cream with brown spots. Belly whitish with brown spots and white granules ( Fig. 5B View FIGURE 5 ). Ventral surfaces of upper arm and arm creamy. Palm reddish. Ventral surfaces of thighs, tarsus and tibia creamy, sole black. Iris golden and pupil black.
In preservative, the coloration is almost the same than the one in life. The coloration of the dorsum becomes light brown. Gular region and venter become cream. The iris loses its coloration. Fingers and toes become cream ( Fig. 6 View FIGURE 6 ).
Variation. There is little variation among specimens of the type series ( Table 2). Sexual dimorphism was detected in SVL, with 13.0– 14.5 mm (13.65 ± 0.43 mm, n = 10) in males and 19.56–20.4 mm (19.98 ± 0.59 mm, n =2) in females. Three specimens (INPA-H 39775, INPA-H 39780, INPA-H 39776) have few granules on posterior region of head. The specimen INPA-H 39779 present a white line from head to cloaca. There is variation in the size (mm) of the FI and FII between individuals, while some (e.g. INPA-H 39778, INPA-H 39774, INPA-H 39783) present Finger I> Finger II and other present Finger I Ż Finger II (e.g. INPA-H 39782, INPA-H 39784, INPA-H 39775). Subarticular tubercles more protruding and swollen in females. The specimen INPA-H39777 shows small spots on dorsolateral surfaces. Dorsum with different tonalities of brown (light brown to brown). Spots on venter vary in sizes (small to medium size). Thighs, shanks and tarsus between cream and whitish coloration, in ventral view. Palm and sole present different tonalities of cream, in ventral view. In preserved specimens, the palmar tubercle is more flattened.
Advertisement call. Two different call types were recorded ( Fig. 7 View FIGURE 7 A–D), both of them from the same two individuals. Additional males, not recorded, also emitted both call types. We did not categorize these vocalizations as a single composed advertisement call because each one of the call types can be repeated for several minutes. The calls of type 1 ( Fig. 7A View FIGURE 7 ) consist of one multipulsed note ( Fig. 7C View FIGURE 7 ) with average duration of 0.248 ± 0.020 s (0.206– 0.292 s) and are composed by 72 ± 2 pulses (69–77 pulses). Calls type 1 are emitted in series ( Fig. 7A View FIGURE 7 ) and are interleaved by silent intervals of 1.076 ± 0.162 s (0.621– 3.779 s). Calls type 1 have a mean dominant frequency of 3526 ± 170 Hz (3338–4264 Hz), low frequency of 3058 ± 119 Hz (2674–3234 Hz) and high frequency of 4478 ± 173 Hz (4277–4744 Hz).
Different from calls of type 1, the calls of type 2 are arranged in bouts ( Fig. 7B View FIGURE 7 ) of notes and have a duration of 1.9 ± 0.77 s (0.779– 3.191 s). The mean number of notes per call is 13 ± 5 (6–22 notes) and notes are 0.034 ± 0.001 s (0.010– 0.065 s) long in average. The duration of the silent interval between notes in call type 2 is 0.125 ± 0.028 s (0.085– 0.206 s). Notes are composed of 12 ± 1 pulses (9–14 pulses) ( Fig. 7D View FIGURE 7 ). The mean dominant frequency of calls type 2 is 3450 ± 68 Hz (3220–4680 Hz), while the mean low frequency is 3200 ± 18 Hz (2961–3410 Hz) and high frequency is 4002 ± 91 Hz (3345–4680 Hz).
Phylogenetic relationships. Our phylogenetic tree inferred from 559 aligned sites of 16S rRNA ( Fig. 8 View FIGURE 8 ) was partially concordant with Rojas et al. (2018a). All nominal species of Amazophrynella in our reconstruction formed well supported clades (PP Ż 0.95), excepted A. siona , and A. teko . Amazophrynella bilinguis sp. nov. was recovered as the sister taxon of Amazophrynella sp. 3 ( sensu Rojas et al. 2018a) from Parque de Desenvolvimento Jatobá (Tapajós River, Pará, Brazil) with low support (PP = 0.74).
Genetic distance between A. bilinguis sp. nov. and Amazophrynella sp. 3 was relatively low (p and K2P = 2%) compared to distances calculated between A. bilinguis sp. nov. and other Amazophrynella species that ranged from 3% (p) and 4% (K2P) ( Amazophrynella sp. 2) to 14% (p) and 16% (K2P) ( A. siona ). Amazophrynella bilinguis sp. nov. presented 4% (p and K2P) genetic distance from samples of A. bokermanni , the nominal species that was placed closest to it in the phylogenetic tree. See Table 3 for genetic distances among all analyzed species.
