
Himantolophus litoceras, Stewart, Andrew L. & Pietsch, Theodore W., 2010
|
publication ID |
https://doi.org/ 10.5281/zenodo.199134 |
|
DOI |
https://doi.org/10.5281/zenodo.6197496 |
|
persistent identifier |
https://treatment.plazi.org/id/60038798-FFAC-0755-FF30-FF2FC3D7FCA7 |
|
treatment provided by |
Plazi |
|
scientific name |
Himantolophus litoceras |
| status |
sp. nov. |
Himantolophus litoceras View in CoL new species
Figures 1 View FIGURE 1 , 2 View FIGURE 2 , 3 View FIGURE 3
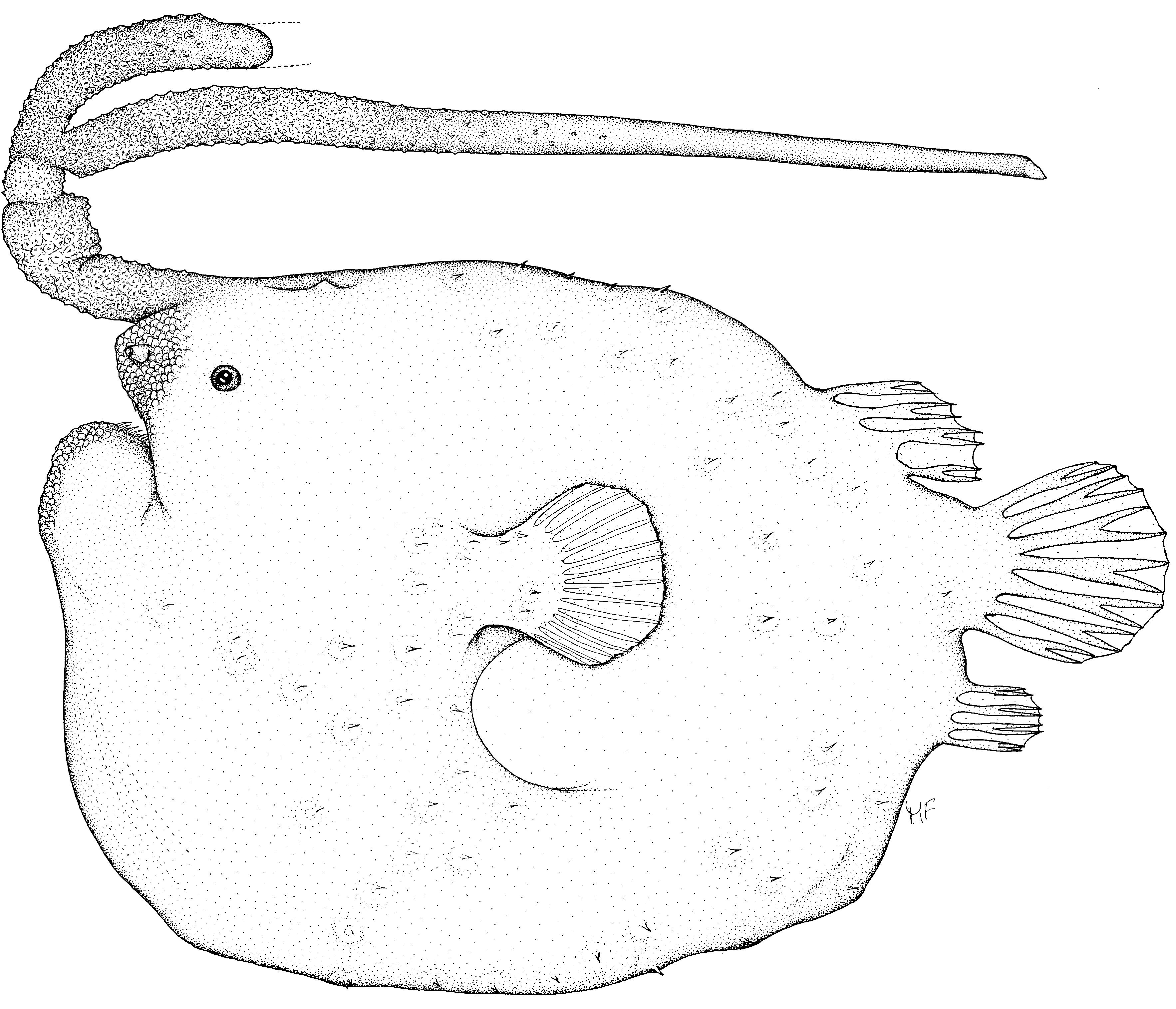
Holotype. NMNZ P. 042004, female, 276 mm SL (263 mm SL preserved), F/V Seamount Explorer, Station OBS 2170/130, South Cavalli Seamount, east of North Cape, North Island, New Zealand, 34º12.0'–14.0'S, 175º05.0'–07.0'E, bottom trawl, 654 m, collected by Ted Turton, MFish Scientific Observer, 7 November 2005.
Diagnosis. Metamorphosed females of Himantolophus litoceras differ from those of other members of the H. cornifer -group in having a pair of simple distal escal appendages, each lacking any trace of lateral appendages or filaments. It further differs in having the following combination of character states: length of illicium 31.1% SL (28.2% preserved); width of escal bulb 11.9% SL (10.5% preserved); illicium and esca without filaments; illicium, esca, and proximal one-half of distal escal appendage covered with tiny, close-set dermal spinules; distal escal appendages darkly pigmented, tip of intact filament silvery white in freshly thawed specimen.
Description. Holotype somewhat damaged by trawl, lower jaw broken, partially missing; about threequarters length of left distal escal appendage lost prior to capture, tip healed, showing no recent signs of injury; tip of intact distal escal appendage with some signs of damage, may not be complete. Distal escal appendages separated nearly from base, escal bulb with two lobes, escal pore on posterior margin of bulb, opening on a small raised protuberance. Stem of illicium darkly pigmented, escal bulb at base of distal escal appendages white; body without unpigmented patches of skin. Dorsal-fin rays 5, two anteriormost rays simple, but closely aligned (appearing as one), remaining rays bifurcated; anal-fin rays 4, bifurcated; pectoralfin rays 16, all simple and flattened; caudal-fin rays 9, upper- and lowermost ray closely aligned with adjacent ray for most of length (each appearing as one), innermost five rays bifurcated. Four dermal spines on lateral surface of each pectoral fin lobe; approximately 55 dermal spines of each side of body. Papillae of snout and chin well developed. Number of teeth in upper jaw 29 + 28, longest tooth 4.1 mm; longest tooth in lower jaw 7.8 mm. Eye diameter 9.8 mm (6.4 mm preserved); stem of illicium 86 mm (74.3 mm preserved); width of escal bulb 33 mm (27.8 mm preserved); length of intact distal escal appendage 327 mm (319 mm preserved). Additional characters as provided for H. cornifer -group by Bertelsen and Krefft (1988) and Pietsch (2009).
Discussion. Himantolophus litoceras n. sp. is a member of the H. cornifer -group as defined by Bertelsen and Krefft (1988), lacking posterior escal appendages. Within the H. cornifer -group, the new species differs from the other members in lacking lateral branching along the distal escal appendage. It is most similar to H. macroceratioides Bertelsen and Krefft, 1988 (known from two specimens) in the length of the distal escal appendage and width of the escal bulb as a percentage of SL, as well as in escal pigmentation. However, the base of the illicium and most of the distal escal appendage are naked or only sparsely covered with dermal spinules in H. macroceratioides , whereas these regions are densely covered with spinules in H. litoceras .
Himantolophus litoceras View in CoL n. sp. is similar to H. macroceras Bertelsen and Krefft, 1988 View in CoL (known from five specimens) in the diameter of the escal bulb, and length of the illicium and distal escal appendage as percentage of SL. It also shares with H. macroceras View in CoL the dense covering of dermal spinules along most of the illicial stem, escal bulb, and proximal half of the distal escal appendage. However, the distal half of the distal escal appendages of H. litoceras View in CoL is naked, whereas the spinules continue to the tips of the appendage in H. macroceras View in CoL .
Himantolophus azurlucens Beebe and Crane, 1947 View in CoL is represented by two specimens, each bearing a side branch on only one branch of the distal escal appendage, but the length of these appendages is considerably shorter (23% SL right, 32% SL left).