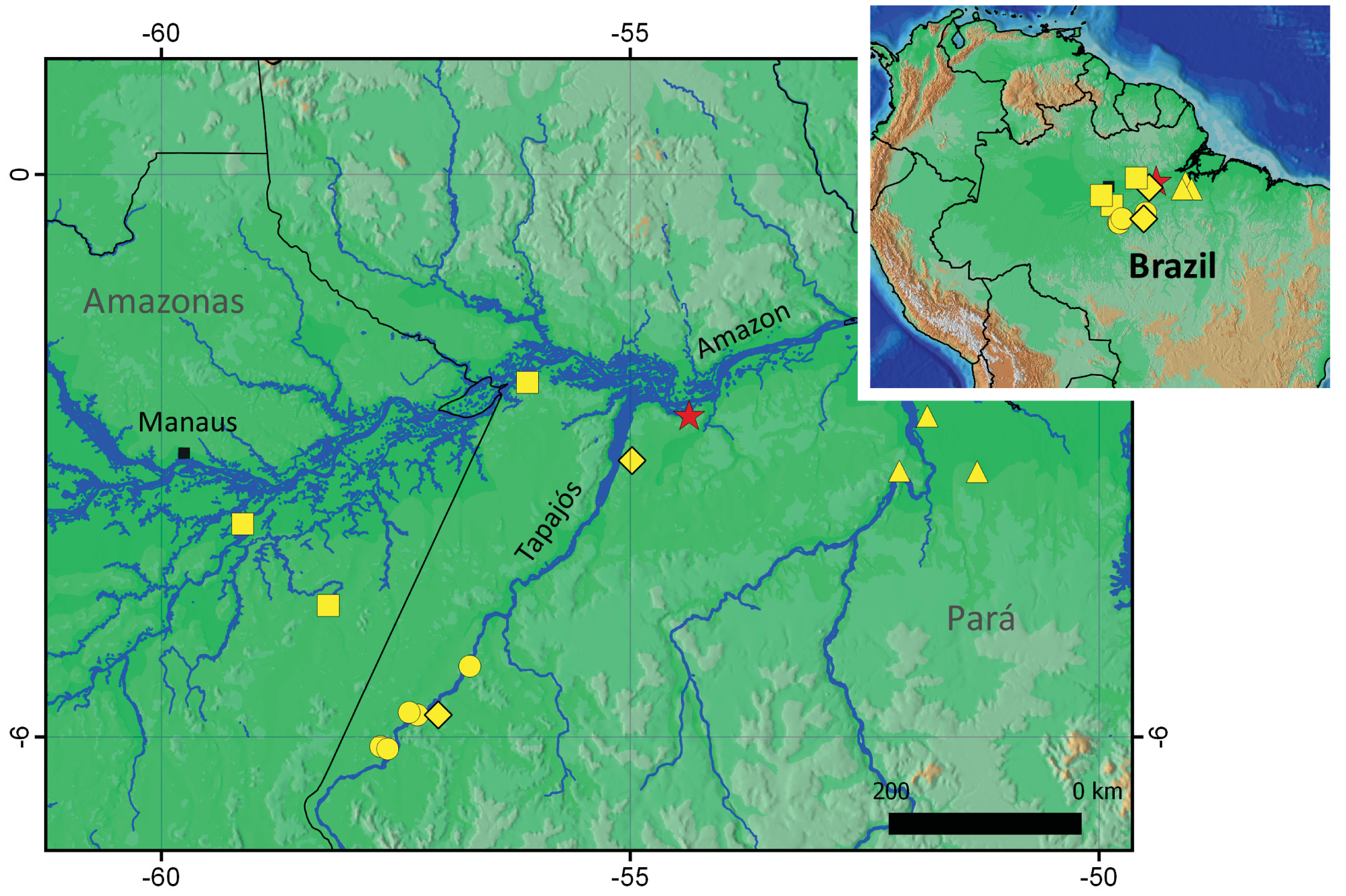
Distribution and natural history. Amazophrynella bilinguis sp. nov. is known only from Fazenda Taperinha ( 2°34’48.4” S; 54°22’16.6” W, 7 m a.s.l.) on south margin of the Amazon River, Municipality of Santarém, Pará State, Brazil ( Fig. 9 View FIGURE 9 ). However, it is likely that sampling efforts, which are historically scarce at that region, might expand the known distribution of this taxon.
Like in other species within the genus, males of this species were found calling during the day, usually perched up to 10–40 cm above the ground on vegetation, fallen logs, branches or on leaf litter on the forest floor. Females and vocally active males were found near a stream and associated shallow ponds, where egg deposition and tadpole development may occur. After collection, a pair (female = INPA-H 39779, SVL = 20.8 mm; male = INPA-H 39774; SVL = 15.4 mm) engaged in axillary amplexus inside a plastic bag in the improvised field laboratory ( Fig. 4A and 4B View FIGURE 4 ).
Etymology. The epithet " bilinguis " is Latin and means "bilingual". It refers to the two distinct advertisement calls emitted by the male individuals.






No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.