Himantolophus cornifer Bertelsen and Krefft, 1988 View in CoL (eight specimens) has one to three distal bifurcations of the distal escal appendage, whereas that of H. litoceras View in CoL n. sp. is bifurcated only at its base. Himantolophus compressus ( Osório, 1912) View in CoL was known from a single specimen that was subsequently destroyed by a fire at the Museu Bocage, Lisbon, in 1978. The illustration by Osório, reproduced by Maul (1961:101, figure 4), is somewhat crude, showing no details of the esca. Maul was unable to examine the type, but relied on Osório’s account and communication with an assistant at the Museu Municipal do Funchal. He noted that there was damage to the esca, most notably to the distal escal appendage ( Maul, 1961:103). Prior to its loss, the holotype was re-examined by Bertelsen who reported that only about 5 mm of the distal escal appendage remained ( Bertelsen and Krefft, 1988:73). Bertelsen noted, however, that the distal escal appendage arose from a single point on the bulb and bifurcated some distance up, differing from that of H. litoceras View in CoL , which bifurcates at its base. The distal escal appendage percentage length and presence/absence of lateral appendages was not reported by Osório (1912).
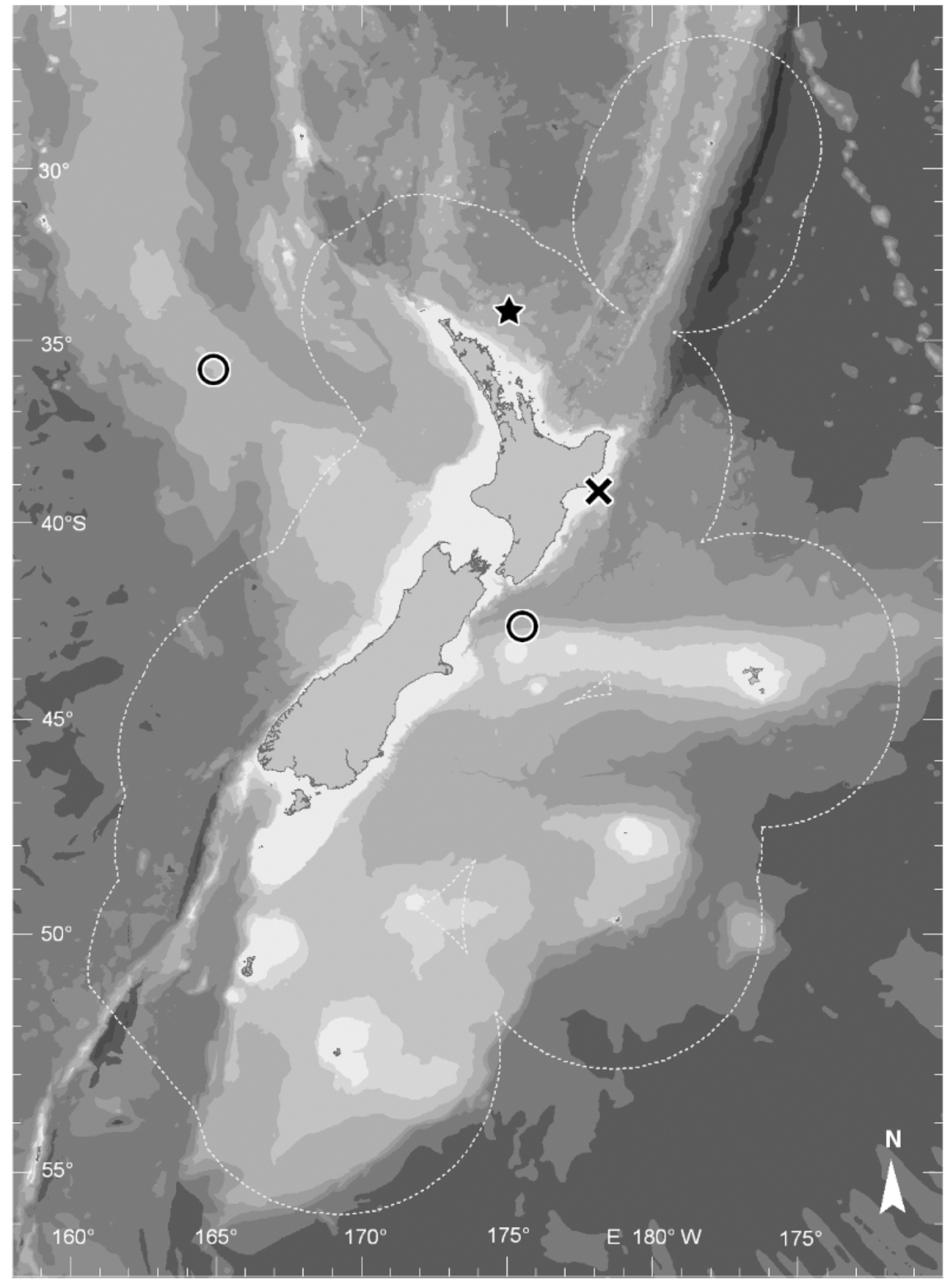
Distribution. Himantolophus litoceras n. sp. is known from a single specimen taken off the northeast coast of the North Island, New Zealand, in 654 m, which is relatively shallow water ( Fig. 3 View FIGURE 3 ). Members of the H. cornifer -group have been caught over a wide depth range, from 170 to 2150 m depth, tending to form a bimodal vertical distribution between 170 m and 900 m and 1650 m and 2150 m. However, with so few specimens of the H. cornifer -group in collections, it is impossible at this time to determine if there is species specific depth stratification.
The five other species in the cornifer -group are all recorded from equatorial to subtropical waters ( Pietsch, 2009:210, figs. 213, 214). The exception to date has been Himantolophus compressus , which was caught off Portugal at 38° 20' N. A recent record of H. cornifer from 29° 25' S was recovered from the stomach of a beached sperm whale ( Anderson and Leslie, 2001:8). Himantolophus litoceras n. sp is the first record of the H. cornifer -group from the South Pacific and is the southernmost record for this group.
Metamorphosed females of ceratioids of most species do not appear to undertake sustained swimming. Their body shape is generally globose with weak musculature and small posteriorly set dorsal and anal fins. In situ observations of a species of Oneirodes confirmed that they are, for the most part, drifters ( Luck and Pietsch, 2008). Species of the Himantolophidae have the same body pattern as Oneirodes , so the drifting mode can be expected to apply to them as well.
Himantolophus litoceras n. sp was taken in the subtropical water mass, subject to the East Auckland Current, which diverges from the eastward-flowing Tasman Front. The northeast coast of the North Island is also a region that experiences a number of eddies spinning off from this current (e.g., North Cape Eddy and East Cape Eddy) before it breaks up to the south-bound East Cape Current and less-defined currents moving north and east ( Carter et al., 1998). Based on this water movement and the distribution of other himantolophid anglerfishes across the New Zealand and south-eastern Australian region, this species can be expected to also occur in the Tasman Sea as well as the southwest Pacific Ocean.
Etymology. From a combination of the Greek litos meaning “simple” or “plain,” and keras meaning “horn,” in reference to the lack of lateral branching or filaments on the distal escal appendages.
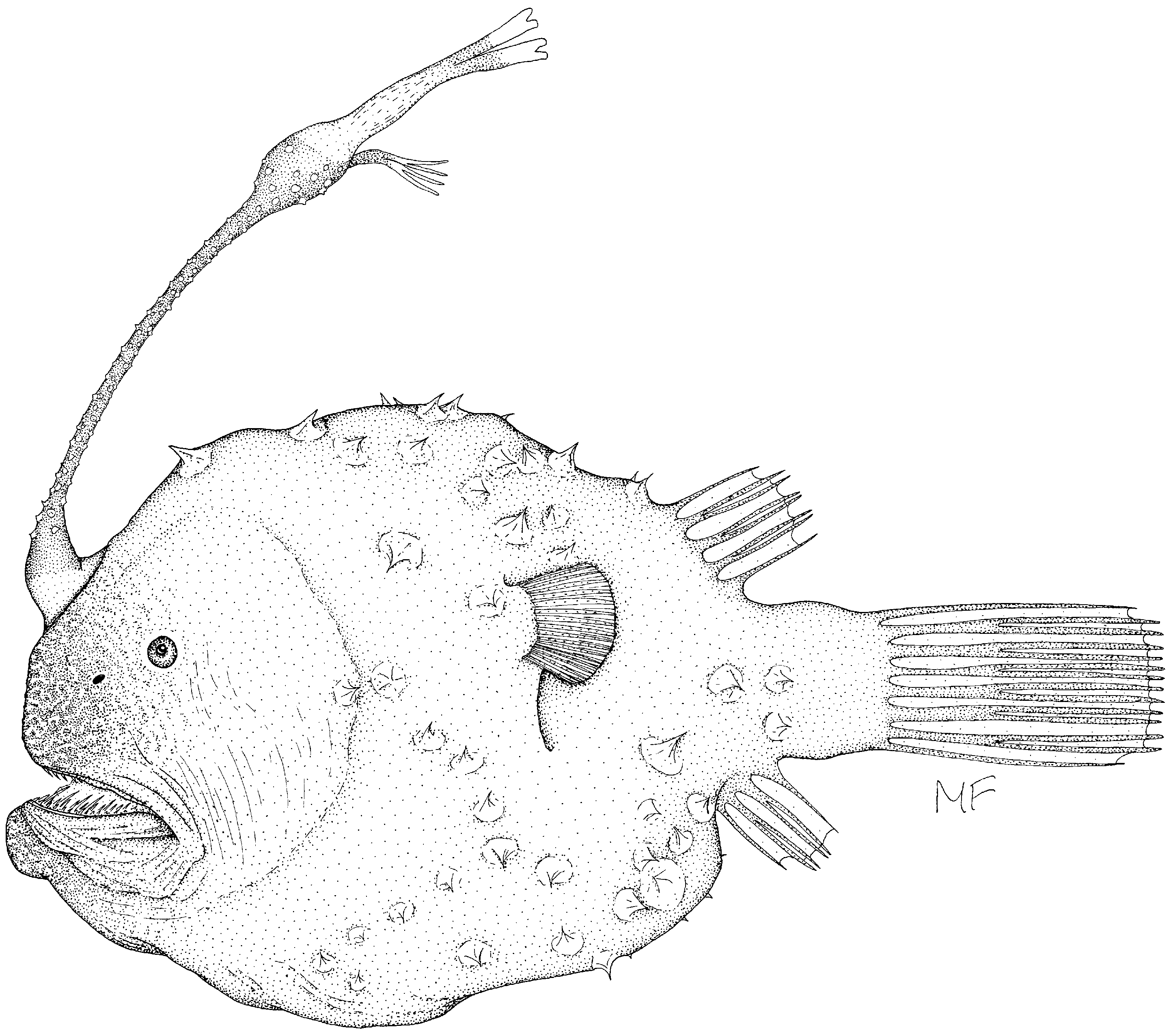
Comments on the status of Himantolophus pseudalbinares Bertelsen and Krefft, 1988 . In his paper recording Himantolophus albinares from the Lord Howe Basin, Iglésias (2005) described a single large (207 mm SL) specimen. On the basis of what he believed to be a divided tip on one of the pair of distal escal appendages, he proposed that Himantolophus pseudalbinares is a synonym of H. albinares . Re-examination of Iglésias’s specimen, two specimens of Himantolophus albinares held by the Museum of New Zealand Te Papa Tongarewa (Fig 4,5), and the second known specimen of H. pseudalbinares reported by Stewart and Pietsch (1998) ( Fig. 6 View FIGURE 6 ), revealed a number of features that challenge Iglésias’s conclusion.
Both the illustration of the holotype of H. pseudalbinares ( Bertelsen and Krefft, 1988: Fig. 23) and the second specimen reported by Stewart and Pietsch (1998) clearly show that the tips of both branches of the distal appendages are precisely and symmetrically divided ( Fig. 6 View FIGURE 6 ). The apparent division of the specimen figured by Iglésias is long, distinctly ragged, and well down from the tip. Comparison with a specimen of similar size and condition in the Museum of New Zealand collection (NMNZ P. 031256; 35° 50.9'S, 164° 57.0'E) shows similar ragged tips to the DA ( Fig. 4 View FIGURE 4 ). In the case of the NMNZ specimen, this is due to the distal escal appendages becoming thin and flaccid, with indications that the esca is beginning to break down. Dissection of that specimen also revealed that the ovaries were large with extremely thin transparent walls and no sign of any eggs. Indications are that this specimen, and the specimen described by Iglésias, were probably undergoing senescence.
The diagnosis of Himantolophus albinares provided by Bertelsen and Krefft (1988) and the description by Iglésias (2005) record the absence of dermal spinules on the upper escal bulb and appendages. However, the second specimen of Himantolophus pseudalbinares has small but distinct spinules along most fo the length of the distal escal appendages, extending almost onto the tips. The senior author sent late E. Bertelsen this second specimen for his evaluation and had the identification confirmed. Small differences between that specimen and the holotype (relatively longer illicium, absence of distinctive illicial filaments and spines on the illicium) were, he believed, within the range of variation observed in its nearest relative (E. Bertelsen, pers. comm. 1992).
On the basis of our examinations and analysis, we agree with Iglésias that his specimen (MNHM 2003- 00118) is Himantolophus albinares , but that the supposed division of the distal appendage on that specimen is an artefact of illicial tissue break-down rather than evidence supporting synonymy. Therefore, in full agreement with Pietsch (2009:344), we propose that Himantolophus pseudalbinares should be retained as a valid species supported by the diagnostic characters defined by Bertelsen and Krefft (1988).
Conclusion. The capture of a new species in relatively shallow water, and the extended distribution of what has up until recently been considered an Atlantic species group, raises an number of issues about ceratioid distributions within the New Zealand EEZ.
Ceratioids tend to be small (<300 mm SL), soft-bodied and occur in low densities; one per 800,000 m 3 ( Helfman et al., 1997:298). Most captures in the New Zealand EEZ have been as incidental trawl by-catch whilst targeting deep-water commercial species (e.g. orange roughy Hoplostethus atlanticus and oreo dories Oreosomatidae ). The use of large mesh with a 4 cm cod-end, the need to maximise commercial returns or the constraints of stratified research tows have tended to result in few ceratioid specimens being caught intact and fewer being retained. This is compounded by the mid-water depths below 1200 m being rarely sampled within the New Zealand region.
Research trawling within the New Zealand EEZ by the former Ministry of Agriculture and Fisheries and NIWA (subsequent to 1995) over the 38-year period 1961–1997 covered 5326 km 2, most of which was shallower than 1200 m depth, and sampling by other institutions (universities, museums, etc.) amounted to an additional 2 km 2 ( Nelson and Gordon, 1997). This is in an EEZ of over 4 million km2, of which 72% lies below 1000 m depth and 51% is below 2000 m.
Metamorphosed female ceratioids tend to be most abundant below 1000 m depth ( Paxton, 1990; Nelson, 2006), therefore, the faunal list of this group for the New Zealand region (e.g. Roberts et al., 2009) is probably far from complete. Gaps in collecting reflect gaps in the knowledge of the distribution of the group as a whole. Fourteen percent of the 160 ceratioid species recognised to date are known from a single specimen; 48% from six or fewer ( Pietsch, 2009). Whilst some families such as the Ceratiidae have been relatively stable in their composition, others have continued to have new species added. The majority of these have been in the Oneirodidae (e.g., Pietsch, 2004, 2007), with some in the Thaumatichthyidae and others ( Pietsch, 2005). In addition, new catches of rare species have warranted expanded re-descriptions (e.g., Kharin and Pietsch, 2007) or re-evaluations (e.g., Pietsch and Kharin, 2004).
Given the low sampling intensity and large volume of habitat available, together with small body size, discovery of new ceratioid species and expanded distributions of known taxa may be expected to continue in the New Zealand EEZ.


| NMNZ |
Museum of New Zealand Te Papa Tongarewa |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Himantolophus litoceras
| Stewart, Andrew L. & Pietsch, Theodore W. 2010 |
H. macroceras
| Bertelsen and Krefft 1988 |
Himantolophus cornifer
| Bertelsen and Krefft 1988 |
Himantolophus azurlucens
| Beebe and Crane 1947 |
Himantolophus compressus ( Osório, 1912 )
| Osorio 1912 |